编码桔小实蝇抗阿维菌素的GABA 受体基因5'端序列片段克隆与分析
2014-12-09章玉苹陆永跃梁广文
章玉苹,陆永跃,梁广文,曾 玲
(1.华南农业大学昆虫生态研究室,广州 510642;2.广东省农业科学院植物保护研究所,广东省植物保护新技术重点实验室,广州 510640)
桔小实蝇Bactrocera dorsalis (Hendel)是世界许多国家和地区的重要危险性害虫,也是我国热带亚热带地区重要经济害虫之一,对水果等作物造成了巨大的经济损失。该虫原发于我国台湾和日本琉球群岛一带,后传到20 多个国家和地区,现已在我国许多地区发生(曾玲等,2006;梁广勤等,2002;刘玉章等,1981)。自2003年以来我国桔小实蝇田间种群的抗药性问题已突显出来,2004年台湾报道了桔小实蝇种群对十种农药(二溴磷 Naled、敌百虫 Trichlorfon、杀螟硫 磷Fenitrothion、倍硫磷 Fenthion、灭蚜硫Formothion、马拉硫 Malathion、灭多威Methomyl、氟氯氰菊酯 Cyfluthrin、氯氰菊酯Cypermethrin、氰戊菊酯Fenvalerate)抗性发展规律,发现桔小实蝇种群对二溴磷的抗性上升最慢,对马拉硫磷的抗性上升最迅速(潘志萍等,2005;Hsu et al.,2000;章玉苹等,2008)。我国大陆华南地区桔小实蝇发生为害猖獗,2004年已发现部分地区的桔小实蝇种群已对阿维菌素产生了低水平的抗性,2005年低水平抗性或敏感的田间种群抗性均上升,大部分达到了中抗水平,甚至接近高抗水平(曾玲等,2006;林进添等,2004;章玉苹等,2008)。
阿维菌素(Avermectin)是一种大环内酯抗生素类杀虫杀螨剂,它主要是通过干扰虫体内的γ-氨基丁酸(GABA)系统,使虫体处于麻痹状态而死亡,具有广谱高效、低毒、低残留、安全性高等特点(Lasota and Dybas,1991)。害虫对阿维菌素的抗性问题仍不容忽视。目前已发现家蝇Musca domestica L.、马铃薯甲虫Leptinotarsa decemlineata Say、二斑叶螨Tetranychus urticae Koch、德国蜚蠊Blattela germanica Linnaeus、小菜蛾 Plutella xylostella Linnaeus、三叶斑潜蝇 Liriomyza trifoli Burgess 等对阿维菌素产生了一定的抗性(梁广勤等,2002;潘志萍等,2005;章玉苹等,2008;Kaku and Matsumura,1994;Thompson et al.,1993)。其神经不敏感性抗性机制的分子机理主要有两方面:GABA 受体数目的变化和受体的质变。作为杀虫剂重要的靶标之一,GABA 受体的研究已成为杀虫剂毒理机制研究的热点之一。到目前为止,虽然关于阿维菌素对许多昆虫作用机制的研究较多,分子水平研究越来越迅速发展,但在桔小实蝇体内的作用机制的研究仍主要集中于生理生化水平。关于桔小实蝇GABA 受体基因克隆和序列分析国内外均未见报道。本文在前人对其它昆虫GABA受体研究的基础上,对编码桔小实蝇的GABA 受体基因进行研究,主要是通过克隆室内选育的对阿维菌素有高水平抗性的抗性品系及敏感品系中编码GABA 受体基因序列片段进行比对,初步从基因水平上探索桔小实蝇对阿维菌素产生抗性的分子机理。
1 材料与方法
1.1 供试昆虫
1.1.1 饲养方法
敏感品系(Susceptilble strain,简称SS):华南农业大学昆虫生态研究室提供。
抗性品系(Avermectin-selected strain of LC90,简称AvR90):以2003年3月采自广州市郊的野外桔小实蝇种群作为抗性品系筛选的原始材料,将采集的桔小实蝇在养虫室饲养1-2 代稳定后,幼虫采用微量点滴法处理,将药液点滴于幼虫胸部背面,处理后的幼虫放入湿润的沙中,待其化蛹、羽化正常的成虫继续饲养。成虫采用药膜法,测定亲代(F0)成虫对阿维菌素的毒力后,在此基础上采用LC90浓度进行群体筛选,每隔三代进行一次毒力测定,并分别用所测定的LC90浓度药剂处理后三代虫,使其种群形成不同抗药性品系,经过多代汰选培育形成。
1.1.2 毒力测定与计算方法
采用药膜法。将试药剂用丙酮配制成母液,稀释成5-6 个浓度。将5 mL 药液倒入250 mL 三角瓶中,摇动,当瓶壁上形成均匀药膜后,将药液倒掉。待丙酮完全挥发后,引入桔小实蝇成虫,放入沾有5%蜜糖水棉球后用纱布封住瓶口并用橡皮圈扎紧,倒立放置于室温25-28℃下,设丙酮为对照。每处理3 次重复,每重复10 头,72 h 后检查死亡率。以将其翻身后30 s 内不能翻转为死亡。对照组死亡率在10%以下为有效试验。
1.1.3 毒力计算方法
试验数据均用Excel 软件处理,计算出各药剂的毒力回归方程、致死中浓度(LC50)及其95%置信度、相关系数(r)等相关参数。抗性倍数(Rm)=供试种群的LC50/敏感种群的LC50,以LC50的95%置信限是否有重叠作为判断不同种群间药剂敏感度差异是否显著的标准,抗性水平划分标准:<3 倍为敏感;3-10 倍为低水平抗性;10.1-40 倍为中等抗性;40.1-160 倍为高水平抗性;>160 倍为极高水平抗性(唐振华和吴士雄,2000)。
1.2 供试药剂及仪器
90%阿维菌素原粉,深圳瑞德丰有限公司提供。微量点滴仪(0.04-0.05 μL),南京农业大学制。RNAiso Reagent、RNA PCR Kit (AMV)Ver3.0、PMD20-T 质粒、PMD18-T 质粒、Ex-Taq DNA 聚合酶、Agarose Gel DNA Purification Kit Ver.2.0 和DL2000 等均购自Takara 生物工程有限公司;IPTG、X-gal、Amp 购自上海生工生物工程技术服务有限公司。Thermocycler PCR 仪(Bio-Rad);PTC-100 型 PCR 仪 (MJ research);Centrifuge 5180R 冷冻高速离心机 (Effendorf);UVP 凝胶成像系统(Gene);SUB-Cell 电泳仪(Bio-Rad);超静工作台(Holten Lanim);紫外分光光度计(Beckman);DHP-9052 型电热恒温培养箱(上海一恒科技有限公司);HZQ-F160 全温振荡培养箱(哈尔滨市东联电子技术开发有限公司)。
1.3 总RNA 提取及cDNA 第一链合成
两品系总RNA 均用RNAiso Reagent 试剂进行总RNA 提取,所有操作按试剂盒说明书进行,利用RNA PCR Kit (AMV)Ver3.0 试剂盒反转录mRNA成cDNA 第一链,操作均按试剂盒说明书进行。
1.4 PCR 扩增反应
1.4.1 引物设计
根据已报道并在GeneBank 登录的地中海实蝇(登录号:AF172352)、果 蝇 (登录号:AY017266)及家蝇(登录号:AB177547)等编码GABA 受体基因的保守区域,以及同源序列氨基酸密码子的偏好性,设计1 对特异引物克隆编码桔小实蝇GABA 受体的5'端cDNA 片段。
上游引物:5'-ATGCCGCGTGCACGTCTCGTC-3'
下游引物:5'-CATAAATCTTTGTTTTCGCATTTG-3'
1.4.2 5'端cDNA 片段双链的合成
反应体系(25 μL):1 μL 的cDNA 第一链,5 μL的PCR Buffer (5×),0.125 μL 的Ex-Taq酶(5U),0.5 μL 的正反引物(25 mM),17.875 μL的ddH2O。反应条件:94℃3 min,94℃30 s,56℃30 s,72℃2 min,35 个循环,72℃延伸10 min,4℃保存。取5 μL 的DNA,用1.5%琼脂糖电泳鉴定PCR 产物,紫外线下观察电泳结果。
1.5 PCR 产物克隆、测定及同源性比较
利用Agarose Gel DNA Purification Kit Ver.2.0试剂盒纯化PCR 产物,纯化产物经连接反应连接到pMD20-Tvector 上,再将重组体克隆转化到感受态大肠杆菌中,通过蓝、白斑筛选将阳性克隆进行扩大培养直接委托上海生工生物工程技术服务有限公司测序,并将测得的cDNA 片段翻译成氨基酸序列,用BLAST 软件进行同源性比较和分析。
2 结果与分析
2.1 敏感与抗性品系的毒力测定结果
桔小实蝇对阿维菌素的抗性发展缓慢而稳定上升,筛选至27 代时,抗性倍数增长到113.12;抗性上升最快的阶段是第21 代至第24 代时,抗性倍数从51.76 上升至93.77。从各代次的毒力回归线的斜率来看,该品系毒力回归线从平变陡后进入较平稳状态,表明该种群已处于抗性纯合子数量占绝大多数的阶段,群体异质性越来越小(表1)。

表1 阿维菌素抗性品系(AvR90)抗药性测定结果Table 1 The resistance results of AvR90strain to avermectin
2.2 编码GABA 受体基因5'端序列克隆结果分析
PCR 产物经琼脂糖电泳检测,获得与预期的片段大小相符合的目的带(如图1)。经纯化(图2)、克隆后测序,获得20 条长度为1053 bp 的5'端cDNA 片段。
2.3 序列分析
通过20 个单菌落测序结果,扩增得编码GABA 受体的5'端cDNA 片段1053 bp (表2),编码351 个氨基酸(表3)。敏感品系SS 与抗性品系AvR90的该cDNA 碱基序列片段有一个碱基差异,即A496T,但根据该两序列推导的氨基酸序列完全相同,因此该碱基突变为同义突变。
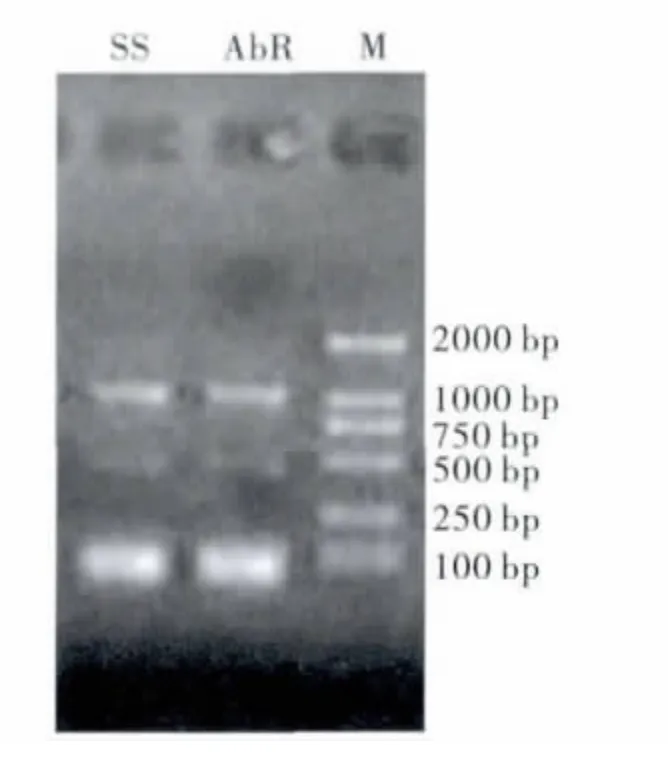
图1 编码桔小实蝇GABA 受体基因5'cDNA 片段PCR 产物从左至右:AvR90品系;SS 品系;M 为DL2000Fig.1 PCR result of cDNA fragment of encoding GABA receptor gene (Left to right:AvR90strain,SS strain,DL2000)
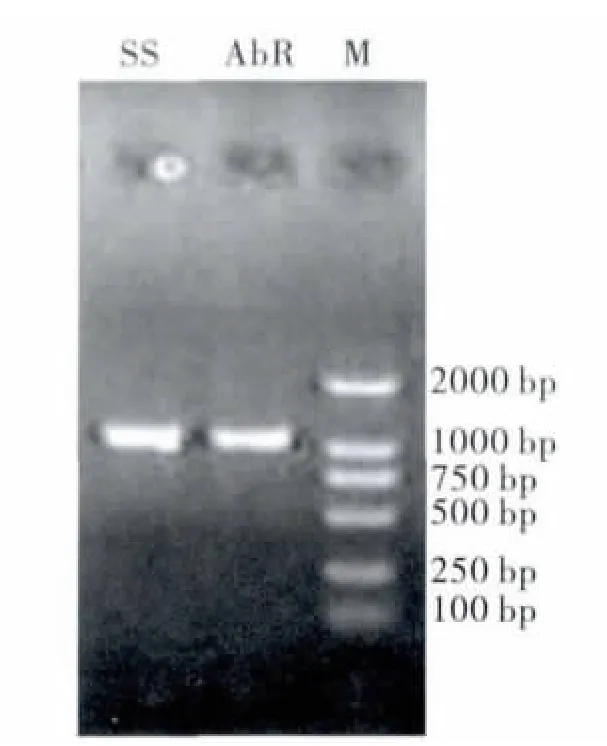
图2 编码桔小实蝇GABA 受体基因5' cDNA 片段纯化从左至右:AvR90品系;SS 品系;M 为DL2000Fig.2 Purified result of cDNA fragment of encoding GABA receptor gene (Left to right:AvR90strain,SS strain,DL2000)
利用NCBI 的Blast 程序进行同源分析,结果显示克隆获得的桔小实蝇敏感品系SS 及抗性品系AvR90的编码GABA 受体基因的5'端cDNA 片段编码的氨基酸片段与Genebank 上与其他昆虫的同源序列具有很高的同源性,其推导的氨基酸序列与地中海实蝇Ceratitis capitata、家蝇、黑腹果蝇Drosophlia melanogaster Meigen、绿 蝇 Lucilia cuprinaedit、意大利蜜蜂Apis mellifera、稻飞虱Amazona aestiva、小菜蛾等分别有99%、99%、99%、98%、97%、97%、96%同源性。

表2 编码桔小实蝇GABA 受体基因的序列片段(1053 bp)Table 2 Partial sequence of encoding GABA receptor gene of B.dorsalis
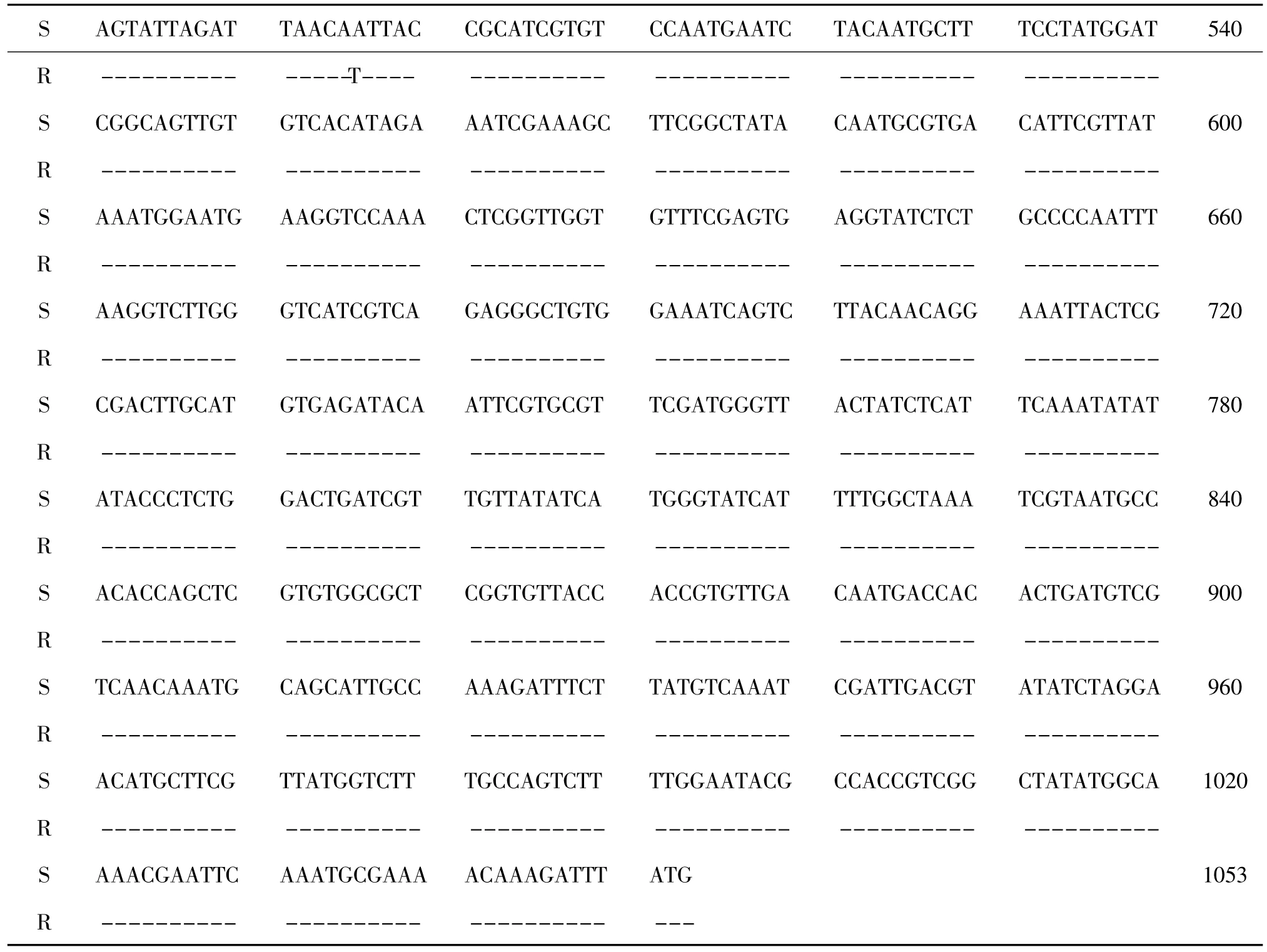
(续上表)

表3 编码桔小实蝇GABA 受体基因片段推导的氨基酸序列片段Table 3 Partial sequence of amino acid of encoding GABA receptor gene of B.dorsalis
3 结论与讨论
阿维菌素是以GABA 受体为主要靶标的一类生物源农药,主要作用于GABA 门控的氯离子通道(潘志萍等,2008),阿维菌素可刺激突触前膜大量释放GABA 促使GABA 门控的氯离子通道持续开放,大量氯离子涌入膜内,造成神经膜电位超极化,形成抑制性突触电位,影响害虫正常神经活动,最终导致死亡。其神经不敏感性抗性机制的分子机理主要有两方面:GABA 受体数目的变化和受体的质变。Matsumura 等报道其使蜚蠊抗性品系受体密度显著减小(Matsumura et al.,1987)。Ffrench-constant 等则发现GABA 受体的质变,其cDNA 处有两处发生突变,第995 位的G 突变为T,1174 位的G 突变为A,结果导致氨基酸序列第302 位丙氨酸变为丝氨酸,第361 位的甲硫氨酸变为异亮氨酸,此外大量研究表明许多害虫抗阿维菌素品系均出现了第302 位丙氨酸突变为丝氨酸,这说明害虫的GABA 受体基因具有高度保守性(Ffrench-Constant et al.,1991;Ffrench-Constant et al.,1993)。
而本文首次获得编码桔小实蝇GABA 受体基因的5'端片段序列1053 bp,经序列分析表明抗性品系和敏感品系推导的氨基酸序列完全相同,两品系编码GABA 受体基因的该段氨基酸序列没有存在位点突变,通过比较相似序列而知本文所得的序列的第305 位氨基酸即为编码GABA 受体基因的保守突变位点,但两品系该位氨基酸仍为丙氨酸,未发生突变。因此可推断抗阿维菌素品系在此抗性倍数下其抗性产生的原因并不是该保守位点发生突变的结果,导致抗性的原因可能是其他因素,如代谢酶活性的变化、GABA 受体数目的变化或者是编码GABA 受体序列3'端片段其他位点突变造成,如第361 位的甲硫氨酸突变为异亮氨酸,具体情况还需要进一步研究。
References)
Ffrench-Constant RH,Rocheleau TA,et al.A point mutation in a Drosophila GABA receptor confers insecticides resistance [J].Nature,1993,363 (3):449-451.
Ffrench-Constant RH,Mortlock DP,Shaffer CD,et al.Molecular cloning and transformation of cyclodiene resistance in Drosophila:An invertebrated GABA receptor locus [J].PNAS,1991,88:7209-7213.
Hsu JC,Feng HT,Wu WJ.Resistance and synergistic effects of insecticides in Bactrocera dorsalis (Diptera:Tephritidae) in Taiwan [J].Journal of economic entomology,2004,97 (5):1682-1688.
Hsu JC,Feng HT.Insecticide susceptibility of the oriental fruit fly(Bactrocera dorsalis (Hendel))(Diptera:Tephritidae)in Taiwan[J].Chinese J.Entomol.,2000,20:109-118.
Kaku K,Matsumura F.Identification of the site of mutation within the M2 region of the GABA receptor of the cyclodience resistant german cockroach[J].Comparative Biochemistry and Physiology Part C:Pharmacology,Toxicology and Endocrinology,1994,108 (3):367-376.
Lasota JA,Dybas RA.Avermectins,a novel class of compounds:implications for use in arthropod pest control [J].Annual review of Entomology,1991,36:91-117.
Liang GQ,Liang F,Wu JJ,et al.The study on strategies of Bactrocera dorsalis control [J].Guangdong Agriculturl Science,2002,(2):37-40.[梁广勤,梁帆,吴佳教,等.实蝇防除策略和措施的研究[J].广东农业科学,2002,(2):37-40]
Liu YZ.Study on the oriental fruit fly [J].Journal of Entomological Society,1981,16 (1):9-26.[刘玉章.台湾东方果实蝇之研究[J].兴大昆虫学会会报,1981,16 (1):9-26]
Lin JT,Zeng L,Lu YY,et al.Advances on biological characteristics and control of Bactrocera dorsalis [J].Journal of Zhongkai Agrotechnical College,2004,17 (1):60-67.[林进添,曾玲,陆永跃,等.桔小实蝇生物学特性及防治研究进展[J].仲恺农业技术学院学报,2004,17 (1):60-67]
Matsumura F,Tanaka K,Ozoe Y.GABA-related systems as targets for insecticides in“Sites and Action of Neurotoxic Pesticides,ACS Symposium Series 356”[C].Holling-worth RM,Green MB,eds.American Chemical Society,Washington D.C,1987,44-70.
Pan ZP,Zeng L,Lu YY.Monitoring of resistance of oriental fruit fly adults to insecticides in South China [J].Journal of South China Agricultural University,2005,26 (4):23-26.[潘志萍,曾玲,陆永跃.华南地区桔小实蝇对几种农药的抗药性研究[J].华南农业大学学报,2005,26 (4):23-26]
Pan ZP,Lu YY,Zeng L,et al.Development of resistance totrichlorophon,alphamethrin,and avermectin in laboratory populations of the oriental fruit fly,Bactrocera dorsalis (Hendel)(Diptera:Tephritidae)[J].Acta Entomologica Sinica,2008,51(6):609-617.[潘志萍,陆永跃,曾玲,等.桔小实蝇实验种群对敌百虫、高效氯氰菊酯和阿维菌素的抗性增长规律[J].昆虫学报,2008,51 (6):609-617]
Pan ZP,Zeng L,Lu YY.Monitoring of resistance of oriental fruit fly adults to insecticides in South China [J].Journal of South China Agricultural University,2005,26 (4):23-26.[潘志萍,曾玲,陆永跃.华南地区桔小实蝇对几种农药的抗药性研究[J].华南农业大学学报(自然科学版),2005,26 (4):23-26]
Tang ZH,Wu SX.Heredity and Evolution of Insect Resistance to Pesticides [M].Shanghai:Shanghai Scientific and Technical Literature Publishing House,2000.[唐振华,吴士雄,2000.昆虫抗药性的遗传与进化[M].上海科学技术文献出版社.2000]
Zhang YP,Zeng L,Lu YY,et al.Monitoring of Insecticide Resistance of Bactrocera dorsalis (Hendel)in South China [J].Journal of Huazhong Agricultural University (Natural Science Edition),2008,27 (4):456-459.[章玉苹,曾玲,陆永跃,等.华南地区桔小实蝇田间种群抗药性的监测[J].华中农业大学学报,2008,27 (4):456-459]
Zhang YP,Zeng L,Lu YY,et al.Monitoring of insecticide resistance of Bactrocera dorsalis adults in South China [J].Journal of SouthChina Agricultural University,2007,28 (3):20-23.[章玉苹,曾玲,陆永跃,等.华南地区桔小实蝇抗药性动态监测[J].华南农业大学学报,2007,28 (3):20-23]
Zeng L,Lu YY,Lin JT,et al.Strategies and technical solution of Bactrocera dorsalis control [C].The Plant Protection Bulletin,Kunming,Yunnan,2006,11:19-23.[曾玲,陆永跃,林进添,等.桔小实蝇的防治策略和技术.成卓敏主编:中国植物保护学会2006年学术年会“科技创新与绿色植保”[C].北京:中国农业科学技术出版社.云南昆明,2006,11:19-23]
