重组分泌型人类免疫缺陷病毒Tat蛋白在真核细胞中的表达及活性检测*
2014-11-21韦红玉曾怡唐华英赵丽娟右江民族医学院微生物学与免疫学教研室广西百色533000
韦红玉,曾怡,唐华英,赵丽娟(右江民族医学院微生物学与免疫学教研室,广西百色 533000)
人类免疫缺陷病毒1型(HIV-1)感染细胞可释放大量的可溶性蛋白,其中包括Tat蛋白。Tat蛋白是HIV-1病毒复制所必需的反式激活因子,但Tat蛋白没有分泌信号,只是当T细胞急性感染HIV-1时,Tat蛋白可被释放到细胞外间质,并可作用于其他细胞或细胞内感染的其他病毒,发挥细胞转化或激活病毒复制周期的作用。已有研究证实Tat蛋白与HIV-1 RNA 5′端启动子上的反式激活反应元件(TAR)结合,激活HIV-1长末端重复序列(LTR)使其下游的基因得以表达,从而促进整个HIV-1转录本的表达[1]。研究表明Tat蛋白可激活人类疱疹病毒6型、8型,也可促进血管内皮细胞和艾滋病(AIDS)相关的卡波济肉瘤(KS)细胞的生长、浸润、分化及黏附等。
为研究细胞外Tat蛋白对其他病毒复制周期的影响,本研究在构建了重组HIV-1tat基因的分泌型真核表达载体的基础上,对Tat蛋白的表达及生物活性进行了检测,为进一步研究Tat蛋白的生物学作用,探索细胞外Tat蛋白对正常细胞或其他细胞内潜伏感染病毒的影响奠定基础。
1 材料与方法
1.1 材料
1.1.1 质粒、细胞 pcDNA3.1(+)/Tat质粒在tat基因末端加入Linker序列GGAGGC,并融合Flag序列GACTACAAGGACGACGATGACAAG,以便利用抗Flag标签抗体检测Tat蛋白的表达。BCBL-1细胞为人类疱疹病毒8型感染、EB病毒阴性的体腔渗出性淋巴瘤细胞系,用含10%小牛血清的1640培养基,于5%CO2、37℃培养。人胚肾上皮细胞293细胞用含10%小牛血清的DMEM 培养基,于5%CO2、37℃培养。pcDNA3.1(+)/Tat质粒、LTR-CAT 质粒、BCBL-1细胞由南京医科大学卢春教授馈赠。293细胞为本实验室保存。分泌型真核表达载体pSecTag2B 购自美国Invitrogen 公司。
1.1.2 试剂与仪器参考文献[2]设计聚合酶链反应(PCR)引物,并在PCR 上游引物的5′端引入HindⅢ酶切位点,下游引物的5′端引入BamHⅠ酶切位点,为增强其表达,在上游引物5′端起始密码子ATG 前添加了KOZAK 序列(GCCACC)以增强基因的表达,同时为便于Tat蛋白的检测,原质粒中tat基因下游融合的Flag序列一并扩增。上游引物序列(方框内为KOZAK 序列,划线部分为HindⅢ酶切位点)P15′-taa tgccat gag ttc cac ac-3′,下游引物序列(划线部分为BamHⅠ酶切位点)gca g-3′;预期片段大小为807bp。同时设计另一对引物,上游引物5′端引入BamHⅠ酶切位点,P35′-taacac cat gag ttc cac ac-3′,下游引物5′端引入HindⅢ酶切位点,P45′-gga caa gct tga tga aca gta gca gca g-3′,此对引物扩增获得的反向序列同时构建重组分泌型载体,作为重组对照质粒。PCR 引物均由上海英骏生物工程公司合成。Lammda DNA EcoRⅠ+HindⅢ分子量标记、100bp ladder plus DNA 分子量标记、PCR 所用试剂Taq DNA 多聚酶及缓冲液、dNTP、限制性内切酶、T4DNA 连接酶、小牛碱性磷酸酶购自Fermentas公司,感受态DH5α购自北京天根生化科技公司,质粒提取试剂盒、DNA 胶回收试剂盒购自美国Omega 公司。Western blot所用试剂anti-Flag、anti-βactin、HRP-羊抗兔抗体、ECL发光试剂购自碧云天生物技术研究所。其他试剂均购自Amresco。研究中主要使用下列仪器:1-15PK 冷冻离心机(德国Sigma)、Mycycler PCR 仪(美国Bio-Rad)、垂直电泳转印系统(美国Bio-Rad)、Chemidoc XRS型凝胶成像系统(美国Bio-Rad)。
1.2 方法
1.2.1 HIV-1tat基因的扩增及纯化取pcDNA3.1(+)/Tat质粒做模板,用PCR 引物扩增HIV-1tat基因,PCR 反应体系包括:上游引物10pmol;下游引物10pmol;10×Taq DNA 聚合酶缓冲液5μL;MgCl 2.5mmol/L;dNTPs每种2.5 mmol/L;Taq DNA 聚合酶2单位;总反应体积50μL。扩增条件:预变性94℃5 min;按94℃1 min,59℃30s,72℃1min,30个循环;72℃延伸5min。取5μL PCR 反应产物用10g/L琼脂糖凝胶电泳后,在紫外灯下观察确认扩增成功。平衡酚:氯仿法纯化剩余的PCR 反应产物。
1.2.2 tat基因重组分泌型真核表达载体的构建及tat基因插入方向的鉴定分别取1μg PCR 产物和1μg pSecTag2B,经HindⅢ酶切,10g/L琼脂糖凝胶电泳后切胶回收DNA 片段。T4DNA 连接酶16℃连接过夜后,转化感受态DH5α,涂布含100μg/mL氨苄西林(Ampicillin,Amp)的LB平板,37℃培养16~18h。挑取5个生长的细菌克隆至含100μg/mL Amp的LB液体培养基,37℃250r/min 振荡培养16~18h。提取质粒,HindⅢ与BamHⅠ酶切初步鉴定。经HindⅢ与BamHⅠ酶切初步鉴定为阳性的质粒,送上海生工公司进行序列测定。确定tat基因正向插入克隆(命名为pSecTat)和tat基因反向插入克隆(命名为pSecTat-AS)。
1.2.3 Western blot检测tat基因重组质粒在293细胞中的表达及其分泌表达采用Invitrogen 公司生产的Lipofectamine 2000转染试剂,按使用说明书进行转染试验。将pSecTat转染至293细胞中,培养于六孔板,于转染后24、48h收集293细胞,Western blot检测真核细胞内Tat蛋白表达;用1×SDS凝胶上样缓冲液裂解细胞,煮沸变性后进行聚丙烯酰胺凝胶电泳(SDS-PAGE),并转移PVDF 膜。分别以anti-Flag、anti-βactin 为一抗、HRP-羊抗兔抗体为二抗,Western blot检测Tat蛋白细胞内表达情况,用增强的化学发光试剂ECL检测,最后用柯达X 线片感光。
1.2.4 CAT-酶联免疫吸附试验(ELISA)检测细胞内表达的重组Tat蛋白的调控活性将pSecTat与LTR-CAT 质粒(质粒中氯霉素乙酰转移酶CAT 由HIV LTR 启动,LTR 激活则CAT 表达)共转染293细胞和BCBL-1细胞,转染后24h,收集细胞,按CAT-ELISA 试剂盒说明书检测CAT 表达水平;从而检测细胞内表达的Tat蛋白的调控活性,同时以Tat基因反向重组质粒作为阴性对照。
1.2.5 CAT-ELISA 检测表达分泌至细胞外表达的重组Tat蛋白的调控活性将pSecTat转染至293细胞,培养于Transwell培养系统上层,将LTR-CAT 质粒转染BCBL-1细胞,培养于Transwell培养系统下层,采用Transwell培养系统检测pSecTat转染至293细胞后分泌Tat蛋白及其胞外调控活性,分别于转染后6、24、48、72h收集BCBL-1细胞;另采用同样方法检测pSecTat在肾系膜细胞中的分泌表达及其对BCBL-1细胞内LTR-CAT 的调控活性。收集的细胞按CAT-ELISA试剂盒说明书检测CAT 表达水平;同时以Tat基因反向重组质粒作为阴性对照。
2 结果
2.1 HIV-1tat基因的扩增用引入限制性酶切位点的PCR引物,经PCR 扩增并用10g/L 琼脂糖凝胶电泳后,在300bp附近出现条带,与预期的324bp大小相符。见图1。
2.2 tat重组分泌型真核表达载体的鉴定挑取正向和反向阳性克隆各2个提取的质粒用限制性内切酶HindⅢ与BanHⅠ酶切,并用10g/L琼脂糖凝胶电泳后,共有2个正向克隆和1个反向克隆在约330bp和约5200bp位置出现条带,其中1个反向克隆未见目的条带,见图2。将2个正向克隆质粒和1个反向克隆质粒送上海生工公司测序,结果显示克隆的tat基因序列与GenBank中登记的HIV-1tat基因100%同源。

图1 PCR 扩增HIV-1tat基因

图2 tat重组分泌型真核表达载体的酶切鉴定
2.3 Western blot检测tat基因重组质粒在293细胞中的表达 Western blot结果显示,pSecTat在能在293细胞中表达,用anti-Flag抗体可检测到约20×103的条带,与预期的Tat蛋白的19×103分子量大小一致,而反向插入tat基因的对照质粒pSecTat-antise转染293细胞则检测不到该条带(图3)。

图3 重组Tat蛋白的Western blot检测
2.4 CAT-ELISA 检测重组Tat蛋白在293细胞中的活性pSecTat与LTR-CAT 质粒共转染293细胞后24、48h,CATELISA 检测293细胞内CAT 的表达水平(图4)。结果显示,与pSecTat-AS对照质粒相比,pSecTat可刺激CAT 表达(P<0.05),并随转染时间增加,CAT 表达增加。
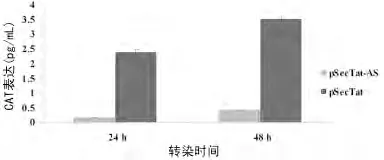
图4 CAT-ELISA 检测重组Tat蛋白在293细胞中的活性
2.5 重组Tat蛋白在293细胞、BCBL-1细胞中的活性比较pSecTat与LTR-CAT 质粒共转染293细胞、BCBL-1细胞后48h,CAT-ELISA 检测293细胞、BCBL-1细胞内CAT 的表达水平(图5)。结果显示,与pSecTat-AS对照质粒相比,pSecTat在293细胞及BCBL-1细胞中均可表达,但在293细胞中的表达效率高于在BCBL-1细胞中的表达(P<0.05)。

图5 重组Tat蛋白在293细胞、BCBL-1细胞中的活性比较
2.6 重组分泌型Tat蛋白的细胞外活性检测采用Transwell培养系统共培养转染pSecTat的293细胞与转染LTRCAT 的BCBL-1细胞,通过检测BCBL-1细胞中CAT 表达水平,判断pSecTat在293细胞中表达后分泌至细胞外的Tat蛋白的活性能否刺激BCBL-1细胞内CAT 表达。结果显示,与pSecTat-AS对照相比,pSecTat转染的293细胞可使BCBL-1细胞中CAT 蛋白表达显著。但转染时间延长至72h时,分泌型Tat已无活性(图6)。
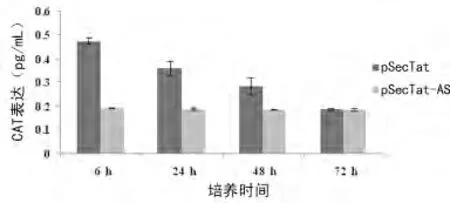
图6 CAT-ELISA 检测重组分泌型Tat蛋白的细胞外调控活性
3 讨论
HIV-1反式转录激活因子(Tat)是HIV-1长末端重复序列(LTR)转录起始和病毒增殖所必需的调节蛋白,含86~101个氨基酸[3-4]。除了在病毒基因表达中的转录激活作用之外,Tat蛋白还可从HIV-1感染的完整细胞内释放至细胞外,并保持活性。在HIV-1感染者血清中或体外培养细胞上清液中的Tat蛋白浓度可达40ng/mL[5-6]。这种外源的Tat蛋白可进入HIV-1未感染细胞诱导细胞凋亡,或进入潜伏感染细胞激活病毒基因组转录。Tat蛋白在HIV-1的致病机制中不仅是HIV-1复制所必需的蛋白,还发挥了细胞外毒素的作用。
本研究采用PCR 从pcDNA3.1(+)/Tat质粒中扩增融合Flag标签的tat全基因,利用HindⅢ与BamHⅠ双酶切后,将其正向克隆入分泌型真核表达载体pSecTag2B 中,同时构建反向克隆作为对照。进行基因测序确定基因的序列和克隆方向后,进一步利用抗Flag抗体,通过Western blot检测到Tat-Flag融合蛋白在293细胞内的表达。本研究还通过pSecTat与LTR-CAT 共转染293细胞、pSecTat与LTR-CAT 分别转染Transwell培养系统中上下层的细胞,进一步确认表达的Tat-Flag融合蛋白不但在293细胞内具有转录激活活性,还可分泌至细胞外,并进入BCBL-1细胞内,激活细胞内转的LTR-CAT 报告质粒表达CAT。结果表明,本研究成功构建了tat基因重组分泌型真核表达载体,该载体不仅可在细胞内表达有转录激活活性的Tat-Flag蛋白,该融合蛋白还可分泌至细胞外,通过旁分泌作用,发挥其生物学活性。
在美国,有超过20%的AIDS患者同时患有KS,这类KS称为AIDS-KS。多项研究表明,卡波济肉瘤相关疱疹病毒(KSHV)的感染与KS的发生密切相关,而HIV-1感染者更容易发生KS。本课题组先前的研究证实,细胞内表达的Tat蛋白可激活潜伏的KSHV 的复制周期,并促进Kaposin蛋白介导的类KS肿瘤形成[7-8]。本研究在检测重组Tat蛋白的分泌活性时,检测了293细胞分泌的Tat蛋白对KSHV 感染细胞BCBL-1细胞内LTR-CAT 质粒的调控,结果证实分泌型的Tat蛋白可以进入BCBL-1细胞,并诱导LTR-CAT 质粒中的CAT 表达。表明分泌型Tat蛋白具有穿入BCBL-1细胞的能力,并具有转录激活活性。本研究使用的BCBL-1细胞是KSHV 潜伏感染细胞系,构建重组tat基因分泌型载体对研究Tat蛋白在AIDS-KS发生机制中的作用有重要意义。
[1]Barillari G,Ensoli B.Angiogenic effects of extracellular human immunodeficiency virus type 1Tat protein and its role in the pathogenesis of AIDS-associated Kaposi's sarcoma[J].Clin Microbiol Rev,2002,15(2):310-326.
[2]卢春,钱超,唐桂霞,等.含HIV-1Tat基因重组反转录病毒表达载体构建与表达Tat蛋白功能检测[J].南京医科大学学报:英文版,2003,17(6):261-269.
[3]Jeang KT,Xiao H,Rich EA.Multifaceted activities of the HIV-1transactivator of transcription,Tat[J].J Biol Chem,1999,274(41):28837-28840.
[4]Fujisawa J,Seiki M,Kiyokawa T,et al.Functional activation of the long terminal repeat of human T-cell leukemia virus type I by a trans-acting factor[J].Proc Natl Acad Sci U S A,1985,82(8):2277-2281.
[5]Ensoli B,Barillari G,Salahuddin SZ,et al.Tat protein of HIV-1stimulates growth of cells derived from Kaposi's sarcoma lesions of AIDS patients[J].Nature,1990,345(6270):84-86.
[6]Xiao H,Neuveut C,Tiffany HL,et al.Selective CXCR4 antagonism by Tat:implications for in vivo expansion of coreceptor use by HIV-1[J].Proc Natl Acad Sci U S A,2000,97(21):11466-11471.
[7]Zeng Y,Zhang X,Huang Z,et al.Intracellular tat of human immunodeficiency virus type 1activates lytic cycle replication of kaposi's sarcoma-associated herpesvirus:role of JAK/STAT signaling[J].J Virol,2007,81(5):2401-2417.
[8]Chen X,Cheng L,Jia X,et al.Human immunodeficiency virus type 1 Tat accelerates Kaposi sarcoma-associated herpesvirus Kaposin A-mediated tumorigenesis of transformed fibroblasts in vitro as well as in nude and immunocompetent mice[J].Neoplasia,2009,11(12):1272-1284.
