毛乌素沙南缘紫穗槐根系垂直分布特征
2014-11-07杜明新周向睿周志宇卢鑫梁坤伦周媛媛陶晓慧
杜明新,周向睿,周志宇,卢鑫,梁坤伦,周媛媛,陶晓慧
(1.草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州 730020;2.甘肃农业大学草业学院,甘肃 兰州 730070)
紫穗槐(Amorphafruticosa)又名:棉槐、棉条、穗花槐,是一种喜光,耐寒、耐旱、耐湿、耐盐碱、耐沙压、抗逆性极强的多年生豆科丛生小灌木,属半旱生植物。是一种毛乌素沙南缘沙地营造水土保持林和薪炭林的主要灌木。许多学者对紫穗槐做了大量的研究,主要集中在固沙效益[1],形态特征[2],抗性研究及营养化学成分的提取[3-4],但是对于紫穗槐根系分布特征研究还鲜见报道。邹丽娜等[3]研究了盐分胁迫对紫穗槐幼苗生理生化特性的影响;梁坤伦等[2]对青藏高原紫穗槐主要形态特征进行了变异分析;周心澄[1]对紫穗槐固沙造林方法和固沙效益进行定位观察。
根系研究最早是由Hales提出的,经过多年的发展形成:1)根系形态学研究[5],2)根系生理学研究[6],3)根系环境胁迫研究[7],4)根际研究[8],5)根系生态学研究[9-10]。周艳松等[5]研究分析大针茅(Stipagrandis)根系构型在不同退化程度的草地中的变化情况;余爱等[6]研究认为不同施磷浓度对柱花草(Stylosanthesguianensias)和黑籽雀稗(Paspalumatratum)根系分布有一定的影响;任伟等[8]对阿拉善荒漠灌木根际中、微量元素含量特征进行了分析;胡建忠等[10]对退耕地人工植物群落根系生态位及其分布特征进行了研究。植物根系生长依赖地上部生长状况、土壤环境、介质层次等因素,而且具有适应性和可塑性的功能器官。良好的根系分布特征,不仅提高了根系对土壤养分和水分利用效率,也是构建稳定群落的基础。因此,以毛乌素沙南缘不同年限种植紫穗槐的根系为研究对象,探明紫穗槐在沙地生境下根系分布特征。通过测定紫穗槐根系分布参数及地下分配的变化,充分揭示紫穗槐对沙地生境的生态适应性,固沙能力及水分吸收利用效率。了解不同年限紫穗槐根系生长规律,就能够模拟植被根系在整个生活史中的空间分布,这不仅为草地营养学、恢复生态学等学科的发展提供重要的科学依据和理论基础,而且也对荒漠化治理中有效种植灌木具有特别重要的意义。
1 材料与方法
1.1 试验地概况
试验区位于榆林市靖边县毛乌素沙南缘, 东经107°28′~111°15′,北纬36°57′~39°34′,海拔980~1534 m。平均温度7.9℃,绝对最高温度37.6℃,绝对最低温度-27℃,无霜期150 d,年平均降水量438.4 mm。地貌主要为流动、半固定或固定沙地;土壤类型为各类风沙土;植被类群主要为沙生灌木:沙柳(Salixpsammophila)、杂交杨(hybrid yang)、紫穗槐、沙蒿(Artemisiaordosica)、沙葱(Alliummongolicum)、酸刺(acidthorn)、苦豆子(Sophoraalopecuroides)、沙枣(Elaeagnusangustifolia)、花棒(Hedysarumscoparium)、沙打旺(Astragalusadsurgens)、柳树(Salixbabylonica)、柠条(Caraganakorshinskii)、沙蓬(Agriophyllumsquarrosum)等。
1.2 研究方法
2011年8月在榆林市靖边县毛乌素沙南缘,选取1951年(60龄)、1976年(35龄)、1983年(28龄)、2005年(6龄)种植的紫穗槐各3块样地,在每块样地内随机选取5株中等大小样株,测量株高、枝条数(WN)、枝条直径、枝条长度、冠幅,然后在所挖掘的植株附近取4个点,利用土钻以20 cm为一层,分层取5个土样(将同层4个点的样品混合均匀),测定其土壤含水量(SW)。采用全株挖掘法, 按土壤层次(每20 cm为一层, 80 cm以下合并)挖出根系,除去土壤。将根系带回实验室进行冲洗, 随机选取5段测根系的平均直径,然后用量筒溢水法测出各层的体积(RV),用公式L=4V/πd2和S=4V/d求出总根长(RL)和根系表面积(RSA)。最后,将各层根系与植株地上部分放入80℃烘箱,烘干至恒重,计为根系生物量(RB)[11]。比根长和根长密度计算公式:比根长 (SRL)=L/W;根长密度(RLD)=L/V。式中,SRL:比根长(cm/g);RLD:根长密度(m/m3);W:干根重(g);L:根长(m);V:土壤体积(cm3)。根比例=某层根面积/总根面积。根系消减系数Y=l-βh。式中,Y为从地表到一定深度的根系生物量累积百分比(%);h为土层深度(cm); β为根系消弱系数[12]。
1.3 数据分析
采用SPSS 18.0和 Excel 2007进行数据统计分析。用单因素方差分析(one way-ANOVA)和 Duncan 多重比较检验土层间、各龄间测量值差异性, Pearson 双侧检验根系分布参数之间的相关性,用主成分分析法确定影响根系分布的主要因素。
2 结果与分析
2.1 紫穗槐地上部分特征
60龄紫穗槐株高、地上生物量、枝条长度、根冠比、茎长比根长等形态指标均显著(P<0.05)大于其他株龄的相应指标(表l),28龄、6龄紫穗槐枝条数、枝条直径均显著(P<0.05)大于35龄。
2.2 土壤水分特征
水分在干旱环境中是主要限制因子,决定着荒漠植物的数量和种类[14]。通过对土壤垂直分层可以研究根系对土壤水分的利用情况,以及根系在水分的影响下所采取的适应策略[29]。不同株龄紫穗槐根系土壤含水率测定表明(图1):随着土层深度的加深不同株龄紫穗槐根系土壤含水率出现先增大后减小的趋势,最高点出现在20~40 cm和40~60 cm;0~20 cm土壤含水率显著低于20~40 cm(P<0.05);60龄紫穗槐根系各层土壤含水率显著高于其他株龄(P<0.05),其最高含水率为4.91%,是35龄、28龄、6龄最高含水率的1.62,1.64,2.05倍。

表1 紫穗槐地上部分形态特征Table 1 A. fruticosa morphological characteristics of aerial part
不同字母表示差异显著(P<0.05)。Different letters mean significant difference at 0.05 level.下同。The same below.
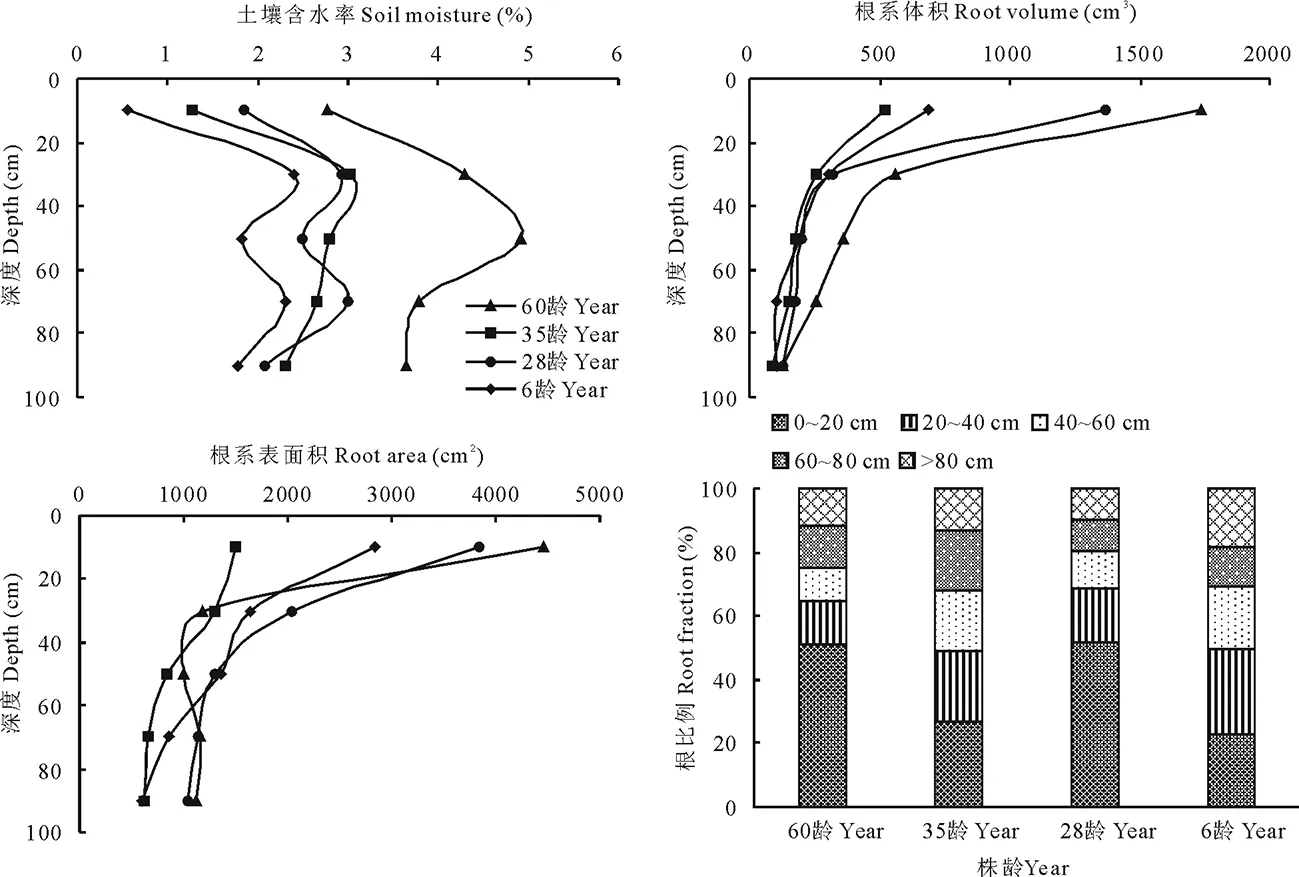
图1 不同株龄紫穗槐土壤含水率,根系体积,根表面积,根比例Fig.1 Different year aged A. fruticosa soil moisture, root volume, root surface area, root fraction
2.3 根系分布特征
2.3.1根系体积、表面积、根比例 根系体积越大,根系搜索土体内的水分和养分的范围越大,占据的资源越多,在极端条件下抗逆性也会比较强。根系表面积是根系与土体的接触面积,从某种意义上代表了植物对土壤水分、养分的吸收能力。从图1可以看出,不同株龄紫穗槐根系体积、表面积均表现为随着深度的加深呈现指数函数递减的趋势,符合指数Y=ae-bx方程(a、b为常数,Y为深度,x为根系体积、根系表面积)。0~20 cm土层60龄紫穗槐根系体积显著高于其他株龄根系体积(P<0.05),其次是28龄,6龄次之,35龄最小。根比例随着深度加深逐渐变小;60龄紫穗槐在0~20 cm根比例最大,占总根量52.29%,其次是28龄51.57%,35龄27.04%次之,6龄23.15%最低。20~40 cm根比例6龄>35龄>28龄>60龄。
2.3.2根系比根长、根长密度 比根长是衡量根直径的重要参数之一,随着直径增加而显著减小[15],在一定程度上能够反映根系的直径及其吸收能力,比根长值越大,说明根越细小[16]。从图2可以看出,随着深度的加深各株龄紫穗槐比根长总体趋于减小。0~20 cm各株龄紫穗槐比根长显著大于其他各层(P<0.05),是最低层60龄的10.67倍,35龄的1.97倍,28龄的2.43倍,6龄的5.39倍,而其他各层之间差异不显著,说明0~20 cm主要是以细根为主。0~20 cm土层6龄紫穗槐比根长显著大于其他各株龄,其次28龄,60龄次之,35龄最小。根长密度是表示根系长度在单位土体中的分布参数。0~20 cm与20~40 cm之间差异不显著,与其他各层差异显著(P<0.05)。35龄、28龄最高值在0~20 cm,根长密度分布呈典型的指数型,根长密度(x)与土层深度(Y)进行回归统计,回归方程为:35龄Y=265.74e-0.165x(R2=0.9967);28龄Y=577.9e-0.55x(R2=0.929)。6龄先增大然后逐渐减小,最高点出现在20~40 cm。60龄呈现先减小后增大,然后又减小的趋势,最高点出现在0~20 cm。
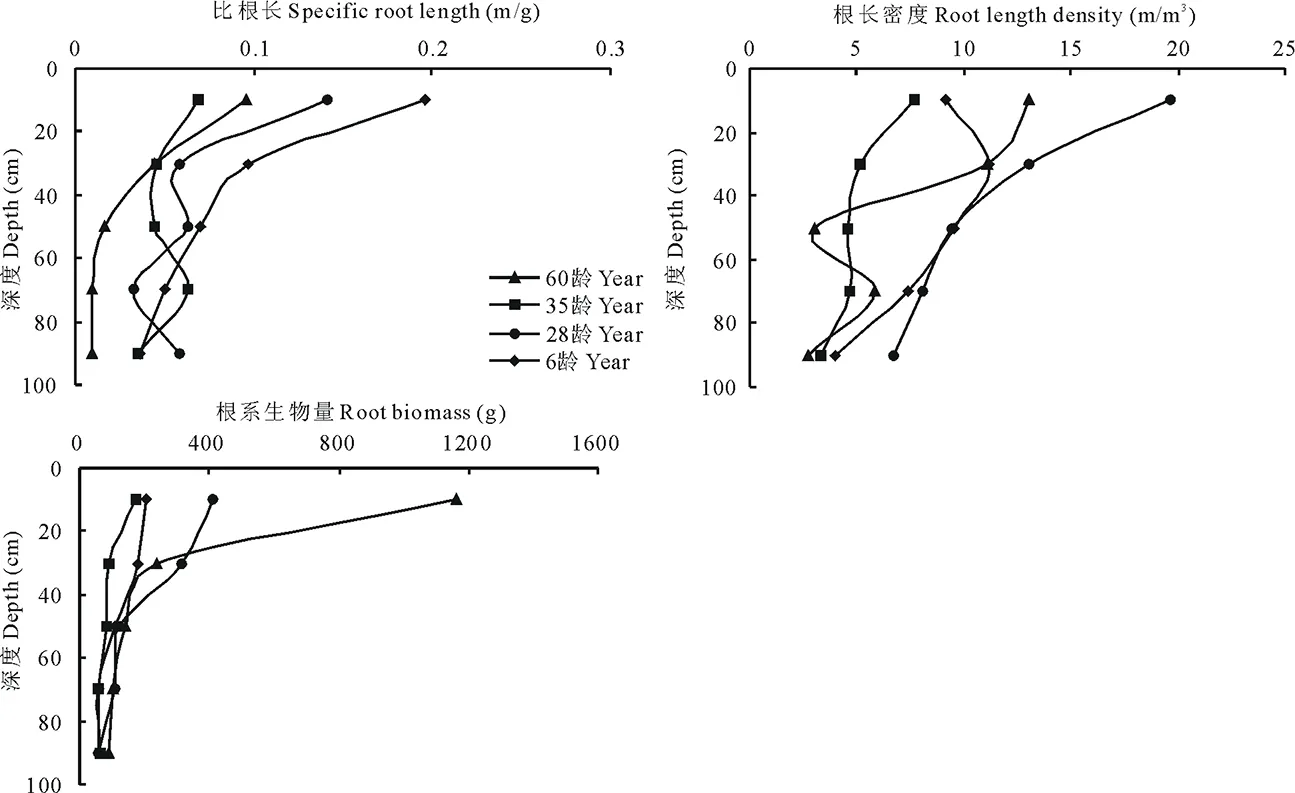
图2 不同年龄紫穗槐根系比根长、根长密度、根生物量Fig.2 Different year aged A. fruticosa specific root length, root length density, root biomass
2.3.3根系生物量、根系消弱系数 植物根系对于外界环境的改变具有可塑性,根系生物量能够随环境变化做出相应的适应, 从而直接反映根系的分布特点[17-18]。从图2可以看出,不同株龄紫穗槐根系生物量均表现为随着深度的加深呈现指数函数递减的趋势, 符合指数Y=ae-bx方程(a、b为常数,Y为深度,x为根系生物量)。 根系生物量在0~20 cm土层为集中分布区,分别占各株龄总生物量的66.8%,36.73%,40.13%和33.33%,该土层中各株龄根系生物量存在较大差异,大小依次为60龄>28龄>6龄>35龄,其中60龄是35龄的5.74倍。根据Gale对根系在土壤剖面的分布公式Y=l-βh对根系的生物量垂直分布进行统计(表2),各株龄β值总体呈现增大趋势,0~40 cm土层β值偏小,在0~20 cm土层60龄显著小于其他株龄β值(P<0.05),而在20~40 cm土层60龄显著大于其他株龄β值(P<0.05),其他各株龄各层之间差异不显著。
2.3.4相关性分析与主成分分析 从表3可以看出y1、y2主成分特征值累计贡献率达89.82%,已超过85%,说明前2个主因子基本上能概括16个变量的主要信息。从表4可以看出,不同株龄紫穗槐土壤含水量与地上生物量、根生物量、比根长的相关性达到显著水平,与根表面积呈正相关,这与周梦华等[19]的结果基本一致。根体积与根表面积、根生物量的相关性达到极显著水平,与枝条数达到显著相关;根直径与根总长、比根长呈负相关达显著水平;根总长与根表面积、比根长相关性达到显著水平。

表2 不同株龄紫穗槐根系消弱系数Table 2 Different year aged A. fruticosa root extinction coefficient
不同大写字母表示不同株龄之间差异显著(P<0.05) Different capital letters mean difference between year aged is significant at the 0.05 level; 不同小写字母表示不同深度之间差异显著(P<0.05) Different small letters mean different depth is significant at the 0.05 level.

表3 根系分布参数主成分分析Table 3 Root distributed parameter principal component analysis
3 讨论
60龄紫穗槐株高、地上生物量、枝条长度、根冠比均显著(P<0.05)大于其他株龄的相应指标表明:随着株龄增加,紫穗槐没有出现衰退的现象,其根系有自疏与更新的能力,不断适应环境的变化。由于植物器官组成成分不同,会产生地上/地下与实际植物生长不相符的情况,将茎长同根长相比,更能反映植物地上与地下关系[13]。28龄紫穗槐比值小,说明其生长的环境较为波动,植物根系搜索的土体范围较大,需要对根系投入更大的能量,以维持正常代谢。相比较之下60龄紫穗槐比值较大,说明其生长环境较为稳定,地上部分不需要对根系提供能量,生长较旺盛。Wilson[20]认为植物根与冠之间存在一种逆境中“资源优化配置”的关系。如果地下部环境不利(例如水分、养分亏缺)则光合产物分配到地下部的比例较高,以保证根系优先生长,促使根系发达,扩大与外界环境的接触面积,从而提高植物抵抗逆境的能力。本研究中枝条数目与根体积呈显著正相关,因此可以推断地上部分形态特征在一定程度上可以反映根系的分布特征。为了保证在干旱环境下生存,紫穗槐通过改变根冠比、根系伸长、增大根系的吸收面积,从而满足地上部分生长所需之水分和营养物质的需求。

表4 不同株龄紫穗槐根系分布参数之间与土壤水分相关性分析Table 4 Different year aged A. fruticosa relativity between root and soil moisture
*显著相关Correlation is significant at the 0.05 level.**极显著相关Correlation is significant at the 0.01 level.
在毛乌素沙地区,降水十分有限,又存在强蒸腾条件,水分是制约紫穗槐生长和繁殖的主要生态因子,而紫穗槐根系分布特征又决定其利用水分的策略。60龄紫穗槐对土壤贮水量有明显的改善,这可能与紫穗槐根系不断的进行自我更新有关。衰老根在微生物的作用下形成腐殖质, 产生的腐殖质,可以增加沙土的团聚结构,减少毛细管的孔隙度,同时衰老根还是地表水进入地下的孔隙和通道,使根系可以充分利用深层土壤水。Burton等[22]证实土壤表层或上层分布的细根以吸收养分和水分为主,而下层细根的主要功能是吸收深层土壤水。本研究发现,0~20 cm与20~40 cm土壤含水量差异很大,这可能是由于沙地结构松散,土壤毛细管吸收作用力弱,地表存在干沙层对沙地的物理蒸发具有一定的阻碍作用[11]。还有可能是由于紫穗槐根系大量向地表集聚,造成根系分布范围内土壤水分的大量消耗。
根系是植物与土壤环境之间进行物质和能量交换的主要桥梁[23],根系的分布情况反映了植物对环境的适应情况和竞争能力,决定了获取资源的多寡[24]。本研究中根系生物量随着深度的加深而逐渐减小,可能是由于随着土层深度的增加,土壤养分、水分含量下降,土壤温度降低造成的[25],而土壤中不同深度的生物量,可以反映紫穗槐在某一土层深度的生长能力[19]。60龄紫穗槐在0~20 cm土层积累的生物量越多,说明在该层中利用土壤养分、水分和微量元素的能力越强。Jackson等[21]认为,土壤空间异质性是导致根系分布空间异质性的主要原因,根系对土壤空间异质性的基本反应是调整生物量和根长密度。本研究发现,不同株龄紫穗槐根系体积、表面积最大值均出现在0~20 cm,同时根系表现出明显的浅层化,而起支撑作用和构架作用的粗根,表现出明显的分层现象,这也是紫穗槐适应沙漠恶劣环境的策略。不同株龄紫穗槐根系体积、表面积、生物量都以指数形式递减,Coile[26]研究证实,达到一定株龄后根系的垂直分布模式已建立,而根系分布特征受遗传和环境因子双重控制,不同株龄紫穗槐根系分布特征的差异就表现在根系消弱系数的不同。根据Gale对根系在土壤剖面的分布公式Y=l-βh说明根系的垂直分布特征与深度的关系。β值越大说明根系在深层土壤中分布的百分比越大,反之β值越小说明有更多的根系集中分布于接近地表的土层中。β值的大小与根系体积或者根系密度无关[12],从β值分布规律可以看出,紫穗槐根系呈现明显分层现象,0~40 cm以细根为主浅层分布; 40~80 cm以下以骨干根为主深层分布,这样的分布特征可以保证紫穗槐根系在浅层吸收养分和水分,在深层土壤中起支撑和构架作用。根系分布各参数之间表现不同程度的相关性,这与周梦华等[19]在研究云雾山本氏针茅(Stipacapillata)群落根系分布特征的结果基本相符。紫穗槐长期适应于气候波动、高温和干旱的环境中,根系作为其与土壤接触最为紧密的器官,根系分布也会影响根系对营养元素的吸收和抗侵蚀能力。Lawlor[27]研究发现,在供氮不足时通过增加根系体积和密度可以提高氮的吸收,Mamo和Bubenzene[28]研究结果证实:根表面积,根长密度与抗侵蚀能力成正比。根系分布是由遗传因素和环境条件共同控制的一个复杂性状,没有哪一个指标可以确定根系分布特征。所以从不同株龄紫穗槐16个参数(株高、枝条数、枝条直径、地上生物量、枝条长度、根冠比、茎长/根长、土壤水分、根系体积、根系直径、根系表面积、根比例、根系总长、比根长、根长密度、根系生物量)中选取对根系分布影响较强的因子,以发现哪几个参数的作用更为突出。y1中地上生物量、土壤含水量、根系生物量特征向量绝对值系数比较大,可以认为第一主成分为贮能控制因子的综合,属于水分控制因子。y2中根长密度、比根长、根表面积特征向量绝对值系数比较大,可以认为第二主成分为机械支撑和构架作用控制因子的综合,属于构架因子。水分控制因子与构架因子的关系:增加水分和养分的吸收、储能的同时,形成比较发达的机械组织,保持根系分布形态,从而又提高了吸水分和养分的效率。
通过对不同株龄紫穗槐根系分布特征研究,我们发现紫穗槐根系自疏与更新有明显的周期性。我们可以模拟植被根系在整个生活史中的空间分布:幼年期垂直根优先生长,水平根迅速向外伸展,水平根系表现出较强的分枝能力,也较易从水平根上诱发吸收根,垂直根系自然分枝能力较低。进入成年期,垂直根的分生能力进一步下降,而水平根仍然具有不断分枝的能力。到衰老期,垂直根和水平根差别更为明显,垂直根先衰老死亡,形成腐殖质,水平根继续生长,分枝能力增强。紫穗槐根系表现出这种现象,主要是由环境变化的周期性决定的,其中土壤水分、土壤养分对根系的影响很大。另一方面,根系自疏与更新可能与刈割有一定的关系,当地居民为了饲料和碳薪的需要,每年最少刈割2次,这样会刺激地上部分新枝条发生,而新枝条的发生又会刺激新生根产生,这也可能是紫穗槐根系不断更新的原因之一。然而对于土壤养分、水分空间异质性对紫穗槐根系分布特征的响应机制,紫穗槐根系自疏与更新周期所需时间,影响紫穗槐根系自疏与更新的因素等问题还需借助地统计学、分子生物学、根系解剖生理学等方法进行深入研究。
4 结论
1)随着株龄的增加紫穗槐没有出现衰退的现象,其根系有自疏与更新的能力,会不断适应环境的变化,同时紫穗槐地上部形态特征在一定程度上可以反映根系的分布特征。
2)随着紫穗槐株龄的增加,紫穗槐根系贮水能力有所提高,自身根系分布特征决定利用水分的策略。
3)紫穗槐根系呈现明显分层现象,0~40 cm以细根为主,浅层分布,40~80 cm以下以骨干根为主,深层分布,这样的分布特征可以保证紫穗槐根系在浅层吸收养分和水分,在深层土壤中起支撑和构架作用。紫穗槐根系分布在水分因子和构架因子共同影响下,具有自疏与更新能力。紫穗槐根系这种特性,可以达到生态与经济互利共赢的模式,为沙漠周边的居民带来生态环境与经济上的改善。
