庐山若干常绿树种对冬季低温的生理生化响应
2014-10-22杜有新何春林张乐华黄强黄勇丁园
杜有新 ,何春林,张乐华,黄强,黄勇,丁园
1.江西省、中国科学院庐山植物园,江西 庐山 223900;2.南昌航空大学环境与化学工程学院,江西 南昌 330063
植物在越冬期间,体内发生着一系列生理生化过程,合成抵御低温环境的生物酶和相应的生化产物(邵文鹏,2009)。在低温逆境下,植物可通过提高可溶性糖含量来调节细胞液的浓度,从而增强细胞的保水能力(刘金文等,2004);通过合成超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等一系列抗氧化酶,以清除活性氧自由基(ROS),减少低温对植物的危害,如 SOD的作用是通过歧化反应清除生物细胞中的超氧阴离子自由基(O2-·),POD 和 CAT 的主要作用是酶促降解H2O2,减少自由基对机体的伤害(SILIN等, 1993;ISHIKAWA 等, 1995)。丙二醛(MDA)是膜脂过氧化作用的产物,可反映植物细胞膜脂的受损程度(庄炳莉,2008;陈钰等,2007)。
冬季低温是本地区重要的环境胁迫因子之一,由于气温低且持续时间较长,对引进植物的保存及其生长发育起着决定性作用(沈漫等,1997)。虽然自上世纪 70年代以来,许多学者在庐山自然动植物资源、植被分布、土壤等方面展开了积极的研究,但在环境对植物生长发育的影响及植物的响应方面的研究很少(胡亮和王婷,2011),尤其缺乏外来物种对本地区低温环境适应性的机理探讨。本研究通过探讨不同植物对庐山低温环境的响应机制,评价各树种对庐山环境的适应性,为本地区植物引种栽培的树种选择及栽培管理等措施提供科学依据。
1 研究地概况
庐山位于江西北部(115°52′~116°8′ E,29°26′~29°41′ N),虽处亚热带东部季风区域,但山体上部具温带气候特征,年均气温 11.4 ℃,最高气温32 ℃,极端低温达-16.8 ℃。庐山降水充沛,全年平均降水量1917.8 mm,比同纬度地区多500 mm左右,降水集中在4—7月份,约占全年降水量的70%,年平均相对湿度78%(杜有新等,2013)。庐山从12月份到次年的3月份属于寒冷的冬季,根据植物园室外气温的实测数据,2012年12月上旬平均气温为(2.95±1.72)℃,变异系数为58.05%,2013年 1月上旬为(-3.09±2.09) ℃,变异系数为67.73%,3月上旬均温为(4.21±7.92)℃,变异系数为188.36%。
庐山植物园是我国亚热带地区植物种质资源保存的重要基地之一,位于庐山东南部含鄱口沟谷,地形比较复杂,占地面积近300 hm2,海拔为1000~1360 m。土壤为山地黄壤和黄棕壤,有机质含量较高,偏酸性。
2 材料与方法
2.1 研究对象
以庐山植物园内引种栽培的乐东拟单性木兰(Parakmeria Latungensis)、红花木莲(Manglietia insignis)、桂南木莲(Manglietia chingii)、四川含笑(Michelia szechuanica)、深山含笑(Michelia maudiae)和广玉兰(Magnolia grandiflora)等 6种木兰科常绿树种为研究对象,庐山乡土树种云锦杜鹃(Rhododendron fortunei)为对照(表1)。生长表现较好的有深山含笑和红花木莲,桂南木莲、四川含笑和广玉兰冬季冻害较严重,特别是四川含笑在原产地是常绿乔木,引种庐山上后成为落叶灌木。
2.2 采样与分析
在2012年12月初至2013年3月底,根据气温变化,每隔1个月左右分别采集目标树种(红花木莲、深山含笑、广玉兰、乐东拟单性木兰、桂南木莲、四川含笑和云锦杜鹃)冠层新鲜叶片约20 g,每个树种采3株,所采叶样装入保温箱并当日带回实验室处理分析。
生化分析参照《植物生理生化实验原理和技术》(李合生,2000)。可溶性糖含量采用蒽酮比色法测定;丙二醛含量采用硫代巴比妥酸(TBA)法测定;超氧化物歧化酶活性采用氮蓝四唑(NBT)法测定。
2.3 数据处理
采用单因素方差分析(ANOVA)和多重比较判别差异水平,方差分析后采用LSD标准进行显著性检验,数据处理、统计分析及制图在Excel 2003和SPSS 17.0软件上完成。
3 结果与分析
3.1 主要生理生化指标的变化特征
可溶性糖作为植物细胞渗透性的调节物质,其含量与植物的抗寒性密切相关(ISHIKAWA等,1995)。由图1a)可以看出,在冬季期间,可溶性糖含量最高的为四川含笑在1月份的90.38 mg·g-1,最低为红花木莲在12月的含量51.01 mg·g-1。大部分树种的可溶性糖含量呈现出与气温变化相反的趋势即先上升后下降的趋势,只有深山含笑在1月份,比12月份下降了7.54%,其他树种增加幅度从桂南木莲的11.56%到四川含笑的 34.49%之间;深山含笑和广玉兰的可溶性糖含量呈现不断上升趋势,在3月随着气温升高,深山含笑和广玉兰比1月份分别增加12.96%和25.04%。除了桂南木莲和广玉兰外,可反映植物耐寒程度的丙二醛含量呈现不断上升的趋势(图1b)),从12月到1月,上升幅度较大的有红花木莲、深山含笑和四川含笑,而从1月份到3月份,增加幅度较大的为广玉兰、桂南木莲、深山含笑和云锦杜鹃等树种,表明1月低温对这些树种叶片的细胞膜伤害较大。
超氧化物酶(SOD)和过氧化物酶(POD)是植物体内膜系统的重要保护酶,能有效清除超氧自由基和活性氧,降低不良环境对膜脂的伤害程度(庄炳莉,2008)。除了乐东拟单性木兰和深山含笑呈逐渐增加的趋势外,大部分树种的 SOD活性都呈现与气温相似的变化趋势(即先下降后上升的趋势)(图1c));广玉兰的POD活性先上升后下降,而其他树种POD活性则不断上升,但不同树种POD活性大小及变化幅度不尽相同,POD活性最高且变异性最大的是乐东拟单性木兰(图1d))。
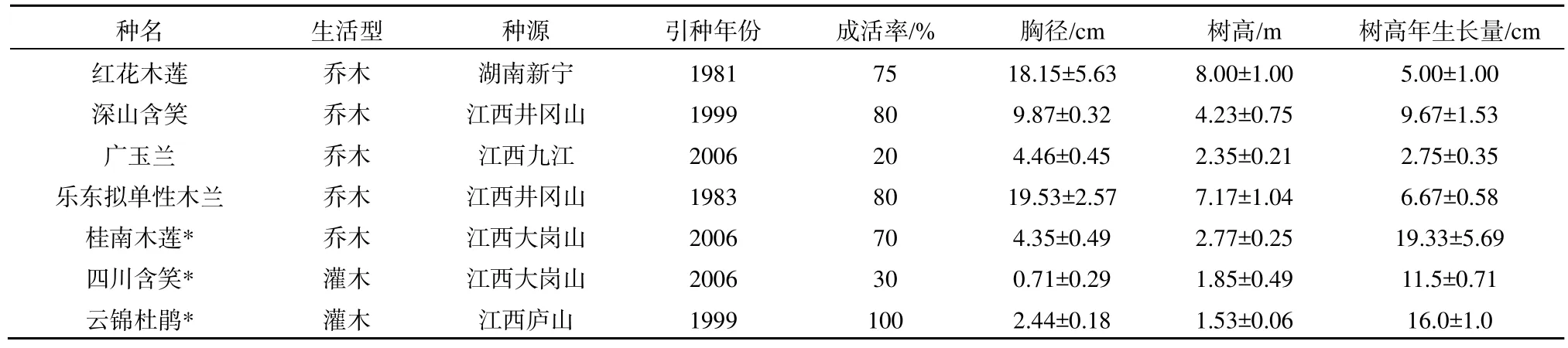
表1 7种树种的生长状况Table 1 Growth condition of 7 tree species in Lushan botanical garden
3.2 主要生化指标的种间差异及其关联特征
通过对不同温度下各树种生化指标比较,发现各树种不同温度间以及各温度下不同树种间的可溶性糖含量存在极显著差异;各树种不同温度间的MDA和SOD存在极显著差异,但种间差异不显著;各树种不同温度间的 POD活性存在显著差异,而各温度下不同树种间存在极显著差异(表2)。
各生化指标的关联特征表明,乐东拟单性木兰(P=0.028)、深山含笑(P=0.047)和广玉兰(P=0.006)冬季叶片中的丙二醛含量和超氧化物歧化酶活性之间具有显著正相关性关系,红花木莲具有显著负相关关系((P=0.047),而对照种云锦杜鹃相关性不显著(P=0.082),但其可溶性糖与超氧化歧化酶显著负相关(P=0.01)。桂南木兰(P=0.009)和四川含笑(P=0.013)的丙二醛与过氧化物酶具有显著正相关关系。说明不同树种在低温胁迫下,做出不同的生理生化响应机制,对低温导致的损害采取不同的修复策略。
4 讨论
植物的生理生化指标可以表征植物经受低温胁迫的生理反应,低温胁迫下植物可溶性糖含量与其抗寒性密切相关。尽管少数品种在低温胁迫下的可溶性糖含量出现下降(龚鹏等,2007;郑元等,2008),但大部分树种通过增加可溶性糖含量来保护细胞,防止细胞过度脱水而受到伤害。本研究表明,大部分树种的可溶性糖含量在初冬时较低,随着气温下降其可溶性糖含量有不同程度升高(GUY等,1992;鲍思伟,2005;蔡仕珍等,2010)。野外调查发现,可溶性糖含量较高的四川含笑当年生的枝叶基本受冻枯死,这说明植物虽然可通过其自身的生理调节来减轻低温的伤害,但仍克服不了其受生物学特性控制的极限低温的伤害;而广玉兰的冻死现象也许说明庐山冬季气温过早下降导致其抵御低温伤害的生理调节机制滞后,由于其生长季节较短(6月份才萌发)导致其木质化程度不高。总体来说,大部分树种在越冬期间可溶性糖含量变化均较明显,通过增加可溶性糖含量的方式来适应冬季低温,抗寒性较强的品种,可溶性糖含量增加幅度大。
丙二醛(MDA)是膜脂过氧化的产物,可反映植物受伤害的程度,丙二醛含量越低则抗寒性越强,含量越高则抗寒性越弱。很多研究表明(庄炳莉,2008;陈钰等,2007),在自然降温过程中,丙二醛在叶片中出现积累,在气温相对最低的1月份达到最大值。本研究表明,大部分树种的 MDA含量随着气温的下降都有不同程度的增加,但3月气温适度回升后,MDA含量都没有降低反而上升(图1b),可能是膜脂过氧化物在深冬时主要分解中间产物自由基,在3月份温度有所回升且温度变化幅度较大时,才分解成最终产物丙二醛,继续对植物叶片造成伤害(滕中华等,2003)。随着温度不断下降,供试树种的丙二醛含量呈现不断上升的趋势,上升幅度较大的有广玉兰、桂南木莲、四川含笑和深山含笑;尤其是四川含笑的MDA含量都比其他树种高,表明 12月底的低温对这些树种叶片的细胞膜伤害较大(罗军武等,2001;何开跃等,2004)。
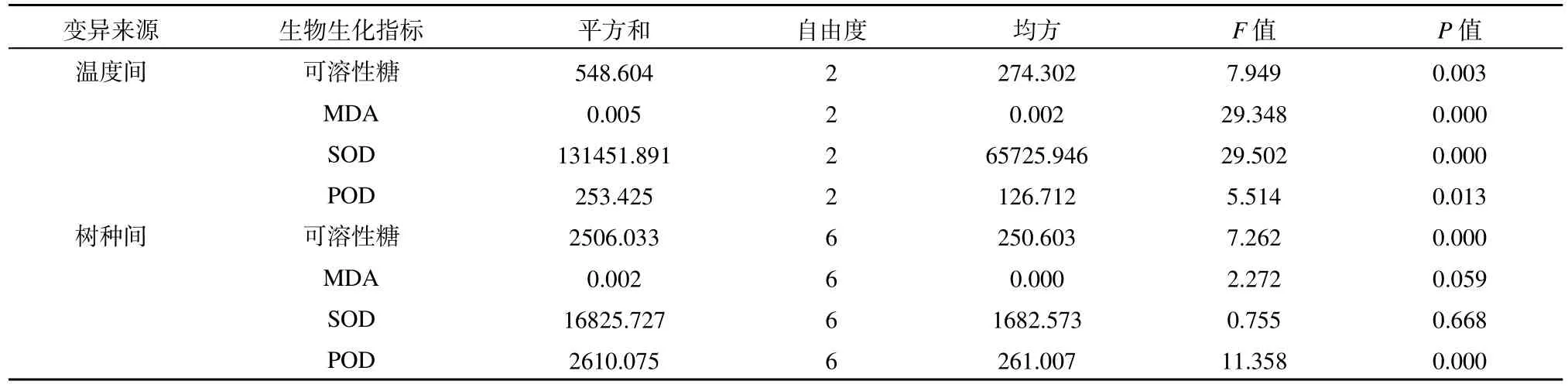
表2 不同温度下各树种可溶性糖、丙二醛、超氧化物歧化酶与过氧化物酶方差分析Table 2 ANOVA of the soluble sugar content, MDA, SOD and POD in leaves among different tree species under different temperatures
在细胞保护酶系统中,SOD的作用是通过歧化反应清除生物细胞中的超氧阴离子自由基(O2-·),同时产生歧化物H2O2和O2,防止过剩自由基对细胞膜的伤害。保护酶活性和植物的抗寒性强弱密切相关,大部分研究认为越冬期间植物的SOD和POD活性先升高后下降(卢芳等,2011;崔帅等,2012;JANDA等,2005;杨向娜等,2006;郭惠红,2004),抗寒能力强的树种 SOD和 POD等保护酶活性高(罗军武等,2001;李建设等,2003)。本研究发现,各树种的SOD活性在越冬期间先下降后上升,7个树种的SOD活性在最低温度的1月份均达到最低值,即随着温度降低,其 SOD活性也降低,当低温有所回升,各树种SOD活性又升高,结合与MDA的变化特征,SOD活性的增加是也许是为了清除植物体内产生的有毒物质。各树种 POD活性分别呈现不断上升或先上升后下降的趋势,不同树种POD活性大小及变化幅度不尽相同,如云锦杜鹃的POD极低,广玉兰的 POD活性先上升后下降,而其他的树种 POD活性则不断上升,这也许说明不同植物在低温胁迫下其保护酶活性的变化趋势不同(朱惠霞等,2007;殷有等,2001)。
5 结论
植物在低温胁迫下,会通过调整组织内部的某些生理生化过程,来规避或减轻低温的伤害。不同树种采用不同的生理响应机制,如乡土树种云锦杜鹃采用提高可溶性糖含量及超氧化物歧化酶活性,其他树种通过提高超氧化物歧化酶或过氧化物酶活性来适应低温环境。4个表征植物耐寒性的生化指标分析表明,可溶性糖含量可以较准确地反映植物随着气温变化所发生的生理生化变化,可作为引种植物抗寒性的主要生理指标之一,而MDA、SOD和POD则存在树种差异。
GUY C L, HUBER J L A, HUBER S C.1992.Sucrose phosphate synthase and sucrose accumulation at low temperature[J].Plant Physiology, 100:502-508.
ISHIKAWA W, ROBERTSON A J, GUSTA L V.1995.Comparison of viability tests for assessing cross- adaptation to freezing, heat and salt stresses induced by abscisic acid in bromegrass suspension cultured cells[J].Plant Science, 107(1): 83-93
JANDA T, KOSA E, SZALAI G, et a1.2005.Investigation of antioxidant activity in maize during low temperature stress[J].Acta Biologica Szegediensis, 49(1/2): 53-54.
SILIN S N, GUY R D, LAVENDER D P.1993.Mefluidide-induced drought resistance in seedings of three cenifer species [J].Canadian Journal of Botany, 71(8): 1087-1092.
鲍思伟.2005.自然降温过程中云锦杜鹃抗寒适应性研究[J].福建林业科技, 32(2): 13-15.
蔡仕珍, 潘远智, 陈其兵, 等.2010.低温胁迫对花叶细辛生理生化及生长的影响[J].草叶学报, 19(1): 95-102.
陈钰, 郭爱华, 姚延涛.2007.杏枝芽内MDA含量和电导率值变化与抗寒性关系的研究[J].天津农业科学, 13(4): 4-6.
崔帅, 赵兰勇, 李承水, 等.2012.卫矛属3 种常绿阔叶树木抗寒性研究[J].中国农学通报, 28(7): 17-20.
杜有新, 何春林, 丁园, 等.2013.庐山植物园 11种植物的根际土壤氮磷有效性和酶活性[J].生态环境学报, 222(8): 1297-1302.
龚鹏, 李秀霞, 李增平, 等.2007.低温胁迫对扁桃枝条细胞膜系统和渗透调节物质的影响[J].新疆农业科学, 44(5): 561-566.
郭惠红.2004.金边卫矛(Euonymus radicans‘Emorald & Gold’)低温抗性研究[D].北京林业大学:54-65.
何开跃, 李晓储, 黄利斌, 等.2004.3种含笑耐寒生理机制研究[J].南京林业大学学报:自然科学版, 28(4): 62-64.
胡亮, 王婷.2011.庐山分布的野生重点保护植物及其保护对策[J].安徽农业科学, 39(15): 9172-9173, 9176.
李合生.2000.植物生理生化实验原理和技术[M].北京: 高等教育出版社:164-165,167-169,199-201,206.
李建设, 耿广东, 程智慧.2003.低温胁迫对茄子幼苗抗寒性生理生化指标的影响[J].西北农林科技大学学报: 自然科学版, 31(l): 90-93.
刘金文, 沙伟, 王艳君.2004.低温胁迫对不同物候期细叶杜香的生理影响[J].植物研究, 24(2): 199-200.
卢芳, 朱鸿菊, 秦登.2011.徐州地区 8种常绿阔叶乔木抗寒性研究[J].江苏农业科学, 39(5): 212-216.
罗军武, 唐和平, 黄意欢, 等.2001.茶树不同抗寒性品种间保护酶类活性的差异[J].湖南农业大学学报: 自然科学版, 27(2): 94-96.
邵文鹏.2009.几种常绿阔叶植物抗寒性研究[D].山东农业大学:9-11.
沈漫, 王明庥, 黄敏仁.1997.植物抗旱机理研究进展[J].植物学通报,14(2): 1-8.
滕中华, 韩发, 师生波, 等.2003.青藏高原黑褐苔草的抗寒性物质季节变化动态研究[J].中国草地, 25(4): 36-40.
杨向娜, 魏安智, 杨途熙, 等.2006.仁用杏3个生理指标与抗寒性的关系研究[J].西北林学院学报, 21(3): 30-33.
殷有, 苏宝玲, 周永斌, 等.2001.抗氧化酶促防御系统对白皮松幼苗抗寒性的影响[J].沈阳农业大学学报, 32(4): 278-250.
郑元, 杨途熙, 魏安智, 等.2008.低温胁迫对仁用杏几个抗寒生理指标的影响[J].西北农林科技大学学报: 自然科学版, 36(1): 163-167.
朱惠霞, 孙万仓, 邓斌, 等.2007.白菜型冬油菜品种的抗寒性及其生理生化特性[J].西北农业学报, 16(4): 34-38.
庄炳莉.2008.自然降温过程中5个常绿阔叶树种的耐寒性研究[D].南京林业大学: 3-7.
