不同辣椒品种光合日变化及影响因子分析
2014-12-07蓬桂华张爱民邢丹苏丹韩世玉
蓬桂华,张爱民,邢丹,苏丹,韩世玉
(贵州省辣椒研究所,贵阳,550006)
光合作用被称为“地球上最重要的化学反应”和“生命界最重大的顶极创造之一”[1],植物干物质生产的95%来自于光合作用[2]。因此,研究作物光合作用的遗传,有助于选育出强光合作用的品种,达到提高农作物产量的目的[3]。
多年来,国内外学者对辣椒光合作用的研究非常重视[4],邹学校等[3]指出,光合作用的强弱不仅受植株叶位、生育期、环境因子、栽培条件的影响,而且不同品种的净光合速率差异很大。颉建明等[2]研究了陇椒系列辣椒的光合特性指出,陇椒系列辣椒具有较高的叶绿素含量和净光合速率,利用弱光的能力较强。徐小蓉等[5]研究两种长势一致的不同辣椒品种(Onza和Cajamarca)的光合特性,表明两个长势相同的辣椒品种的光合速率存在差异。贵州辣椒品种资源丰富、特色突出,栽培历史悠久[6],全省已建立8个不同生态区,10个区域试验点,已审(认)定贵州地方特色辣椒品种42个,杂交品种10个,并形成了黔辣、黔椒、贵椒、遵辣4个系列新品种[7]。然而,关于贵州地方辣椒资源光合特性评价方面的研究还未见报道。所以,本研究以4个贵州地方辣椒品种为材料,探讨不同品种之间的光合日变化及其影响因子,为地方辣椒品种高产及种质资源的筛选、育种和栽培提供理论基础。
1 材料与方法
1.1 试验材料
辣椒种子由贵州省辣椒研究所提供,有罗甸白辣椒(果实向上、乳白色、指形),黄平1号(果实向下、绿色、细线形),独山2号(果实向下、绿色、果皮微皱、长线形)和独山3号(果实向下、绿色、果皮微皱、长线形)。
1.2 试验方法
①辣椒育苗 采用漂浮育苗技术,每穴播2粒种子,待幼苗长出真叶时施入少量复合肥,当幼苗4叶1心时定植于田间。
②整地与施肥 每667 m2施入复合肥(黄金营养元素)20 kg、过磷酸钙50 kg作底肥,开厢种植,厢面宽80 cm,厢距40 cm,厢面上双排种植,株距40 cm。
③光合日变化的测定 待辣椒生长到初花期时,选择晴朗的一天,从 7:00~19:00,以 2 h 为一个测量间隔,利用美国 LI-COM公司生产的 Li-6400XT便携式光合作用测定仪,采用开放式气路,测定辣椒的光合日变化,测定的参数包括净光合速率(Pn,μmol CO2·m-2·s-1)、蒸腾速率(Tr,mmol H2O·m-2·s-1)、胞间 CO2浓度(Ci,μmol CO2·mol-1)和气孔导度 (Gs,mol H2O·m-2·s-1)、 光合有效辐射(PAR,μmol·m-2·s-1)、气温(Ta,℃)、叶温(Tl,℃)、大气 CO2浓度(Ca,μmol CO2·mol-1)和空气湿度(RH,%)。测定时,选择生长健壮、着生方位一致的成熟叶片,每个品种辣椒测量5株,取平均值。

表1 环境因子平均日变化
1.3 数据分析
采用Microsoft Office 2003和DPS进行数据整理与分析。
2 结果与分析
2.1 环境因子的平均日变化
光合有效辐射(PAR)、气温(Ta)、叶温(Tl)、大气CO2浓度(Ca)和空气湿度(RH)的平均日变化详见表1。从表1中可以看出,PAR日变化为先升后降,7:00最低,之后逐渐升高,到 13:00时达到最大值,之后又开始下降。Ta、Tl与PAR的变化相似,也是先升后降,在 7:00~19:00 的测定时间内,最高气温与最低气温相差10.36℃,最高叶温与最低叶温相差 11.55℃, 在 7:00、11:00、17:00 和 19:00,Ta>Tl,而在 9:00、13:00 和 15:00,Tl>Ta,这可能是测量时叶室封闭的环境导致。Ca的变化不明显,以7:00的Ca 最高,达到 411.70 μmol CO2·mol-1,之后开始下降,13:00后又逐步升高,这可能与植物光合作用下降有关。RH的变化不明显,以7:00时最高,之后一直下降,最高空气湿度与最低空气湿度相差14.83%。
2.2 辣椒净光合速率(Pn)日变化
净光合速率又称表观光合作用,是指真正光合作用所同化的CO2量减去因呼吸作用而释放的CO2量,其大小除受自身生物学特点(气孔构造、叶面内部的面积大小等)的制约外,还受光合有效辐射、大气温度、CO2浓度、空气相对湿度等多种生态因子的影响[8]。
从图1可以看出,4个辣椒品种的净光合速率日变化均成双峰曲线,但各品种峰值变化不完全相同。独山3号和独山2号的变化较为相似,峰值分别出现在 11:00 和 15:00, 不同的是 7:00~9:00独山3号的Pn上升速度慢于独山2号,而在9:00~11:00,独山 3 号比独山 2 号上升得快;罗甸白辣椒与黄平1号的第一个峰值均出现在9:00,之后 Pn开始下降,但黄平 1号在 13:00时出现另一个峰值,而罗甸白辣椒继续下降,到15:00才出现第二个峰值;到17:00以后,各品种Pn均快速下降。
2.3 辣椒胞间CO2浓度(Ci)日变化
从图1与图2中可以看出,4个辣椒品种的胞间CO2浓度日变化曲线与净光合速率变化曲线相反,呈倒双峰曲线,但各品种的峰值点出现不一致。
罗甸白辣椒、黄平1号和独山2号在9:00时出现第一个低谷,而独山3号在11:00才出现第一个低谷。罗甸白辣椒与独山3号在15:00时出现第二个低谷,独山2号则在17:00时出现第二个低谷,而黄平1号则在13:00时出现第二个低谷。
2.4 辣椒气孔导度(Gs)日变化
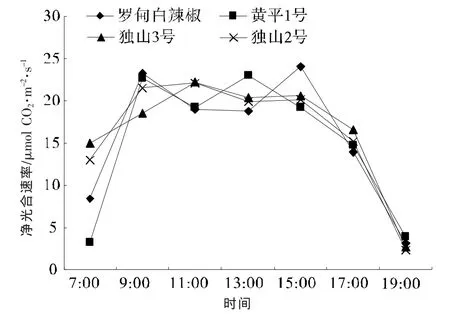
图1 辣椒净光合速率日变化

图2 辣椒胞间CO2浓度日变化
从图3中可以看出,罗甸白辣椒与独山2号的气孔导度日变化呈双峰曲线,峰值分别出现在11:00和15:00,而黄平1号和独山3号的气孔导度日变化呈单峰曲线变化,但峰值出现的时间不同,黄平1号出现在 13:00,独山 3 号出现在 15:00。
11:00以前,随着气温的升高、光线的增强,各品种的气孔导度均快速升高,15:00以后,随着气温的降低和光线的减弱,气孔导度又开始快速下降。中午时,由于温度最高,光线最强,为了保护自身器官免受伤害,植物会关闭气孔或减小气孔的开度。
2.5 辣椒蒸腾速率(Tr)日变化
从图4中可以看出,品种不同,其蒸腾速率日变化不完全相同,但差异主要体现在11:00~15:00之间,其他时间变化较为一致。
从 7:00~11:00,各品种的蒸腾速率快速升高,而 15:00 之后开始快速下降, 以 17:00~19:00 下降最快。罗甸白辣椒在在11:00后开始下降,13:00后又开始升高,呈现双峰变化;黄平1号在11:00后继续升高,到13:00后开始下降,呈现单峰曲线变化;独山3号在11:00后继续升高,一直到15:00之后才开始下降,也呈单峰曲线变化;独山2号11:00~15:00 变化不明显。
2.6 辣椒净光合速率与其他参数的相关性
作物光合作用日变化受诸多因素影响,如植物的种类、环境条件的变化等等。以4种辣椒的净光合速率( Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)、蒸腾速率(Tr)、空气温度(Ta)、叶片温度(Tl)、空气 CO2浓度(Ca)、空气相对湿度(RH)和光合有效辐射(PAR)作为相关变量进行多元相关分析,相关分析结果详见表2。从表2中可以看出,罗甸白辣椒的Pn与 Gs、Tr、Tl和 PAR 呈极显著正相关(p<0.01),与 Ta呈显著正相关(p<0.05),与Ci和 Ca呈极显著负相关(p<0.01),与 RH 的相关性不显著(p>0.05)。黄平 1号的 Pn 与 Gs、Tr、Tl、Ta 和 PAR 呈 极显著 正相关(p<0.01),与 RH 呈显著正相关(p<0.05),与 Ci和 Ca呈极显著负相关(p<0.01)。独山3号与独山2号的表现较为一致,其 Pn 与 Gs、Tr、Ta、Tl和 PAR 呈极显著正相关(p<0.01),与 Ci和 Ca呈极显著负相关 (p<0.01),与RH 的相关性不显著(p>0.05)。

图3 辣椒气孔导度日变化

图4 辣椒蒸腾速率日变化
另外,不论是哪一个品种,其Pn与Ca、Ci都呈极显著负相关,但我们不能因此就说Ca或Ci越低,其Pn就高。结合表1可以看出,早晨空气中的CO2浓度较高,而光线较弱,此时的Pn较低,随着光线的增强,植物光合作用也升高,从而降低了环境中的CO2浓度;而Ci值的大小由叶片周围的CO2浓度、气孔导度及叶肉细胞光合活性3个因素决定,当叶肉细胞的光合活性随光强增高而逐步增大时,必将引发Ci进一步降低。所以,Ca与Ci的变化只是植株光合作用变化的一种结果,而不是其原因。
3 结论与讨论
光合作用是绿色植物特有的生理功能,是生态生理因子共同作用的过程,参与光合作用的生态生理因子大多表现出明显的日变化,因此光合作用也呈明显的日变化,研究其变化特征,对分析作物光合生产力和产量形成有一定的理论和实践意义[9]。徐小蓉等[5]对Onza和Cajamarca的研究指出,两组辣椒的净光合速率在11:30出现最大值,之后一直下降;曹振木等[10]对不同耐热辣椒叶片的光合特性研究指出,4个辣椒品系的净光合速率与蒸腾速率日变化均呈典型的双峰曲线,即存在光合“午休”现象,第一个峰值出现在 9:00~10:00,第二个峰值出现在15:00以后,而气孔导度日变化不完全相同。

表2 不同辣椒品种净光合速率及其影响因子间相关性系数
本文研究指出,4个辣椒品种净光合速率均呈双峰曲线变化,胞间CO2浓度呈“倒双峰”曲线变化,而气孔导度与蒸腾速率的日变化因品种而异,有单峰,也有双峰,但峰值的大小及峰值出现时间因品种不同而表现出一定差异。在所考察的8个影响因子中,气孔导度、蒸腾速率、叶片温度和光合有效辐射与净光合速率呈极显著正相关,胞间CO2浓度和空气CO2浓度与净光合速率极显著负相关。
光合作用是绿色植物物质生产的基础,植物叶片光合性能与其生产能力呈正相关[11],但植物光合作用的大小受诸多因素的影响,如品种、叶绿素含量及比例、气孔大小及数量、光呼吸、暗呼吸、光照强度、光照角度、空气CO2浓度、空气温度等等。本文仅仅简述了4个辣椒品种光合日变化及其影响因子,有关品种之间表现出来的差异及环境因子如何影响辣椒光合进程还有待今后进一步深入分析。
[1]程建峰,沈允钢.试析光合作用的研究动向[J].植物学报,2011,46(6):694-704.
[2]颉建明,郁继华,颉敏华,等.陇椒系列辣椒光合特性研究[J].甘肃农业大学学报,2008,43(1):105-109.
[3]邹学校,马艳青,刘荣云.辣椒净光合速率配合力分析[J].中国农业科学,2006,39(11):2 300-2 306.
[4]陈银华,蒋健箴.光照强度对辣椒光合特性与生长发育的影响[J].上海农业学报,1998,14(3):46-50.
[5]徐小蓉,罗在柒,张习敏,等.辣椒(Onza 和 Cajamarca)光合特性研究[J].贵州科学,2011,29(5):80-84.
[6]王永平,张绍刚,张婧.做大做强贵州辣椒产业的对策思[J].贵州农业科学,2009,37(7):129-132.
[7]张绍刚.贵州辣椒产业发展与品种需求分析[J].中国蔬菜,2011(21):18-19.
[8]丁小球,胡玉佳,王榕楷.三种草坪草净光合速率和蒸腾速率的日变化特点研究[J].草业科学,2001,18(2):62-66.
[9]唐微,邓仲篪,朱名安.籼稻、粳稻和杂交稻的光合速率日变化及其与生态、生理因子的关系[J].湖北农业科学,2001(4):14-16.
[10]曹振木,詹园凤,刘维侠.营养生长期不同耐热辣椒叶片光合特性比较研究[J].热带农业科学,2006,22(11):436-439.
[11]丛伟,李霞,盛婧,等.南京夏季不同叶位凤眼莲叶片光合作用的日变化及其生态因子分析[J].江西农业大学学报,2011,33(3):445-451.
