七带石斑鱼早期形态发育与仔稚幼鱼培育的观察研究
2014-12-15周庆杰于道德高天翔官曙光
周庆杰, 于道德, 高天翔, 官曙光
(1.中国海洋大学 水产学院, 山东 青岛 266071; 2.山东省海洋生物研究院, 山东 青岛 266002)
七带石斑鱼(Epinephelus septemfasciatus) 隶属鲈形目(Perciformes), 鮨科(Serranidae), 石斑鱼属(Epinephelus), 是该属中大型的石斑鱼类。该鱼为暖温性礁栖鱼类, 主要分布于日本、韩国、中国的黄海和东海沿岸的浅水区域, 是石斑鱼在黄海唯一分布的鱼种[1], 因其能够耐受 7~8℃的低温冷水, 又称为“冷水石斑”[2]。七带石斑鱼具有生长快、耐低温、适应能力强、营养物质丰富、肉质鲜美等优点, 是石斑鱼属中唯一可以在较低温下生活、适合在北方养殖的, 如今已成为中、日、韩三国海水鱼类繁育研究的热点, 具有较高的经济价值和市场潜力。
目前, 七带石斑鱼的繁殖生物学[1,3-6]、人工育苗[7-,8]、病害研究[9-11]、肌肉成分分析[12]、染色体核型[13]等研究虽然已有报道, 国内七带石斑鱼的人工繁殖和育苗仍处于试验阶段, 繁育技术尚未成熟, 使七带石斑鱼规模化养殖受到制约。作者详细描述了七带石斑鱼胚胎发育及仔、稚、幼鱼的形态变化特征, 提供了七带石斑鱼生物学资料, 为大规模开展七带石斑鱼工厂化苗种生产提供科学依据。
1 材料与方法
1.1 受精卵孵化
实验于2012年2~8月在烟台百佳水产有限公司进行, 试验亲鱼从韩国购进, 2012年2月选择30尾5~7 龄亲鱼进行培育。采用激素肌肉注射催产, 雌鱼混合催产注射量为绒毛膜促性腺激素(HCG)700IU/kg体质量, 促黄体素释放激素(LRH-A3)6µg /kg体质量。雄鱼注射量为HCG200单位/kg。注射催产激素后 72 h, 将雌鱼用特制担架小心捞起, 盖住鱼眼, 请按鱼腹, 让卵子自然流入烧杯。雄鱼操作大致同雌鱼, 用 1.5 mL吸管吸取精液后, 加入适量的海水激活, 然后加入到盛有鱼卵的烧杯中授精, 搅拌使精卵均匀, 充分受精。将获得的受精卵在水温23.5℃±0.5℃、盐度 31、pH7.9~8.2、溶氧量 6~8mg/L、光照强度1 500 ~2 000 lx、连续微充气, 流水孵化。
1.2 胚胎发育观察
胚胎发育期间, 定时从孵化箱中取样, 在Olympus sz61解剖镜下进行胚胎发育的观察测量,详细记录发育时间及各个发育时期的主要形态特征,并用数码相机对各个发育时期拍照。以 50%的受精卵孵出仔鱼作为胚胎发育结束的标志。
1.3 苗种培育及观察
育苗池为 4 m×4 m×1.5 m 的方形室内水泥池,苗种培育期间的水温变化范围为 24~26 ℃, 盐度28。以长牡蛎(Crassostrea gigas)受精卵及其幼体为仔鱼的开口饵料; 随着仔鱼的不断长大, 所用饵料依次过渡为S型轮虫(Brachionus)、L型轮虫、卤虫(Artemia salina)无节幼体、鱼肉糜、配合饲料。培育期间根据水色和水质的变化情况适时适量添加小球藻(Chlorella)及洁净海水。自布池开始, 仔鱼期连续每天取样, 稚、幼鱼期每隔3~5 d取样, 每次取样20尾, 用Olympus sz61解剖镜和Nikon TS100显微镜进行观察, 详细记录描述其形态特征, 并用数码相机拍照。
2 结果与分析
2.1 胚胎发育
七带石斑鱼受精卵呈圆球形, 无色、透明, 彼此分离, 卵径为(0.854±0.021)mm(n=30), 油球直径为(0.181±0.006)mm(n=30)。受精卵为端黄卵, 胚盘形成于动物极, 在相对静止的状态下, 盐度31时上浮,动物极朝下, 植物极朝上。在水温(23.5±0.5)℃、盐度31的海水中孵化, 历时31 h5 min, 50%以上受精卵孵出仔鱼, 胚胎发育过程完成。与其他石斑鱼属石斑鱼一样, 七带石斑鱼的胚胎发育可分为 5个阶段,即: 卵裂期、囊胚期、原肠胚期、神经胚期和器官形成期。胚胎发育过程及特征见表1。
2.1.1 卵裂期
七带石斑鱼的卵裂方式属于盘状卵裂。受精后15 min, 卵裂开始发生, 卵裂仅发生在动物极所在的半球; 受精后2 h20 min, 经过6次细胞分裂(图1-1~图1-6)进入 64细胞期(图1-7), 细胞排列不规则, 且细胞形状和大小也不尽相同, 分裂面紊乱不清, 细胞层开始出现重叠现象(图1-7); 在受精后2 h50 min,细胞变小变多, 进入多细胞期(图1-8); 在受精后4 h 15 min, 细胞变得更小, 不可计数, 整个细胞团呈圆形, 状如桑葚, 进入桑葚期(图1-9)。
2.1.2 囊胚期
5 h 30 min左右, 随着卵裂继续进行, 细胞数目与细胞层次不断增加, 胚盘与卵黄之间形成囊胚腔,囊胚中部向上隆起, 胚胎进入高囊胚期(图1-10); 此后, 囊胚隆起部分逐渐变低, 6 h 30 min囊胚隆起变得最低, 胚盘逐渐扁平, 细胞开始下包, 进入低囊胚期(图1-11)。

表1 七带石斑鱼胚胎发育过程Tab.1 Embryonic development of E.septemfasciatus
2.1.3 原肠胚期
在囊胚期之后, 根据卵黄被胚层下包的程度,胚胎依次经历原肠早期(图1-12)、原肠中期(图1-13)、原肠晚期(图1-14)。
2.1.4 神经胚期
受精后12 h 45 min胚体背面增厚, 形成神经板,中央隐约可见一条圆柱形脊索, 胚体轮廓清晰, 进入胚体形成期(图1-15)。
2.1.5 器官形成期
受精后14 h 35 min胚体头部两侧出现1对视囊,进入视囊形成期(图1-16); 受精后15 h15 min脊索两侧出现长方形的体节, 胚体进入肌节出现期, 视囊轮廓变得更加清晰(图1-17); 受精后15 h45 min在胚体头部视囊靠后的位置出现1对听泡, 肌节数增多,胚体进入听囊形成期(图1-18); 受精后17 h 35 min胚体背面的视囊之间出现椭圆形板状脑泡, 脑泡尚未分室, 胚体进入脑泡形成期(图1-19); 受精后21 h 25 min胚体背腹两处逐渐形成鳍褶, 尾部有少部分与卵黄囊分离, 尾鳍褶可见, 胚体进入尾芽期(图1-20); 受精后23 h15 min胚体开始间歇抽动, 视囊
内可见折光性强的晶体(图1-21); 受精后26 h15min心脏开始微弱跳动, 胚体进入心跳器(图1-22); 受精后30 h30 min胚体的扭动更频繁有力, 进入出膜前期(图1-23); 受精后30 h45 min进入出膜期, 胚体头部先顶破卵膜, 然后尾部用力摆动, 脱去卵膜。

图1 七带石斑鱼胚胎发育Fig.1 Embryonic development of E.septemfasciatus
2.2 仔稚幼鱼形态发育特征
七带石斑鱼仔稚鱼各期的发育和形态特征如图2所示。
2.2.1 仔鱼期
初孵仔鱼 全长 1.99 mm±0.041 mm, 卵黄囊呈椭球形, 长径 0.92 mm±0.012mm, 短径 0.55 mm±0.014mm。油球 1个, 圆球形, 油球直径 0.18 mm±0.012mm, 大部分仔鱼油球位于卵黄囊后端, 一部分仔鱼的油球位于卵黄囊下端, 极少数仔鱼的油球位于卵黄囊前端。刚孵出的仔鱼, 鱼体透明, 脊索略有弯曲, 肌节 22~24对, 肛前肌节 8~9对, 肛后肌节14~15对。头部上方, 背腹部, 尾部有无色透明的鳍膜连成一体。肛门稍离开卵黄囊, 位于体中央偏后,尚未开口于体外。孵化后5h的仔鱼, 消化管后方背面出现黑色素细胞, 鱼体靠近尾部出现两个上下对称的黑色素细胞。初孵仔鱼腹部朝上, 悬浮于水中,游泳能力较弱, 尾部偶尔快速扭动。

图2 七带石斑鱼仔、稚、幼鱼Fig.2 Larva, juvenile, and young fish of E.septemfasciatus
1日龄仔鱼 全长2.48 mm±0.030mm, 卵黄囊长径 0.70 mm±0.010 mm, 短径 0.24 mm±0.012 mm。油球1个, 圆球形, 油球直径0.14 mm±0.012 mm, 卵黄囊与油球逐渐缩小。脊索稍伸直, 头部增大, 大部分仔鱼头部已脱离卵黄囊并抬起。消化道开始膨大, 肛门位置略前移, 位于鱼体中央, 输尿管细小透明, 后端膨大为膀胱。仔鱼眼部晶体透明, 可清楚看到耳囊,耳囊内有 2个耳石, 很小, 前后排列。肌节呈“V”字形, 23~25对。油球边缘出现一圈黑色素, 卵黄囊前端和眼后方出现星状黑色素, 仔鱼背部也新出现黑色素细胞。仔鱼游泳能力增强。
2日龄仔鱼 全长2.41 mm±0.024 mm, 卵黄囊长径 0.29 mm±0.021 mm, 短径 0.16 mm±0.014 mm。油球直径0.09 mm±0.024 mm, 卵黄囊与油球进一步被吸收, 部分仔鱼卵黄囊明显萎缩。口裂已经形成, 但口不能活动, 未开口。消化管变粗, 前端膨大形成胃,肛门开口于体外。仍然存在卵黄囊的仔鱼, 卵黄囊前端的黑色素颜色加深, 肛门与尾中央脊索上方的黑色素呈辐射状扩散生长; 卵黄囊明显萎缩的仔鱼,眼囊出现黑色素, 部分仔鱼眼囊已布满黑色素。
3日龄仔鱼 全长2.64 mm±0.082 mm, 卵黄囊与油球全部消失或只剩下残余的油球。胸鳍出现, 长0.38 mm。尾基部的鳍膜开始向内凹陷。耳囊明显增大。仔鱼上颌短, 下颌长。消化道膨大变粗蠕动有力,肠道形成第一个肠曲。眼囊黑色素颜色加深, 部分仔鱼眼囊出现虹彩色素。消化道及其上方的体侧原有的黑色素向腹部下方扩散生长。背部和脊索上方的黑色素消失, 在肛门与尾端之间中央处的黑色素向脊索方向扩散生长。仔鱼开始由外源营养转换为内源营养。
4~5日龄仔鱼 全长2.69 mm±0.068 mm, 卵黄囊与油球完全被吸收。仔鱼口裂增大, 张合频率高, 在水中活力增强, 反应灵敏。在消化道进一步膨大, 可观察到大量的牡蛎 D形幼虫存在其中。心脏一心房一心室, 收缩有力。肛门与尾端中央处的黑色素更加密集, 眼囊中虹彩色素加深。仔鱼基本以外源营养为主。
6日龄仔鱼 全长2.74 mm±0.072 mm。消化道加粗, 摄食能力增强, 消化道中最多发现20个D形幼虫, 消化系统逐渐完善, 消化道前端的 D形幼虫呈黄色, 而 D形幼虫进入消化道后端时变得透明, 说明D形幼虫壳内的物质已经被消化吸收。鳃盖骨轮廓明显, 耳囊内耳石清晰可见。腹部色素加深, 脊索瓦状分节。尾椎下方鳍膜可见辐射状的鳍条丝。
7日龄仔鱼 全长2.74 mm±0.052 mm。尾部上方鳍膜也出现辐射状鳍条丝。消化道上方的树枝状黑色素增密, 肛门与尾端中央的黑色素扩散至脊索中央。尾部脊索下方出现数条长枝状黑色素。在显微镜下可清楚的分辨出动脉和静脉。D形幼虫可顺利的经肛门排出。
8日龄仔鱼 全长2.76 mm±0.096 mm。背鳍膜变窄变平并出现第二背棘原基, 但未超出鳍膜。尾鳍膜辐射状鳍条丝更加明显。消化系统发育更加完善, 消化道前部逐渐变得不透明, 看不清摄食的D形幼虫,幽门盲囊可见。背部鳍膜中新增一处向外辐射生长的点状黑色素, 个别仔鱼此处有两个点状黑色素。
9日龄仔鱼 全长3.08 mm±0.088 mm。第二背鳍棘超出鳍膜, 长0.13 mm。鳃盖骨明显, 头部各器官逐渐发育完善。心脏清晰可见。消化系统发达, 幽门盲囊清晰可见, 经解剖发现仔鱼已经摄食轮虫。尾鳍发达, 摆动有力, 摄食能力强, 尾鳍后缘也出现长枝状的黑色素。
10日龄仔鱼 全长3.47 mm±0.094 mm。仔鱼生长差异显著, 大小不均, 较大的仔鱼胸鳍发达, 尾鳍摆动有力, 反应迅速, 已能平游, 仔鱼在光线亮的地方容易集群。腹部黑色素覆盖面积增大, 几乎将整个腹部遮住。背部的点状黑色素继续向外生长。尾部下方的长枝状黑色素延长, 数量增多。
11日龄仔鱼 全长3.73 mm±0.098 mm。鱼体各组织器官发育迅速。下颌明显长于上颌, 胃肠蠕动有力, 心跳明显, 耳囊清晰可见。腹鳍棘长出, 长0.31mm, 腹鳍棘明显长于背鳍棘, 长棘的出现是石斑鱼属鱼类仔鱼发育所特有的。肛门与尾端中央的黑色素明显增多, 俯瞰鱼体几乎环抱此处整个肌体。
12日龄仔鱼 全长3.74 mm±0.086 mm。背、腹鳍棘增长, 腹鳍棘末端已超过肛门, 腹鳍棘明显长于背鳍棘, 长棘末端黑色素细胞增多, 腹部出现黄色素, 肛门与尾端中央的黑色素丛下方也出现 5~6条长枝状黑色素。尾鳍辐射状鳍条丝明显。
13日龄仔鱼 全长4.08 mm±0.074 mm。仔鱼生长差异显著, 最小个体全长只有2.90 mm, 最大的个体全长6.50 mm。鳃盖骨明显, 隐 约可见 鰓 丝。腹部点状黄色素细胞数量增多, 背腹长棘继续伸长, 长棘中部也开始出现少量的黑色素, 腹鳍棘前缘出现2列锯齿状的小棘, 背鳍长棘顶端出现一个无色透明的刺。头部上方及背鳍长棘前后的鳍膜明显变窄变平。仔鱼活动能力增强。
14日龄仔鱼 全长4.32 mm±0.462 mm。头部各组织发育逐渐完善, 耳囊内2个耳石前后排列, 前者小后者大。背、腹鳍长棘明显增长, 长棘上黑色素分布不均匀, 末端较密, 基部黑色素较少。长棘都长出数列锯齿状小棘并出现点状黄色素。第一背鳍棘长出, 长0.11 mm, 第二背鳍棘基部下方出现一丛黑色素细胞。腹部出现虹彩色素, 尾端下方的长枝状黑色素延长, 数量增加。
15日龄仔鱼 全长 4.44 mm±0.531mm。俯视仔鱼, 上颌中央有一明显向内凹入的缺刻, 长度短于下颌, 上颌齿隐约可见。鳃丝清晰可见略显红色。背、腹长棘继续增长, 末端稍膨大, 黑色素密集, 靠近基部处黑色素稀疏, 可见单个的树枝状黑色素, 长棘中下部点状黑色素增多。腹部丛状黑色素逐渐分散开, 黄色素加深, 已看不清消化道。背部向外辐射生长的点状黑色素消失, 仔鱼游动灵敏, 受到惊吓后三叉棘会迅速张开。
16日龄仔鱼 全长5.30 mm±0.681 mm。上颌与下颌长度差异不大。背鳍棘长度已超过全长 1/2, 背腹鳍膜继续缩小, 尾鳍与背鳍之间和尾鳍与臀鳍之间的鳍膜逐渐向内凹陷。背鳍长棘有 3列锯齿状的小棘, 前缘1列, 后缘2列; 腹鳍棘有4列小棘, 前缘2列, 后缘2列。第三背鳍棘超出鳍膜。仔鱼摄食能力明显增强。
17日龄仔鱼 全长5.78 mm±0.642 mm。仔鱼头部颜色加深, 心脏逐渐被发育的鳃盖骨遮盖, 只隐约可见。上颌上方出现黑色素, 腹部的黑色素开始消退, 只有消化道上方的黑色素较密集, 点状黄色素和蓝色素增多。鳔充气, 从鱼体侧面观察在贴近脊柱的下方有一小亮圈, 呈椭圆形。
18日龄仔鱼 全长6.19 mm±0.724 mm。头部黑色素增多, 出现少量虹彩色素, 只能隐约看到耳囊的轮廓。上颌齿清晰可见, 10颗, 透明, 排列稀疏。鳃盖上出现2枚棘。腹部下半部分黑色素已消退, 黄色素颜色加深, 鳔泡已不可见。肌节呈“V”形, 21对。尾部脊索末端上翘, 部分尾鳍鳍条出现。
19日龄仔鱼 全长6.62 mm±0.874 mm。头部黑色素加深, 眼上方出现 1枚棘, 明显看出鳃丝呈红色。背、腹鳍长棘继续增长, 第二背鳍棘已超过全长的2/3, 腹鳍接近全长的1/2, 第一背鳍棘后缘出现锯齿状小棘。尾鳍下方的长枝状黑色素逐渐消褪。
20日龄仔鱼 全长6.74 mm±0.883 mm。腹部黑色素减少, 虹彩色素增多。第二背棘基部下方黑色素增多。尾部鳍膜与背部、腹部鳍膜分开形成尾柄, 背鳍、臀鳍逐渐发育, 尾鳍鳍条数量增加, 脊索末端上翘, 尾下骨增大并连成片状, 中央向内凹陷。肌节呈“W”型, 22对。
21~25日龄仔鱼 全长7.65 mm±1.120 mm。随着鱼体的生长, 头部发育更加完善, 透光性差, 头部黑色素加深, 上颌齿仍为 10颗, 大小没有明显增大,下颌无齿, 长度与上颌相当。眼径增大, 眼囊充满蓝色素。眼眶上方出现一棘, 主鳃盖骨耳囊后方位置向后长出一短棘, 前鳃盖骨出现 3枚棘, 其外侧 2枚,另有一枚向下的短棘。鳃丝红色加深。消化道下部的树枝状黑色素基本消褪, 取而代之的是黄色素和虹彩色素, 肛门与尾中央仍有一丛黑色素。背腹鳍长棘继续增长, 略有弯曲, 第二背棘长度超过全长的3/4, 腹鳍棘长达到全长的 1/2, 各鳍鳍条清晰可见,尾柄末端尾骨分上下两部分, 中央凹入, 鳍条数为17。背腹鳍长棘的末端黑色素颜色深, 越靠近基部颜色越浅。第二背鳍棘前缘锯齿状小棘的数量多于后缘, 腹鳍棘前、后缘小棘的数量相差不大。
26~30日龄仔鱼 全长7.98 mm±1.461 mm。头部各器官逐渐发育完, 头部上方颅骨部位和耳囊后方出现几点星状的黑色素。鳍膜已基本消失, 只在尾柄的上下残留很窄的一点, 各鳍鳍棘和鳍条逐渐明显,第一背鳍出现10根鳍棘, 其中 6根超出鳍膜, 第三背鳍棘明显长于第一背鳍棘, 第二背鳍出现14根鳍条; 腹鳍1根棘4根鳍条; 臀鳍形成, 可见3根棘9根鳍条; 尾鳍鳍条更加明显, 鳍条分6节。尾端下方的长枝状黑色素全部消失。全长9.6 mm的仔鱼, 背、腹鳍长棘的长度分别为7.8、5.3 mm, 达到3根长棘绝对长度的最大值, 随着鱼体的生长发育, 背、腹鳍长棘开始逐渐被吸收, 仔鱼进入稚鱼去期。背、腹鳍长棘上的黑色素开始由基部向末端逐渐消褪, 第二背棘后缘小棘明显比前缘小棘大。肝门与尾鳍之间腹缘的树枝状黑色素后移至尾柄处。骨骼和肌肉系统发育增快, 躯体明显变宽, 脊椎骨明显, 尾椎下方出现红色的血管。肝脏与胆囊已形成。
2.2.2 稚鱼期
31~40日龄稚鱼 全长16.1 mm±1.862 mm。进入稚鱼期后, 鱼体生长速度明显变快, 大小差异显著,稚鱼骨骼和肌肉系统快速发育, 眼眶上方和鳃盖骨上的小棘消失, 颅骨上方的黑色素增多连成片状并开始大量集中。消化道上部的树枝状黑色素逐渐消褪, 只覆盖其 1/3, 腹部肌肉增厚, 随着生长腹部开始呈银白色反光, 摄食卤虫无节幼体后腹部明显膨胀, 呈现红棕色。背、腹鳍长棘继续收缩至已接近全长 1/4, 长棘基部只剩少量黑色素, 末端黑色素仍较密集。尾鳍棘9根, 鳍条15根; 腹鳍棘1根, 鳍条5根; 臀鳍棘3根, 鳍条9根; 尾鳍条17根, 尾骨中央向内凹陷处的上半部分8根, 下半部分9根, 尾鳍鳍条分节数增多。背鳍鳍条基部开始出现星状黑色素,尾柄处出现数枚细小栉鳞。稚鱼活动能力和摄食能力都很强, 反应灵敏, 鱼苗多在水的中下层活动。
41~50日龄稚鱼 全长19.5 mm±1.461mm。背、腹鳍长棘收缩明显, 长度分别为3.85、3.07 mm, 长棘后缘仍有几个较大的小齿。从眼睛上方至脊椎最前端覆盖了较密集的黑色素, 身体背部略有隆起,背部和脊柱上部黑色素细胞明显增多, 呈线状排列。腹部消化道上方的黑色素消褪明显, 只在腹腔上方有小部分星状黑色素分布, 腹部仍以黄色素数量最多, 消化道下方布满蓝色素。尾柄处有一黑色素丛,脊椎明显, 其下方出现一条红色的血管。各鳍逐渐发育完善, 臀鳍第2根鳍棘明显增粗。稚鱼活动能力强,摄食旺盛。
55日龄稚鱼 全长24.5 mm±1.383 mm。全长28.5 mm的稚鱼, 背、腹鳍长棘的绝对长度最小, 分别为2.23 、2.83 mm。各鳍鳍条与成鱼相似。体被小栉鳞,侧线发达, 体表色素急剧增加, 在体表形成数条横带。除体表横带外, 形态特征已和成鱼无多大差异。
2.2.3 幼鱼期
58日龄幼鱼 全长29.6 mm±1.962 mm。幼鱼完成变态, 细小栉鳞覆盖整个鱼体, 体表侧线发达, 7条横带明显。第二背鳍棘和腹鳍棘已完全收缩。除生殖腺尚未发育成熟外, 幼鱼的形态特征和生活习性基本接近成鱼, 幼鱼喜欢栖息在育苗池底。
2.3 仔稚幼鱼的生长与培育管理
七带石斑鱼仔、稚、幼鱼全长及鳍棘长度与日龄的关系如图3所示。

图3 七带石斑鱼仔、稚、幼鱼的生长Fig.3 Growth of larva, juvenile, and young fish of E.septemfasciatus
2.3.1 前期准备
小球藻和金藻(Chrysophyta)培养: 为调节培育池水色、维持培育水体的微生态环境, 购置小球藻、金藻(3011)20 000 mL进行保种室培育、5~6 d后扩种入0.4 m3塑料桶中使用连续充气法培养。7 d后进入二级培养池(4 m3)5~6 d, 后接入三级培养池(15 m3)3 d, 此时所培育藻密度≥300个/mL。
育苗池消毒: 使用漂白粉消毒、刷洗池底和池壁,冲洗3~4遍后用硫代硫酸钠中和残余氯离子。
培育水环境: 布池前加入升温后海水, 水温 24℃,水深80 cm, 光强度1 500 ~2 000 lx, 每个仔鱼培育池加入密度≥300 个/mL的小球藻0.5 m3。
2.3.2 培育过程
七带石斑鱼受精卵的布池密度为4 000~6 000粒/m3。
仔鱼从孵出至 3日龄, 即自孵化出膜至开口的时期, 此阶段不换水、不投饵。应注意的是此时期需要严格控制充气量, 防止水体剧烈波动对仔鱼造成机械损伤。
仔鱼从 4日龄开始摄食, 此时水温逐渐升高到26 ℃。以长牡蛎(Ostrea gigas)受精卵及幼体作为开口饵料。长牡蛎受精卵制备方法: 将活体牡蛎逐个洗净、开壳、检查雌雄, 以雌雄比例10: 1, 用120目尼龙滤网搓碎转入干净容器中, 加海水静置5分钟, 再滤去浮沫及大型组织碎块, 按预定投喂量均匀泼洒于仔鱼培育池, 投喂原则为“少投勤投”(24 ℃水温下牡蛎受精卵22 h左右发育成为D形幼虫, 因其具有几丁质外壳故无法被仔鱼消化吸收)。保持育苗池中牡蛎受精卵的密度为0.5~1个/mL。镜检观察肠道内充满牡蛎受精卵及少量 D形幼虫, 则表明仔鱼摄食良好。每天加升温海水10 cm并补充小球藻保持水色。夜间开启暖风空调以维持室温恒定, 控制育苗池水温稳定。仔鱼 6日龄后逐渐停止投喂牡蛎受精卵, 开始投喂S型轮虫。从9日龄开始, 仔鱼第二背鳍棘和腹鳍棘长出, 鳍棘较脆弱易折断, 因此, 在鳍棘长出之前, 应预先调整育苗池中仔鱼密度, 避免密度过大仔鱼因趋光聚集而导致鳍棘碰撞引起死亡。
10日龄后, 仔鱼开始摄食 L型轮虫, 但摄食量较低, 镜检发现幼鱼肠道中较多 D形幼虫。因七带石斑鱼仔鱼消化道纤细, 单尾仔鱼摄食野生轮虫量1~3个, 多数仅摄食1~2个轮虫。轮虫投喂量过大的弊端: 若环境不适, 轮虫大量死亡, 仔鱼无可摄食饵料导致饥饿死亡; 若条件适当, 轮虫大量繁殖, 造成水质变差, 导致仔鱼大量死亡。所以, 要严格控制轮虫投喂数量, 定时抽样检查育苗池的轮虫密度, 维持在 5个/mL并根据仔鱼的摄食情况调整轮虫的投喂量。从11日龄开始使用80目网箱或滤鼓换水, 日换水量10%。
15~20日龄仔鱼进入开鳔期, 此时背腹棘较长,仔鱼趋光现象明显, 经常聚集于培育池较明亮的角落。此期关键点: 注意水质变化, 控制水体透明度30~40 cm; 适宜光照强度范围1 000 ~1 500 lx, 避免强光、直射光, 增加小球藻浓度以加深水色, 培育车间窗及棚顶加盖遮光网以降低日间光照。发育至此时期的仔鱼易受惊扰, 应减少惊动, 尤其是直射光、强光手电和巨大的声响会导致仔鱼严重受惊吓而死亡。当仔鱼发育到17日龄时, 第二背鳍棘和腹棘已迅速增长, 活动能力增强, , 开始少量投喂卤虫无节幼体。保持育苗池中卤虫无节幼体的密度为0.1个/mL,轮虫密度降至1个/mL。26日龄时停止投喂轮虫, 卤虫无节幼体的密度增至0.5~1个/mL。室内水泥池育苗时七带石斑鱼苗以为卤虫无节幼体主要食物的时间较长, 一直要持续到仔鱼期结束。此时, 很容易用肉眼观察到鱼的摄食情况, 侧面观察鱼体腹部呈红褐色, 实际操作中可以根据仔鱼摄食量来增减饵料的投喂量。日换水量20%。
55日龄后幼鱼全长超过2.5 cm, 开始投喂鱼虾肉糜并每天吸底1次。此时期互残现象较为严重, 个体较大的幼鱼大多会被个体较小的幼鱼噎死, 从而大大降低苗种培育成活率。为避免残食现象的发生,间隔5~7 d分选1次; 提高投喂频率, 在育苗池中设置掩蔽物, 如空心砖、塑料管等, 也可有效减少互相残食。
3 讨论与小结
3.1 胚胎发育
七带石斑鱼受精卵径 0.854 mm ±0.021mm, 比谢菁等[14]报道的(0.8880 mm ±0.024 mm)稍小, 比同属的鞍带石斑鱼(Epinephelus lanceolatus) (0.79 mm±0.85mm)[15]、点带石斑鱼(Epinephelus malabaricus)(0.75 mm±077 mm)[16]大, 比云纹石斑鱼(Epinephelus moara) (0.871 mm±0.013 mm)[17]小。本研究发现七带石斑鱼受精卵约 2640粒/g, 而点带石斑鱼[16]、云纹石斑鱼[18]的受精卵约 1500粒/g, 作者认为可能是七带石斑鱼卵黄较小, 卵内含物质较轻所致。另外, 亲鱼的个体大小、培育环境的不同、饵料营养的优劣、产卵方式等都会造成受精卵卵径的差异, 石斑鱼是分批产卵, 所以同一尾亲鱼排出的卵粒大小也会有差异。本研究中胚胎孵化出膜时间为31 h5 min, 赵明等[18]报道的在温度(22±0.5)℃, 盐度30~35范围内胚胎孵化出膜时间为32 h36 min~32 h42 min, 作者认为可能是本研究提高了受精卵的孵化水温导致胚胎孵化出膜的时间缩短。根据实验观察的结果, 将七带石斑鱼的胚胎发育划分为卵裂期、囊胚期、原肠胚期、神经胚期和器官形成期, 与报道的其他石斑鱼类[15-17,19]结果基本一致。
3.2 七带石斑鱼胚后发育阶段的划分
参照国内外石斑鱼仔稚幼鱼的划分方式[20-23],七带石斑鱼胚后发育主要依据卵黄囊的变化、长鳍棘的生长与收缩、鳞片的出现划分为仔鱼期、稚鱼期、幼鱼期。仔鱼期分为前期仔鱼和后期仔鱼, 前期仔鱼主要由卵黄囊提供营养, 为内源性营养阶段;后期仔鱼阶段, 仔鱼完成从内源性营养到外源性营养的过渡, 仔鱼在形态、生理方面变化剧烈。国内外学者在后期仔鱼结束的标志上观点有所不同, 郭仁湘等[20]认为以仔鱼各鳍基本发育形成作为鞍带石斑鱼后期仔鱼结束的标志; 邹记兴等[23]认为点带石斑鱼仔鱼背腹鳍棘开始收缩, 鳞片长出前为后期仔鱼结束的标志。本研究表明, 30 dph的七带石斑仔鱼第二背鳍棘和腹鳍棘的绝对长度达到早期发育阶段的最大值。根据七带石斑鱼胚后发育的特性, 将仔鱼长鳍棘绝对长度最大作为后期仔鱼结束的标志, 这与邹记兴等[23]观点一致。稚鱼期为长鳍棘开始收缩至鳞被完全形成结束。幼鱼期, 鱼体全身被鳞片覆盖,横带明显, 除生殖腺尚未发育成熟外, 幼鱼的形态特征和生活习性基本接近成鱼。
3.3 七带石斑鱼仔、稚、幼鱼的生长与培育
本研究测得七带石斑初孵仔鱼的全长为1.99 mm±0.041mm, 比陈超等[24]报道的 1.36 mm±0.07 mm 偏大, 周玲等[15]在对鞍带石斑鱼胚胎发育及仔鱼形态发育观察研究中认为, 可能跟受精卵的卵粒大小和卵黄含量的差异有关; 王涵生等[25]在对赤点石斑鱼仔、稚、幼鱼研究中认为这可能是地理种群差别或取样个体、时间差异所致; 作者认为初次产卵的亲鱼得到受精卵卵径比较大, 卵黄含量较高, 因此亲鱼的年龄和体质的不同, 会造成初孵仔鱼大小的差异,具体原因还有待进一步研究。
前期仔鱼阶段(0~3 dph), 鱼苗发育所需营养主要由卵黄囊提供, 个体差别较小。后期仔鱼阶段(4~30dph), 鱼苗由内源性营养转换为外源性营养,由于卵质不良、先天性畸形、不能开口摄食、开鳔等原因造成这期间鱼苗的成活率很低, 另外长鳍棘的长出和趋光性会使鱼苗过度密集, 窒息致死。饵料生物的种类、大小、数量及其某些营养物质的缺少或不足是导致仔稚鱼大量死亡的一个重要因素[15]。人工培养的轮虫及孵化的卤虫无节幼体, 由于缺乏高度不饱和脂肪酸, 连续使用会造成仔稚鱼大量死亡。实践证明, 经过几天藻类培育及营养强化的轮虫或卤虫无节幼体是比较安全的生物饵料[26]。因此在育苗中应及时投喂牡蛎受精卵作为开口饵料, 并根据鱼苗的发育状况完成由牡蛎受精卵到S型轮虫、L型轮虫、卤虫无节幼体的饵料转换。本研究用小球藻和金藻强化轮虫, 用裂壶藻(Schizochytrium limacinum)强化卤虫无节幼体, 有效提高了鱼苗的成活率。每次更换饵料, 要有2~3 d的过渡时间, 以便多数鱼苗能很好地适应新饵料; 更换饵料要适时, 太迟影响鱼的生长, 太早则会造成个别能摄食较大饵料的鱼苗长得特别快, 引起生长不均匀。尽量保持水质稳定, 水色保持淡绿色为佳, 夜间育苗池边不开灯,在鱼苗聚集处放置充气石充氧, 防止鱼苗窒息死亡。
稚鱼期(31~57 dph)最显著的特征是长鳍棘的收缩, 鱼苗转为底栖生活, 摄食饵料由卤虫转换为鱼虾肉糜, 已经出现相互攻击和残食现象。在云纹石斑仔鱼发育出现三叉棘到变态成稚鱼这段时期, 仔稚鱼要经历各种形态学、生理学的变化, 对外界环境的变化十分敏感, 处理不当会导致仔稚鱼的大批量死亡[18]。因此, 要避免各种人为操作的刺激, 如温差变化、强光照射、肆意捞捕等[27]。长鳍棘收缩阶段是石斑鱼苗种培育的关键时期, 虽然长鳍棘的作用机理尚不清楚, 但在这一时期加强培育管理, 保持水环境稳定, 适时筛选分池, 提高喂食频率, 设置掩体,避免相互残食, 可以有效降低苗种死亡率。直至进入幼鱼期(58 dph)长鳍棘完成收缩, 鱼苗的生长速度加快, 全长超过6 cm后残食明显减弱。
在七带石斑鱼仔、稚、幼鱼的生长速度方面, 不同的报道间具有一定差距, 本研究30 dph仔鱼全长7.98 mm±1.461mm, 比陈超等[24]报道的 11.08 mm±0.792 mm 偏小。60 dph的幼鱼全长 30.60 mm±1.961mm, 与北島力等[28]报道的 60 dph全长即可超过30 mm相差无几。作者认为其中的原因是多方面的, 饵料质量、培育池的大小、培育水温(陈超等培育池为25 m3, 水温22~24℃; 本研究培育池为15m3,水温 22~28℃)、布池密度等都会造成仔稚、幼、鱼生长速度的差异。
将本研究结果与鞍带石斑鱼[20]、赤点石斑鱼(E.akaara)[25]、斜带石斑鱼(E.coioides)[29]、云纹石斑鱼[17]早期发育进行比较(表2), 七带石斑鱼胚后发育较慢, 背腹鳍棘较长等特点增加了人工繁育的难度, 成为实现规模化繁育的瓶颈, 因此还有待加强其苗种规模化繁育技术的研究。
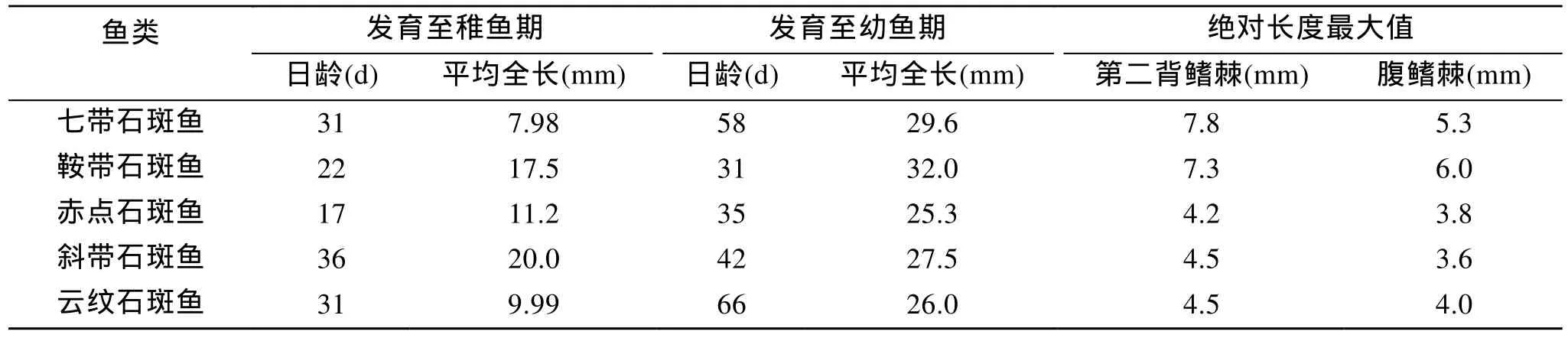
表2 七带石斑鱼与其他4种石斑鱼的相关数据对比Tab.2 Comparison of relevant data of Epinephelus septemfasciatus to those of other groupers
[1]区又君, 廖光勇, 陈超, 等.七带石斑鱼精子活力及其与环境的关系[J].海洋环境科学, 2011, 30(4): 516-519.
[2]王新安, 马爱军, 陈超, 等.七带石斑鱼两个野生群体形态差异分析[J].海洋与湖沼, 2008, 39(6) : 655-660.
[3]Kline R J, Khan I A, Soyano K, et al.Role of folliclestimulating hormone and androgens on the sexual inversion of seven-band grouper Epinephelus septemfasciatus [J].North Am J Aquac, 2008, 70(2) : 266-272.
[4]Koh I C C, Yoloi K, Tsuji M, et al.Cryopreservation of sperm from seven-band grouper,Epinephelus septemfasciatus[J].Cryobiology, 2010, 61(3) : 263-267.
[5]廖光勇, 区又君, 李加儿, 等.七带石斑鱼精子的超微结构[J].上海海洋大学学报, 2011, 20(1) : 71-75.
[6]雷从改, 尹绍武, 陈国华.石斑鱼繁殖生物学和人工繁殖技术研究现状[J].海南大学学报(自然科学版),2005, 23(3): 75-79.
[7]刘新富, 庄志猛, 孟振, 等.七带石斑鱼人工繁育技术研究进展[J].中国水产科学, 2010, 7(5): 1128-1136.
[8]Teruya K, Yoseda K, Oka M, et al.Effects of photoperiod on survival, growth and feeding of seven band grouper Epinephelus septemfaciatus larvae[J].Nippon Suisan Gakkaishi, 2008, 74(4) : 645-652.
[9]Kokawa Y, Takami I, Nishizawa T.A mixed infection in seven band grouper Epinephelus septemfasciatus affected with viral nervous necrosis (VNN)[J].Aquaculture, 2008, 284(1/2/3/4) : 41-45.
[10]陈超, 程波, 于宏, 等.七带石斑鱼繁殖群体“突眼”症病原菌的分离与鉴定[J].渔业科学进展, 2010, 31(1):25-33.
[11]Tanaka S, Takagi M, Miyazaki T.Histopathological studies on viral nervous necrosis of seven band grouper,Epinephelus septemfasciatusThunberg, at the grow-out stage[J].J Fish Dis, 2004, 27(7) : 385-399.
[12]程波, 陈超, 王印庚, 等.七带石斑鱼肌肉营养成分分析与品质评价[J].渔业科学进展, 2009, 30(5) : 51-57.
[13]钟声平, 陈超, 王军, 等.七带石斑鱼染色体核型研究[J].中国水产科学, 2010, 17(1) : 150-155.
[14]谢菁, 区又君, 李加儿, 等.七带石斑鱼胚体和卵黄囊期仔鱼的发育[J].海洋通报, 2009, 28(2): 41-49.
[15]周玲, 翁文明, 李金亮, 等.鞍带石斑鱼胚胎发育及仔鱼形态发育、饵料转变的观察研究[J].中国农学通报, 2010, 26(01): 293-302.
[16]邹记兴, 常林, 向文洲, 等.点带石斑鱼的亲鱼培育、产卵受精和胚胎发育[J].水生生物学报, 2003, 27(4):378-384.
[17]宋振鑫, 陈超, 翟介明, 等.云纹石斑鱼胚胎发育及仔、稚、幼鱼形态观察[J].渔业科学进展, 2012, 33(3): 26-34.
[18]赵明, 陈超, 柳学周, 等.盐度对七带石斑鱼胚胎发育和卵黄囊仔鱼生长的影响[J].渔业科学进展, 2011,32(2): 16-21.
[19]贾瑞锦, 王鲁, 赵从明, 等.条纹锯鱼旨胚胎发育及卵黄囊仔鱼形态变化的观察[J].渔业科学进展, 2012,33(3): 26-34.
[20]郭仁湘, 符书源, 杨薇, 等.鞍带石斑鱼仔稚(幼)鱼的发育和生长研究[J].水产养殖, 2011, 32(4) : 8-13.
[21]许波涛, 李加儿, 周宏团.赤点石斑鱼的胚胎和仔鱼形态发育[J].水产学报, 1985, 9(4): 369-374.
[22]陈国华, 张本.点带石斑鱼仔、稚、幼鱼的形态观察[J].海南大学学报(自然科学版), 2001, 19(2): 151-156.
[23]邹记兴, 向文洲, 胡超群, 等.点带石斑鱼仔、稚、幼鱼的生长与发育[J].高技术通讯, 2003, 4: 77-84.
[24]陈超, 赵明, 柳学周, 等.七带石斑鱼胚胎及仔稚鱼形态观察[J].渔业科学进展, 2011, 32(5): 24-31.
[25]王涵生, 方琼珊, 郑乐云.赤点石斑鱼仔稚幼鱼的形态发育和生长[J].上海水产大学学报, 2001, 10(4):307-312.
[26]苏惠美.石斑, 石斑鱼饵料生物培养与利用[M].基隆:台湾渔业经济发展协会, 2007: 70-103.
[27]刘秉忠.石斑, 石斑鱼养殖要点[M].基隆: 台湾渔业经济发展协会, 2007: 38-47.
[28]北島力, 高屋雅生, 塚島康生, 等.マ ハタの卵内発生および 飼 育による仔稚 魚 の 形 態 態化[J].魚魚学雑誌, 1991, 38(1): 47-55.
[29]刘冬娥, 张雅芝, 方琼珊, 等.斜带石斑鱼仔、稚、幼鱼的形态发育研究[J].台湾海峡, 2008, 27(2): 188-189.
