㮈叶片乙醇酸氧化酶基因全长cDNA 的分离与表达研究
2014-12-14陈桂信潘东明王玉珍吕恃衡姜翠翠
赵 利,陈桂信*,潘东明*,王玉珍,吕恃衡,姜翠翠
(1.福建农林大学 园艺学院,福建 福州 350002;2.福建农林大学 园艺产品贮运保鲜研究所,福建 福州 350002;3.福建省农业科学院 果树研究所,福建 福州 350013)
乙醇酸氧化酶是植物光呼吸代谢的一种关键酶,催化羟乙酸盐氧化成乙醛酸盐同时生成等分子量的H2O2[1]。有报道[2]表明过氧化氢参与信号传导和具有诱导抗逆性,它的快速积累是非病原性信号早期超敏反应的一个特点。GLO 可能与植物的抗病性有关,通过转基因过量表达GLO 基因,可以增强植物的广谱抗病性[4]。从菠菜中分离到GLO 基因,通过农杆菌介导法导入苹果,获得了2 个转GLO 基因的抗性芽[4],水稻[5]等植物上面均有研究。利用蛋白质表达谱比较抗性和易感性的体细胞杂交,表明GLO 在抗病植物中含量丰富,在欧洲油菜和拟南芥之间的体细胞杂交种产生了对黑胫病的显著抗性[6]。同时,在大麦[7]、甜瓜[3]中与致病性真菌接种,其GLO 酶活性增强。此外,GLO 在植物的应激反应中有重要的作用。GLO 酶的活性在各种环境胁迫中的诱导已被证实,如干旱胁迫已在豇豆、豌豆和烟草中得到证实[8-10]。GLO 在植物受到病原微生物侵染后引发的一系列防御反应中起着非常重要的作用,显示出在基因工程抗性育种研究中的广阔应用前景。研究表明,近几年来在玉米[11]、水稻[12]、春兰[13]等植物上克隆出了GLO 基因。GLO 为多基因家族,水稻中发现了5 个GLO 基因[14],但是在果树上,尤其是蔷薇科李属,仅碧桃上克隆到部分GLO 基因序列,其它树种未见报道。
㮈(Prunus salicina Lindl.var.cordata J.Y.Zhang et al.)为蔷薇科李属,桃形李实,是福建省名、特、优的核果类果树,生产上㮈常遭受流胶病、细菌性穿孔病、白粉病、缩叶病等病害[15]的侵袭和危害,严重影响树势、产量和果实品质,甚至缩短树体的生产寿命。本研究在已构建好的㮈叶片cDNA 文库基础上,利用96 孔板法从㮈叶片cDNA 文库中分离获得一个GLO 全长基因,对该基因进行了生物信息学分析及在㮈叶片5 个不同生长发育时期的表达模式分析,为进一步分析该基因在㮈的抗逆性和病虫害的防御过程中提供理论依据。
1 材料和方法
1.1 植物材料与试剂
㮈 叶芽(2012 年2 月)、展开叶(2012 年3 月初)、幼叶(2012 年4 月中旬)、成熟叶(2012 年5 月中旬)、老叶(2012 年8 月中旬)采自福建宁德周宁㮈果园。将上述样品迅速冻存于液氮中,放于-80 ℃。
PrimerScriptTMRT reagent Kit、2×SYBR Premix ExTaqTM、试验所用的各种工具酶、dNTP 购自大连宝生物公司;大提质粒试剂盒购自Qiagen 公司;引物合成与测序委托上海铂尚生物技术公司完成。
1.2 cDNA 文库的构建
本研究以㮈(Prunus salicina Lindl.var.cordata J.Y.Zhang et al.)5 个不同生长发育时期叶片为材料提取总RNA,然后将这些总RNA 按相同比例混合,采用郑鸿昌[16]的方法构建㮈均一化全长cDNA文库,其该文库是以pBluescriptⅡKS(+)为载体,原始文库的滴度为2.0×108pfu/mL,其重组率为98%,插入片段大小为1 800 bp 左右,文库质量良好。
1.3 㮈叶片GLO 基因的分离与序列分析
根据NCBI 上不同物种的GLO 序列,利用DNAMAN 软件设计简并引物,委托铂尚生物技术(上海)有限公司合成。GLOF:5'-GGTKTATGACTACTATGCWTCTGGTGC-3';GLOR:5'-GCATCCTCWGCTGTAAKHACACCCTT-3'。用通用引物验证筛选片段大小,M13R:5'-GGAAACAGCTATGACCATG-3',M13F:5'-GTAAAACGACGCCAGT-3'。采用基于PCR 技术的96 孔板法进行cDNA 文库的筛选[17],PCR 反应程序为:95 ℃5 min,95 ℃12 s,58 ℃30 s,72 ℃53 s,共35 个循环,72 ℃10 min,4 ℃pause。根据目标克隆在文库中的丰度取适量稀释度的文库用DH10B 菌株铺于96 板,建立一级库,每孔大约有40 个克隆,放于培养箱中培养8~10 h,利用PCR 反应,进行阳性克隆的筛选,将筛到的阳性孔再次进行克隆的稀释,建立二级库,每孔大约有6 个阳性克隆,操作步骤同上。将二级库中筛到的阳性克隆铺于含有AMP 抗性的LB 平板上培养过夜,挑单克隆进行PCR 鉴定,以确定阳性单克隆。
将得到的阳性克隆送至铂尚生物技术(上海)有限公司测序,利用DNAMAN 软件以及http://www.ncbi.nlm.nih.gov/等网站提供的各类生物信息学软件对序列进行在线分析。利用ProtParam 程序(http://web.expasy.org/protparam/)预测蛋白质的分子量和等电点;利用MEGA5.2 做进化树分析。
1.4 㮈叶片GLO 基因在㮈不同生长发育时期的表达分析
参照TaKaRa 公司PrimerScriptTMRT reagent Kit 逆转录成cDNA,将反转录的单链cDNA 稀释10 倍,取1 μL 作为模板,以㮈tubuin 基因作为内参基因,参照2×SYBR Premix ExTaqTM(TaKaRa,大连)荧光定量试剂盒说明书进行荧光定量试验。内参基因引物为Nai-tubuinF:5'-TCGGATGATGATGACCTTCTCTGTG-3';Nai-tubuinR:5'-CATACACTCGTCTGCGTTCTCCA-3'。目的基因引物分别是上游:5'-TGAGGCTGGTGTAAGGAA-3',下游:5'-GCGAACGACATCCACTTA-3'。
2 结果与分析
2.1 㮈叶片GLO 基因的分离与序列分析
参考其他植物GLO 基因的保守序列设计简并引物,建立二级库,经2 轮96 孔板筛选以及DH10B菌株平板上单克隆的鉴定,获得GLO 基因cDNA 阳性单克隆(图1,A)。用通用引物M13F 和M13R 对上述的单克隆进行PCR 验证,获得其片段大小为1 500 bp 左右(图1,B)。测序结果(图2)表明:该阳性克隆为1 531 bp,开放阅读框为1 122 bp,编码374 个氨基酸,包括一个起始密码子(ATG)和一个终止密码子(TAA)。GLO 的分子量为126 024.8;理论等电点为5.0;不稳定指数为40.95,为不稳定蛋白。脂肪系数为26.52;其疏水性平均值(GRAVY)为0.688;分子为C4626H7723N1531O1956S310,总原子数为16 146,该基因命名为PsGLO。

图1 㮈PsGLO 不同引物扩增片段电泳检测结果Fig.1 Detection by agarose electrophoresis of PsGLO amplified from the Prunus salicina with different primers
2.2 PsGLO 基因编码的氨基酸序列同源性比对及其进化分析
在NCBI 上进行PsGLO 蛋白比对,该蛋白与其他植物的GLO 蛋白具有很高的保守性,编码的蛋白属于辅基FMN-α 羟基酸氧化酶的同源家族蛋白,此蛋白家族主要存在于原核生物和真核生物中,这个家族的成员主要包括黄色蛋白(FCB2)、乙醇酸氧化酶(GLO)、乳酸单氧酶(LMO)、扁桃酸脱氢酶(MDH)和长链羟基酸氧化酶(CHAO),其中,乙醇酸氧化酶(GLO)存在于绿色植物中,是光呼吸过程中的关键酶之一。乙醇酸氧化酶(GLO)是辅基FMN-α 羟基酸氧化酶的同源家族蛋白之一,近几年来国内外一些研究表明,FMN 诱导植物抗病能力增强与H2O2积累,苯基丙酸类合成途径的激活以及酚类物质的积累是密切相关的[18],由此,可知乙醇酸氧化酶(GLO)也参与其中。利用MEGA5.2 软件最大似然法,将PsGLO 预测蛋白与GenBank 上登录的不同物种的GLO 进行系统进化树分析,图3 结果表明,PsGLO 与陆地棉Gossypium hirsutum(AEX09184.1)、大豆Glycine max(NP_001238412.1)、菜豆Phaseolus vulgaris(AGV54360.1)的亲缘关系相对较近,陆地棉、大豆、菜豆都属于草本双子叶植物,在木本植物中没有克隆该基因。

图2 PsGLO 基因cDNA 全长核苷酸序列和推测氨基酸序列Fig.2 Nucleotide sequence of PsGLO Cdna and deduced amino acid sequence
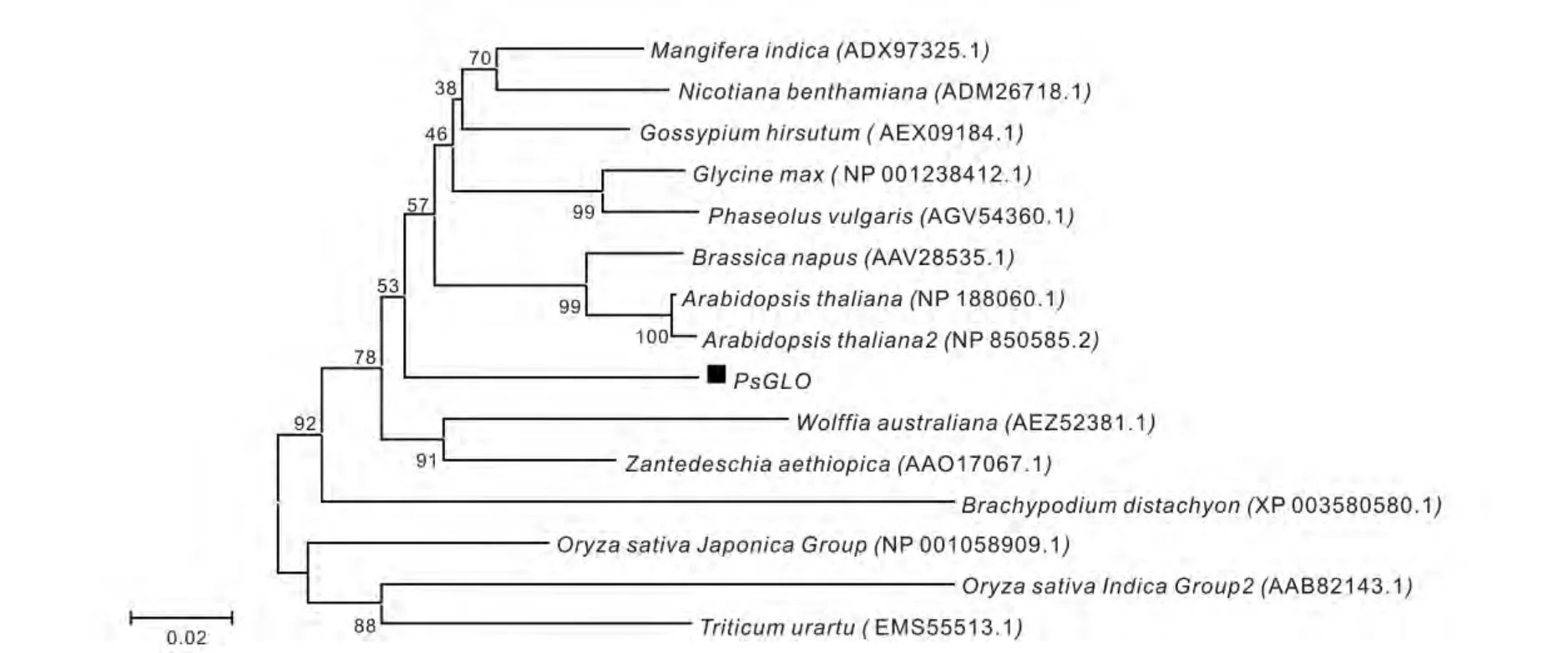
图3 㮈PsGLO 蛋白与其他物种GLO 蛋白进化树分析Fig.3 Phylogenetic analysis of PsGLO from the Prunus salicina and other plants
2.3 PsGLO 基因在㮈叶片不同生长发育时期的表达模式分析
利用实时荧光定量PCR 技术分析PsGLO基因㮈叶片不同生长发育时期的表达模式,图4 结果表明,PsGLO 基因在叶片不同生长发育都有表达,其中在老叶中表达量最高,叶芽中表达量最低,在植物辣椒上最早在1989 年开始研究植物的“年龄相关的抗性”(age-related resistance,ARR)[19],植物上人们把当年老的植株或叶片表现出来的对病害增强或降低的抗性通常就被叫为年龄相关的抗性[20],由此,可以推测PsGLO 基因㮈叶片叶生长发育过程对病害的抗性起着一定的作用,在老叶中PsGLO 的表达量相对其它4 个时期高。

图4 PsGLO 基因在㮈叶片不同生长时期的表达分析Fig.4 Expression of PsGLO gene during leaf of different stages in the Prunus salicina
3 结论与讨论
近几年来国内外研究结果表明,光呼吸是植物长期进化过程中,为了适应环境变化,提高抗逆性而形成光呼吸途径[1],乙醇酸氧化酶(GLO)是光呼吸过程中的关键酶,在植物受到病原微生物的侵染后引发一系列的防御反应中起着非常重要的作用,显示出其在基因工程抗性育种研究中的广阔应用前景。研究表明,在辐照光下,转基因烟草的乙醇酸氧化酶活性降低到阈值一下,可以提高其光抑制作用,增加植物的抗性[21]。在国内,将在菠菜中获得的GLO 基因,通过根癌农杆菌介导将GLO 基因导入苹果,获得两个抗性芽[4]。
本实验从构建的㮈叶片5 个不同时期的RNA,然后将这些总RNA 按相同比例混合,采用郑鸿昌[16]的方法构建㮈均一化全长cDNA 文库,首次成功分离了木本植物中的乙醇酸氧化酶(GLO)基因的全长序列,填补了木本植物中GLO 基因克隆研究的空白。虽然其确切的生物学功能还有待于进一步的试验验证,但是该基因全长序列的获得为深入认识其生物学功能和了解乙醇酸合成成途径奠定了基础。通过已得到的PsGLO 基因序列,在NCBI 上进行氨基酸序列比对,利用MEGA5.2 构建进化树,得到该基因与草本双子叶植物陆地棉亲缘关系较近。因为在木本植物中,没有人对其进行研究,未发现木本植物中的乙醇酸氧化酶(GLO)的氨基酸序列[22]。本试验中实时荧光定量PCR 结果显示,在㮈的叶片中5 个不同生长发育阶段都有表达,其中,在老叶中表达量最高。根据”年龄相关的抗性”(age-related resistance,ARR)的理论基础,前人在不同的植物-病原体系中对植物年龄与病害抗性(age-related resistance,ARR)的研究由来已久[23-27]。研究表明,随着植物年龄的增长,对有些病害表现的更敏感,如在水稻种,老叶片对Xanthomonas campestris pv Oryzae 以及Pyricularia oryzae 的抗性比年轻叶片抗性强。类似的,成年烟草对Peronospora tobacina 的抗性比年轻的植株要强。有荧光定量的结果可知,乙醇酸氧化酶(GLO)在叶片不断生长发育过程中有叶芽到展开叶不断增加,在幼叶中相对前面两个时期也少一些,成熟叶和老叶相对于前面的3 个时期要高,但是,老叶中最高。由此,可以推测在生长过程中老叶对病害的抗性要比年轻的叶片抗性要强。
[1]Foyer C H,Bloom A J,Queval G,et al.Photorespiratory metabolism:genes,mutants,energetics,and redox signaling[J].Annual Review of Plant Biology,2009,60(1):455-484.
[2]Lamb C,Dixon R A.The oxidative burst in plant disease resistance[J].Annual Review of Plant Biology,1997,48(1):251-275.
[3]Taler D,Galperin M,Benjamin I,et al.Plant R genes that encode photorespiratory enzymes confer resistance against disease[J].The Plant Cell Online,2004,16(1):172-184.
[4]张军科,洪小娟,马锋旺.乙醇酸氧化酶基因植物表达载体构建及转化苹果的研究[J].华北农学报,2009(6):33-37.
[5]胥华伟,张改娜,侯典云.乙醇酸氧化酶基因超表达载体的构建与水稻的遗传转化[J].广东农业科学,2010(8):188-190.
[6]Bohman S,Wang M,Dixelius C.Arabidopsis thaliana-derived resistance against Leptosphaeria maculans in a Brassica napus genomic background[J].Theoretical and Applied Genetics,2002,105(4):498-504.
[7]Schäfer P,Hückelhoven R,Kogel K-H.The white barley mutant albostrians shows a supersusceptible but symptomless interaction phenotype with the hemibiotrophic fungus Bipolaris sorokiniana[J].Molecular Plant-Microbe Interactions,2004,17(4):366-373.
[8]Mittler R,Zilinskas B A.Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought[J].The Plant Journal,1994,5(3):397-405.
[9]Mukherjee S,Choudhuri M.Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings[J].Physiologia Plantarum,1983,58(2):166-170.
[10]Rizhsky L,Liang H,Mittler R.The combined effect of drought stress and heat shock on gene expression in tobacco[J].Plant Physiology,2002,130(3):1143-1151.
[11]Zelitch I,Schultes N P,Peterson R B,et al.High glycolate oxidase activity is required for survival of maize in normal air[J].Plant Physiology,2009,149(1):195-204.
[12]Lu Y,Li Y,Yang Q,et al.Suppression of glycolate oxidase causes glyoxylate accumulation that inhibits photosynthesis through deactivating Rubisco in rice[J].Physiologia Plantarum,2013,149(1):194-203.
[13]向林,李伯钧,秦德辉.春兰GLO 基因的克隆和实时定量表达分析[J].浙江农业学报,2011(3):517-522.
[14]Zhang Z,Lu Y,Zhai L,et al.Glycolate oxidase isozymes are coordinately controlled by GLO1 and GLO4 in rice[J].PloS one,2012,7(6):639-658.
[15]黄月淑.油㮈 主要病虫害的发生与防治[J].植物医生,2001,14(6):25-25.
[16]郑鸿昌.夜来香均一化全长cDNA 文库的构建与花香相关基因的遗传转化[D].福州:福建农林大学,2012.
[17]Yim Y S,Moak P,Sanchez V H,et al.A BAC pooling strategy combined with PCR-based screenings in a large,highly repetitive genome enables integration of the maize genetic and physical maps[J].BMC Genomics,2007,8(1):47-49.
[18]Taheri P,Tarighi S.A survey on basal resistance and riboflavin-induced defense responses of sugar beet against Rhizoctonia solani[J].Journal of Plant Physiology,2011,168(10):1114-1122.
[19]Kingkun D,Taeyangkun H,Hongsanho J.Expression of age-related resistance in pepper plants infected with Phytophthora capsici[J].Plant Disease,1989:745-750.
[20]邓本良.核黄素及核黄素结合蛋白对拟南芥抗病抗逆调控作用的初步研究[D].南京:南京农业大学,2012.
[21]Chern M,Bai W,Chen X,et al.Reduced expression of glycolate oxidase leads to enhanced disease resistance in rice[J].Peer J,2013,10(2):28-32.
[22]邬萌萌,谭晓风,周荣,等.油茶EMF2 基因的全长cDNA 克隆及序列分析[D].经济林研究,2013,31(2):7-12.
[23]Barna B,Györgyi B.Resistance of young versus old tobacco leaves to necrotrophs,fusaric acid,cell wall-degrading enzymes and autolysis of membrane lipids[J].Physiological and Molecular Plant Pathology,1992,40(4):247-257.
[24]Kus J V,Zaton K,Sarkar R,et al.Age-related resistance in Arabidopsis is a developmentally regulated defense response to Pseudomonas syringae[J].The Plant Cell Online,2002,14(2):479-490.
[25]Mutty S,Hossenkhan N T.Age-related resistance in commercial varieties of solanum tuberosum to the late blight pathogen,phytophthora infestans[J].Plant Pathology Journal,2008,7(2):168-173.
[26]Rusterucci C,Zhao Z,Haines K,et al.Age-related resistance to Pseudomonas syringae pv.tomato is associated with the transition to flowering in Arabidopsis and is effective against Peronospora parasitica[J].Physiological and Molecular Plant Pathology,2005,66(6):222-231.
[27]Zeier J.Age-dependent variations of local and systemic defence responses in Arabidopsis leaves towards an avirulent strain of Pseudomonas syringae[J].Physiological and Molecular Plant Pathology,2005,66(1):30-39.
