盐胁迫对菊芋干物质和糖分积累分配的影响
2014-11-07李辉康健赵耕毛尹晓明梁明祥
李辉,康健,赵耕毛,尹晓明,梁明祥*
(1.南京农业大学资源与环境科学学院,江苏 南京 210095;2.江苏省海洋生物学重点实验室,江苏 南京 210095)
菊芋(Helianthustuberosus)为向日葵属的多年生植物,菊芋块茎是重要的经济利用器官,块茎的主要成分为菊粉(果聚糖),占块茎干物质含量的50%以上。菊芋具有较高的经济价值和应用前景,利用其块茎生产的低聚果糖已广泛应用于食品和医药行业。近年来,菊芋块茎可用于发酵生产燃料乙醇、生物柴油等新兴生物质能源[1-3]。
碳水化合物是植物光合同化作用的主要产物,光合同化产物在植物体内的积累和分配与产量密切相关[4]。同化物在植物体内的合成、运输和分配受植物生长发育的调控,同时也受环境因素的影响[5]。菊芋块茎的贮藏物质来源于地上部(源),地上部干物质的积累与转运对块茎产量起重要的调控作用,另外,块茎(库)对同化物的竞争能力也直接影响到同化物在植物体各器官之间的分配[6]。在盐逆境下菊芋块茎减产相较地上部更为明显,然而,盐逆境影响菊芋块茎产量的原因和生理机制尚不清楚。叶片是植物糖分的主要来源地,蔗糖是光合同化产物的主要运输形式,同时蔗糖也是果聚糖合成的原料。菊芋叶片和茎中的蔗糖主要向块茎中转运,果聚糖则主要在块茎中合成并贮藏[7-9]。目前的研究对菊芋块茎中糖分的积累和变化关注较多,而对地上部的糖分积累动态及转运分配机制的研究相对较少,盐胁迫下糖分在菊芋各器官中的积累动态和分配机制目前尚未见报道。
本研究通过全生育期试验,研究盐胁迫对两菊芋品种干物质积累与分配的影响,揭示盐胁迫下干物质积累的动态变化规律和干物质在地上部与块茎之间的分配格局,从同化物的“源-库”分配角度来说明盐逆境使块茎减产的原因,为耐盐菊芋品种选育和栽培调控提供参考依据。
1 材料与方法
1.1 试验材料
选用菊芋品种“南芋一号”(H.tuberosuscv. NY-1)和“青芋二号”(H.tuberosuscv. QY-2)为试材,两个品种的块茎均采自南京农业大学863试验基地江苏省大丰市金海农场。
1.2 试验设计
试验于2011年在南京农业大学牌楼温室试验基地进行。试验采用土培盆栽方式,盆栽用土为滨海地区(江苏大丰)含盐量较低的耕层0~20 cm砂壤,所用土壤的基本性质为有机质14.56 g/kg,有效磷65 mg/kg,速效钾24 mg/kg,碱解氮51 mg/kg,盐分0.21 g/kg,pH值8.1。选用底端有孔、透气性良好的塑料桶(直径30 cm、深30 cm)进行盆栽,底端垫塑料托盘。每盆装干土10 kg,施10 g复合肥作为底肥,复合肥总养分为45%,不含氯,N∶P∶K=15∶15∶15。试验用菊芋块茎种子预先在室内进行发芽,切取大小一致的块茎芽眼播于装有石英砂的塑料周转箱中,芽眼朝上,于光照培养箱中培养,昼夜温度分别为30和20℃,光暗周期为16 h∶8 h,保持砂质的湿润。待块茎萌发后,选取长势一致的幼苗进行移栽,每盆3株,待幼苗长势稳定后定苗,每盆留取1株进行盐处理。
试验设2个处理水平:对照(CK)和盐处理(T),根据预实验的研究结果(数据未列出),选取2‰作为盐处理浓度,向土壤中浇灌NaCl稀溶液,使土壤含盐量达到2 g/kg,土壤含盐量(g/kg)=(加入盐量+原始土盐量)/土壤质量×1000。充分供水,使土壤含水量达到最大持水量的60%,同时清洗托盘,将水浇入盆中,以保证盐分不流失。
1.3 采样时间与方法
在移栽后的50,100,140,180,220 d (days after planting)分别取样1次,在第50天取样时只取叶片,在第220天,因叶片掉落,没有对叶片取样,每个处理每次取3株作为重复样。每个时期都将地上部与块茎分开,地上部烘干称干重;块茎称量鲜重,烘干后称干重计算干物质含量;取功能叶片(倒三、四、五)、茎基部(5~10 cm)和块茎烘干用于糖分测定。
1.4 干物质积累速率与分配比例的计算
根据块茎各生长发育时期的干重和间隔时间,计算块茎的干物质积累速率
块茎干物质含量(%)=干重/鲜重×100
块茎干物质积累速率(g/株·d)=(后一期干重-前一期干重)/间隔时间
地上部干物质分配比例(%)=地上部干重/全株干重×100
块茎干物质分配比例(%)=块茎干重/全株干重×100
1.5 糖分含量的测定
可溶性总糖(total soluble sugar,TSS)含量的测定采用苯酚-硫酸法[10]。
总还原糖(total reducing sugar,TRS)含量的测定采用DNS法[11]。
茎基部非还原糖(non-reducing sugar)和块茎果聚糖(fructan)含量的测定采用总糖减去还原糖的方法[12]。
1.6 块茎果聚糖积累量和果聚糖积累速率的计算
块茎果聚糖积累量(mg/g)=果聚糖含量(mg/g)×干物质积累量(g)/1000
块茎果聚糖积累速率(mg/株·d)=(后一期果聚糖积累量-前一期果聚糖积累量)/间隔时间
1.7 数据处理
利用Microsoft Excel软件、SPSS 16.0软件进行试验数据的统计分析,统计学分析使用的是单因素方差分析(OneWay ANOVA)。图片采用SigmaPlot软件绘制。
2 结果与分析

图1 盐胁迫对地上部干物质积累动态的影响Fig.1 Effect of salt stress on the dynamics of dry matter accumulations in shoots CK:对照;T: 盐处理(2‰);*:显著性差异。下同。CK: Control; T: Salt stress (2‰); *: Significant difference. The same below.
2.1 盐胁迫对地上部干物质积累动态的影响
菊芋块茎起始形成后地上部干物质积累动态表明,地上部干物质积累呈单峰曲线变化(图1),两个菊芋品种出现峰值的时间都在140 d,块茎起始形成后,地上部干物质积累量呈增加趋势,随着块茎的逐渐膨大,地上部干物质开始向地下转运。土壤盐处理没有改变两个菊芋品种地上部干物质积累达到高峰的时间,但却显著降低了两个品种地上部干物质的积累量,NY-1和QY-2地上部最大干物质积累量降低幅度分别为44.85%和43.45%。盐胁迫下,NY-1在100~140 d地上部干物质积累速率低于对照,于140~220 d地上部干物质转运速率也明显低于对照;QY-2地上部干物质积累动态与对照差异不明显。以上结果表明,盐胁迫显著降低了两菊芋品种地上部干物质的积累量,盐胁迫虽没有改变地上部干物质积累的动态变化趋势,却限制了块茎膨大过程中地上部贮藏物质的积累速率和向外转运的速率。
2.2 盐胁迫对块茎干物质积累动态的影响
由图2A可知,NY-1块茎干物质积累量呈持续增加趋势;QY-2块茎干物质在100~180 d积累甚少,其后才有显著上升,说明不同品种块茎干物质积累动态存在明显差异。土壤盐处理下,块茎干物质积累量明显减少,块茎成熟期(220 d),NY-1块茎干物质积累量降幅(57.78%)低于QY-2(85.61%)。在块茎生长发育过程中,两个菊芋品种块茎干物质含量(图2B)的变化趋势不同,NY-1块茎干物质含量在180 d达峰值,呈先上升后下降趋势;QY-2块茎干物质含量呈持续上升趋势,在220 d达到最高。盐处理明显降低了两个菊芋品种块茎的干物质含量;盐处理下,两菊芋品种块茎干物质含量均在140 d达到最高,都呈先上升后下降趋势。以上结果表明,盐胁迫显著降低了块茎干物质的积累量,而且使块茎干物质含量的峰值提前,限制了块茎中贮藏物质的持续有效积累。

图2 盐胁迫对块茎干物质重(A)和干物质含量(B)积累动态的影响Fig.2 Effect of salt stress on the dynamics of dry matter weights (A) and dry matter content (B) accumulations in tubers
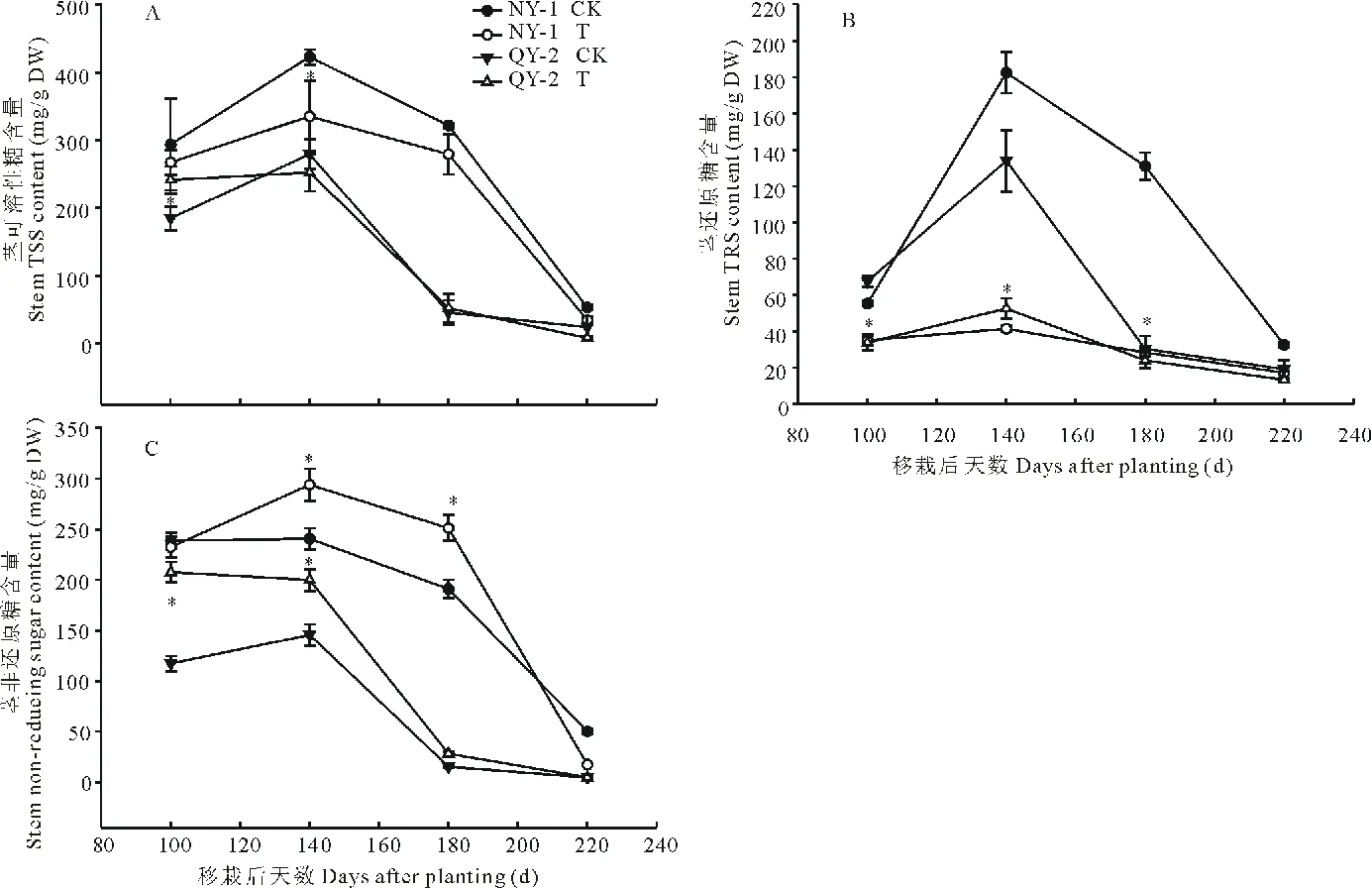
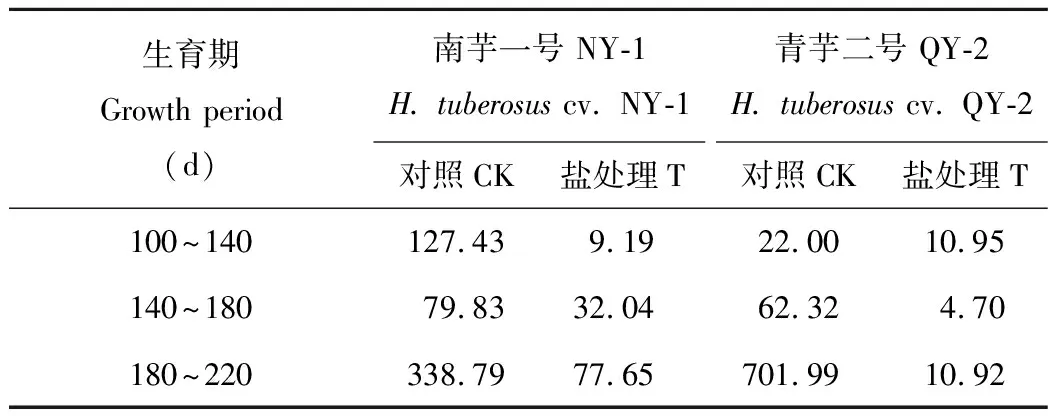
表1中的结果表明,两个菊芋品种的块茎干物质积累速率变化有明显的差异,NY-1块茎干物质积累速率呈“快-慢-快”的变化趋势,即在块茎形成后,块茎干物质有两个快速积累时期,分别在100~140 d和180~220 d;QY-2块茎干物质积累速率在100~180 d都很低,在180~220 d,块茎干物质积累速率才有明显提高,说明QY-2块茎干物质的积累主要集中在块茎膨大的后期。土壤盐处理使两菊芋品种的块茎干物质积累速率都显著降低,NY-1在180~220 d的块茎干物质积累速率才与对照100~140 d的相当,QY-2在整个块茎发育过程中的干物质积累速率都较慢;在180~220 d,块茎干物质积累速率降幅NY-1(29.19%) 表1 盐胁迫对块茎干物质积累速率的影响Table 1 Effect of salt stress on accumulation rates of tuber dry matter weight per plant g/(株Plant·d) CK: 对照 Control;T: 盐处理(2‰) Salt stress (2‰). 下同The same below. 如图3A所示,除NY-1对照外,地上部干物质的分配比例呈先上升后下降趋势,NY-1对照呈持续下降趋势。盐处理下,地上部干物质的分配比例明显高于对照。在块茎发育过程中,块茎干物质分配比例(图3B)的变化趋势与块茎干物质积累量基本相似,NY-1块茎干物质分配比例持续增加,QY-2块茎干物质分配比例呈先下降后上升趋势。盐胁迫显著降低了两菊芋品种的块茎干物质分配比例,块茎成熟期(220 d)NY-1和QY-2块茎干物质分配比例分别比对照降低了42.43%和68.27%,降幅NY-1 图3 盐胁迫对地上部(A)与块茎(B)干物质分配比例的影响Fig.3 Effect of salt stress on the dry matter distribution ratios of shoot (A) and tuber (B) 菊芋叶片中总可溶性糖(图4A)和还原糖(图4B)含量的变化总趋势基本相同。在对照中,NY-1叶片中总可溶性糖和还原糖含量都呈先上升后下降趋势,在140 d达到高峰;QY-2叶片中总可溶性糖含量在50~100 d呈上升趋势,在100~180 d趋于平稳,而QY-2叶片中还原糖含量在100 d出现峰值;NY-1叶片中总可溶性糖和还原糖含量的最大值均高于QY-2。 土壤盐处理未改变叶片中总可溶性糖和还原糖含量的总体变化趋势;在50~100 d,NY-1的总可溶性糖和还原糖含量有所增加,在100~180 d,叶片中总可溶性糖和还原糖含量低于对照;QY-2的总可溶性糖和还原糖含量只有在50 d时高于对照。盐胁迫下,在100~140 d,两个品种叶片的总可溶性糖和还原糖积累速率都低于对照;在140~180 d,NY-1叶片中可溶性糖的输出速率低于对照,而QY-2叶片中可溶性糖的下降幅度与对照无明显差异。以上结果表明,盐胁迫提高了菊芋快速增长期(50~100 d)叶片中的总可溶性糖和还原糖含量,但在块茎形成后(100~180 d),叶片中可溶性糖的积累和输出均受到抑制。 菊芋块茎形成后,茎基部的总可溶性糖(图5A)、还原糖(图5B)和非还原糖(图5C)含量均呈单峰曲线变化,在100~140 d总可溶性糖含量上升, 至140 d达到峰值,其后下降。盐处理没有改变茎基部中糖组分含量的变化趋势,但降低了茎基部的总可溶性糖和还原糖含量,同一品种盐处理与对照同期相比,茎基部还原糖含量的下降幅度显著大于总可溶性糖;盐胁迫下茎基部糖分向外输出的速率也有所下降;然而,盐处理却使茎基部的非还原糖含量在100~180 d都明显高于对照。两个品种中,盐胁迫下NY-1和QY-2茎基部的最大总可溶性糖含量分别比对照降低了20.74%和9.66%,降幅NY-1> QY-2;茎基部最大还原糖含量的降幅NY-1(77.34%)>QY-2(60.79%)。以上结果表明,盐胁迫虽没有改变茎基部糖分积累和向外输出的时间,但糖含量有所下降,其中还原糖的下降幅度很大,然而,盐处理却诱导了非还原糖在茎基部有较多的积累。 图4 盐胁迫对叶片总可溶性糖(A)和还原糖(B)含量的影响Fig.4 Effect of salt stress on total soluble sugar (A) and reducing sugar (B) contents in leaves 图5 盐胁迫对茎基部总可溶性糖(A)、还原糖(B)和非还原糖(C)含量的影响Fig.5 Effect of salt stress on total soluble sugar (A), reducing sugar (B) and non-reducing sugar (C) contents in stem bases 如图6A所示,在块茎膨大过程中,两个菊芋品种在非盐害条件下块茎的总可溶性糖含量呈持续上升趋势。在盐胁迫下,块茎中总可溶性糖含量呈先增加后减少趋势,NY-1块茎中总可溶性糖含量在140 d最高,QY-2在180 d达到最高值;在100~140 d,NY-1块茎总可溶性糖含量稍高于对照,在140~220 d,其总可溶性糖含量显著低于对照;QY-2块茎可溶性糖含量在100~180 d高于对照,而在220 d显著低于对照。由此说明,盐胁迫有利于膨大前、中期块茎总可溶性糖含量的升高,而在块茎膨大后期,块茎总可溶性糖含量的积累明显受到限制。 图6 盐胁迫对块茎总可溶性糖(A)、还原糖(B)和果聚糖(C)含量的影响Fig.6 Effect of salt stress on total soluble sugar (A), reducing sugar (B) and fructan (C) contents in tubers 在块茎膨大过程中,两个菊芋品种块茎中还原糖含量(图6B)都呈先上升后下降趋势,在100~140 d,还原糖含量增加,其后开始下降。盐处理条件下,两菊芋品种块茎中还原糖含量都显著低于对照,NY-1块茎中还原糖含量在140~220 d的下降幅度都较大,而QY-2只在100~140 d有较大的降幅。 由于块茎中主要成分为果聚糖,因此在块茎中可以近似认为果聚糖为可溶性总糖与还原糖的差值。如图6C所示,块茎膨大过程中果聚糖含量的变化趋势与总可溶性糖含量一致,说明块茎中积累的糖分主要是果聚糖。同样,盐胁迫使果聚糖含量积累的时间提前,NY-1在140 d的果聚糖含量显著高于对照;QY-2在100~140 d的果聚糖含量显著高于对照;从180 d后,两菊芋品种块茎中果聚糖含量开始下降,均显著低于对照。以上结果表明,盐胁迫改变了块茎中果聚糖含量积累的时间,限制了块茎中果聚糖的持续有效积累。 图7 盐胁迫对块茎果聚糖积累量的影响Fig.7 Effect of salt stress on fructan accumulations in tubers 在块茎发育过程中,两个菊芋品种块茎果聚糖积累量均呈逐渐增加趋势(图7),在220 d果聚糖积累量达到最大值。盐处理显著降低了两个品种块茎中果聚糖的积累量,至块茎成熟(220 d),NY-1和QY-2块茎中果聚糖积累量分别比对照降低了74.02%和93.81%,降幅NY-1 由表2可知,在对照中,NY-1在块茎发育过程中的果聚糖积累速率呈“快-慢-快”的趋势;QY-2在100~180 d的果聚糖积累速率较低,在180~220 d,其果聚糖积累速率显著增加。由此说明,不同菊芋品种块茎中果聚糖积累速率变化趋势不同,NY-1有两个果聚糖快速积累期,而QY-2的果聚糖积累主要集中在块茎膨大后期。盐处理条件下,两个菊芋品种的块茎果聚糖积累速率明显下降,最大降幅NY-1(77.08%) 表2 盐胁迫对块茎果聚糖积累速率的影响Table 2 Effect of salt stress on fructan accumulation rates in tubers mg/(株Plant·d) 干物质积累是光合作用碳同化产物在植物体内贮藏的主要表现形式。菊芋的一个典型特征就是先在地上部积累储存同化物然后再向地下块茎转运[8-12]。本研究表明,在块茎起始形成后,地上部干物质的积累动态呈先增加后减少趋势,也表现出先积累后转运的特征。土壤盐处理没有改变两个菊芋品种地上部干物质积累达到高峰的时间,但却显著降低了两个品种地上部干物质的积累量,NY-1和QY-2地上部最大干物质积累量降低幅度分别为44.85%和43.45%。盐胁迫下,NY-1在100~140 d地上部干物质积累速率低于对照,于140~220 d地上部干物质转运速率也明显低于对照;QY-2地上部干物质积累动态与对照差异不明显。盐胁迫显著降低了两菊芋品种地上部干物质的积累量,盐胁迫虽没有改变地上部干物质积累的动态变化趋势,却限制了块茎膨大过程中地上部贮藏物质的积累速率和向外转运的速率。 菊芋在块茎发育过程中,块茎不断膨大,干物质含量和积累总量随生长的进行而增加,且与收获季节有关[13]。由图2A可知说明不同品种块茎干物质积累动态存在明显差异。本研究表明,块茎起始形成后,块茎干物质积累量呈增加趋势,在非盐胁迫下,NY-1块茎的干物质积累速率呈“快-慢-快”的变化趋势,表明其块茎有两个较快的膨大时期(100~140 d和180~220 d),而QY-2块茎干物质的积累主要集中在块茎膨大后期(180~220 d);钟启文等[14]对“青芋1号”生长发育动态研究表明其块茎膨大速率在第18周和第21周有2个高峰;由此说明,菊芋块茎的发育动态因品种的不同而有所差异;造成差异的原因可能与地上部干物质向地下块茎转运的时间和速率有关,本研究结果显示NY-1地上部干物质向块茎转运的时间是在140 d后,而QY-2在180 d后才开始转运,而且NY-1的转运速率大于QY-2;另外,由于地下块茎的生长发育不易观察,对菊芋的生长发育时期尚未有清晰完整的定义,所以在与文献报道的结果进行比较时可能会有较大的差异。土壤盐处理下,块茎干物质积累量明显减少,块茎成熟期(220 d),NY-1块茎干物质积累量降幅(57.78%)低于QY-2(85.61%)。盐处理明显降低了两个菊芋品种块茎的干物质含量;盐胁迫显著降低了块茎干物质的积累量,而且使块茎干物质含量的峰值提前,限制了块茎中贮藏物质的持续有效积累。 干物质的积累与分配直接关系到植物的目标产量,菊芋的块茎产量最终取决于同化产物在植物体各个部分的分布与再分配,一方面取决于同化产物的来源是否充足,另一方面,也取决于其自身的生长潜力(库强)[8]。本研究中,盐处理降低了菊芋地上部和块茎的干物质积累量,而且地上部干物质的积累和转运速率也都有所下降。地上部作为块茎同化物的主要来源,其干物质的积累与转运直接影响到块茎干物质的积累,相关性分析结果也表明,地上部干重与块茎干重呈显著正相关,因此,盐胁迫降低菊芋块茎产量的一个重要原因是减少了其地上部同化物的供给。植物库器官对同化物的竞争能力或者说对同化物的利用能力可能决定了植物体内同化物的分配[4]。在菊芋块茎发育过程中,块茎的干物质积累速率在一定程度上决定了其同化物有效积累的潜力[8]。盐胁迫显著降低了块茎的干物质积累速率,说明盐胁迫限制了块茎中干物质的有效快速积累,从而降低其最终产量。因此,盐胁迫从“源”和“库”两方面限制了菊芋块茎中同化物的积累,即地上部来源减少和块茎库强变弱。 植物同化物的分配主要受植物遗传特性、生理过程和环境因子的影响[4]。盐胁迫、干旱等逆境胁迫可改变干物质在植物各个器官的分配[15-19]。本研究结果表明,盐胁迫改变了菊芋干物质的分配格局,地上部分配比例增大而块茎分配比例减小,且块茎干物质积累量的降幅比地上部大,说明盐胁迫对块茎的限制作用更大,Newton[7]基于大田重粘土盐分对块茎产量的影响研究表明,盐胁迫对块茎的影响比地上部更大。相关性分析结果显示,地上部分配比例与块茎干重呈极显著负相关,块茎分配比例与块茎干重呈极显著正相关,说明盐胁迫下菊芋干物质的分配格局由块茎的干物质积累量所主导。 本研究中,盐胁迫对两个菊芋品种干物质积累与分配的影响明显不同。盐处理下,虽然NY-1地上部干物质最大积累量的降幅大于QY-2,但NY-1块茎干重和干物质积累速率的降幅却都小于QY-2,说明盐胁迫对QY-2块茎产量的影响更加明显,其原因可能是盐胁迫对QY-2块茎膨大的限制作用更大,不利于地上部干物质向块茎的转运。 可溶性糖是植物光合产物的主要形式,在植物体内的合成、运输和分配受植物生长发育的调控,在植物生长发育的不同时期,各营养器官中的可溶性糖含量会发生规律性的变化,同时也受到环境因素的影响[5]。在菊芋的生长发育过程中,叶片是糖分的主要来源,茎秆作为糖分的临时贮藏器官,而块茎的糖分只能来源于地上部的转运,糖分在茎中的积累和再分配对块茎的生长形成起重要的调节作用[8-9,18]。本研究结果表明,叶片和茎中的可溶性总糖含量均呈单峰曲线变化,在块茎形成后,茎叶中的总糖含量开始减少,糖分逐渐向地下块茎转运,盐胁迫没有改变茎叶中糖分的变化趋势,表明盐胁迫并没有影响地上部糖分的积累转运时间。 菊芋叶片中总可溶性糖和还原糖含量的变化总趋势基本相同。在对照中,NY-1叶片中总可溶性糖和还原糖含量都呈先上升后下降趋势,在140 d达到高峰;QY-2叶片中总可溶性糖含量在移栽后100 d内呈上升趋势,在100~180 d趋于平稳,而QY-2叶片中还原糖含量在100 d出现峰值;NY-1叶片中总可溶性糖和还原糖含量的最大值均高于QY-2。通过HPLC-ELSD检测发现叶中主要是还原糖。土壤盐处理未改变叶片中总可溶性糖和还原糖含量的总体变化趋势;盐胁迫下,在100~140 d,两个品种叶片的总可溶性糖和还原糖积累速率都低于对照;在140~180 d,NY-1叶片中可溶性糖的输出速率低于对照,而QY-2叶片中可溶性糖的下降幅度与对照无明显差异。盐胁迫提高了菊芋快速增长期(移栽后100 d内)叶片中的总可溶性糖和还原糖含量,但在块茎形成后(100~180 d),叶片中可溶性糖的积累和输出均受到抑制。 菊芋茎秆中贮藏的可溶性碳水化合物主要是果聚糖、蔗糖及果糖等[19,21-22],果聚糖在茎中临时性储存,其含量在不同生理期的变化很大,茎中的最大糖含量可占干物质的25%~70%,蔗糖是糖分在植物体内的主要转运形式[9,18,20]。外界环境因素可以通过改变茎秆中各糖组分的比例,从而影响糖分在植物体内的分配与再分配。本研究表明,盐处理降低了茎中的总糖含量,其中还原糖含量下降极为明显,但是,茎中的非还原糖(果聚糖、蔗糖)含量反而高于对照。研究表明,果聚糖除作为贮藏性碳水化合物外,还是重要的渗透调节物质,果聚糖可以保护质膜不被冷害、干旱等胁迫损伤[23-27]。那么盐胁迫使茎中果聚糖等非还原性糖含量升高,其原因可能是通过增加渗透调节物质而使其抵抗胁迫的能力增强。但从另一个角度来看,果聚糖比例的升高意味着糖分的去向发生改变,茎中果聚糖含量升高,相对就会减少果聚糖向地下块茎的再转运,减少了块茎糖分的来源,从而限制块茎的膨大和糖分的积累。同一品种盐处理与对照同期相比,茎基部还原糖含量的下降幅度显著大于总可溶性糖;盐胁迫下茎基部糖分向外输出的速率也有所下降;然而,盐处理却使茎基部的非还原糖含量在100~180 d都明显高于对照。两个品种中,盐胁迫下NY-1和QY-2茎基部的最大总可溶性糖含量分别比对照降低了20.74%和9.66%,降幅NY-1> QY-2;茎基部最大还原糖含量的降幅NY-1(77.34%)>QY-2(60.79%)。盐胁迫虽没有改变茎基部糖分积累和向外输出的时间,但糖含量有所下降,其中还原糖的下降幅度很大,然而,盐处理却诱导了非还原糖在茎基部有较多的积累。 在块茎膨大过程中,两个菊芋品种在非盐害条件下块茎的总可溶性糖含量呈持续上升趋势,而盐处理下,块茎中总糖含量的变化趋势发生改变,NY-1块茎中总可溶性糖含量在140 d最高,QY-2在180 d达到最高值;在100~140 d,NY-1块茎总可溶性糖含量稍高于对照,在140~220 d,其总可溶性糖含量显著低于对照;QY-2块茎可溶性糖含量在100~180 d高于对照,而在220 d显著低于对照,说明在块茎发育前期含量较高,表明盐胁迫诱导了块茎中糖分合成积累的提前。有研究表明,对菊芋伸长生长有抑制作用的任何因素(渗透胁迫、抗赤霉素类物质等)都可以诱导块茎形成和膨大,且这些因子具有叠加效应[8]。如此看来,盐处理所产生的渗透胁迫可能诱导了菊芋块茎形成提前,而使其糖含量在块茎形成初期比对照高,但盐胁迫降低了块茎发育后期的糖含量,不利于块茎中糖分的持续有效积累。在块茎膨大过程中,两个菊芋品种块茎中还原糖含量(图6B)都呈先上升后下降趋势,在100~140 d,还原糖含量增加,其后开始下降。盐处理条件下,两菊芋品种块茎中还原糖含量都显著低于对照,NY-1块茎中还原糖含量在140~220 d的下降幅度都较大,而QY-2只在100~140 d有较大的降幅。由于块茎中主要成分为果聚糖,因此在块茎中可以近似认为果聚糖为可溶性总糖与还原糖的差值。如图6C所示,块茎膨大过程中果聚糖含量的变化趋势与总可溶性糖含量一致,说明块茎中积累的糖分主要是果聚糖。同样,盐胁迫使果聚糖含量积累的时间提前,NY-1在140 d的果聚糖含量显著高于对照;QY-2在100~140 d的果聚糖含量显著高于对照;从180 d后,两菊芋品种块茎中果聚糖含量开始下降,均显著低于对照。盐胁迫改变了块茎中果聚糖含量积累的时间,限制了块茎中果聚糖的持续有效积累。 总之,盐胁迫大大降低了两个菊芋品种的生物量,改变了植物体内的糖分分布模式,从“源”和“库”两方面限制了菊芋块茎中同化物的积累。盐处理下,不同菊芋品种块茎受到的胁迫程度不同,NY-1块茎干物质积累量和积累速率相对较高,QY-2块茎产量和干物质积累速率下降幅度较大,说明NY-1可能比较适合在盐碱地的种植。以上研究对于以后耐盐菊芋品种的开发和盐碱地的开发利用提供了重要的理论依据。
2.3 盐胁迫对地上部与块茎干物质分配比例的影响

2.4 盐胁迫对菊芋叶片中糖含量的影响
2.5 盐胁迫对菊芋茎基部中糖含量的影响

2.6 盐胁迫对菊芋块茎中糖含量的影响

2.7 盐胁迫对菊芋块茎中果聚糖积累量及其积累速率的影响

3 讨论
3.1 盐胁迫对菊芋干物质积累与分配的影响
3.2 盐胁迫对菊芋糖分积累与分配的影响
