结核杆菌热休克蛋白70作为表位肽载体体外诱导HBV特异性免疫应答*
2014-09-26彭明利
罗 莉,何 松,罗 娜,彭明利
(1.重庆市渝北区人民医院消化内科 401120;2.重庆医科大学附属第二医院消化内科 400010;3.重庆医科大学附属第一医院急诊科 400016;4.重庆医科大学病毒性肝炎研究所 400016)
乙型肝炎病毒(HBV)感染是一种世界性的传染病,全球约有3.5亿 HBV慢性携带者,严重危害人类健康[1]。而主要组织相容性复合物MHC-Ⅰ限制的细胞毒性T淋巴细胞(CTL)在预防、控制、清除HBV感染中起着核心作用[2-3]。已有研究运用结核杆菌热休克蛋白70(TB.HSP70)作为表位肽载体制备疫苗免疫治疗 HBV感染[2]。HBV18-27核心肽的CTL表位在大多数HBV病毒株中高度保守,与HLA-A2.1e有高度亲和力,能与几种 HLA-A2的亚型产生交叉反应[4]。已有研究表明,HSP70-HBcAg(18-27)-CTL 融合蛋白和HSP70-HBcAg(18-27)-CTL复合物可有效诱导 HBV 特异性细胞免疫反应,HBcAg(18-27)的增殖活性较弱,它对淋巴细胞的特异性杀伤作用影响不大[2]。因此,本研究借助结核杆菌HSP70作为表位肽载体,设想将 HBcAg HLA-A2限制性CTL表位 HBcAg(18-27)融合到TB.HSP70的C端,并引入Th和B表位,应用Pichia pastoris酵母分泌表达目的蛋白,体外考察疫苗对慢性HBV感染者外周血淋巴细胞和外周血来源的树突状细胞的作用,进行免疫学功能研究,为慢性乙型肝炎(CAH)的治疗开辟新的途径。
1 材料与方法
1.1 材料 Pichia pastoris GS115表达宿主菌、表达质粒pPIC9和pPIC9K、重组人白细胞介素(IL)-2为Invitrogen公司产品;含TB.HSP70基因的表达载体H37Rv、HepG2.2.15细胞为本实验室构建、保存;限制性内切酶(大连宝生物公司);ATP-agarose(美国Sigma公司);辣根酶标记的羊抗鼠IgG抗体(北京康为公司);鼠抗-TB.HSP70单克隆抗体、FITC标记的鼠抗CD1a、CD86、HLA-DR单克隆抗体及同型对照抗体(Sigma公司);FITC标记的鼠抗CD40单克隆抗体(Abcam公司);检测IL-12p70、IL-1β、肿瘤坏死因子-α(TNF-α)和干扰素-γ(IFN-γ)的酶联免疫吸附测定(ELISA)试剂盒(深圳晶美公司);淋巴细胞分离液(天津TBD公司);PCR引物由大连宝生物公司合成。
1.2 方法
1.2.1 酵母表达质粒的构建与鉴定 以结核分枝杆菌H37Rv作为模板,PCR法扩增含TB.HSP70编码序列的DNA片段(1 900bp),在XhoI和EcoRI位点将扩增的TB.HSP 70基因亚克隆到pPIC9表达载体上获得毕赤酵母质粒pPIC9-HSP70。然后将来自pPIC9-HSP70质粒的 HSP70基因亚克隆到pPIC9K载体的SacI和HpaI位点上获得pPIC9K-HSP70 质 粒 (PIC9/P1)[2,5]。 以 pPIC9-HSP70 为 模板,将 HBcAg(18-27)融合到 TB.HSP70的 C端,并引入 Th和B表位,按照上述方法经双酶切后亚克隆到pPIC9K载体上,构建质粒 pPIC9K-HSP70-HBcAg(18-27)(PIC9/P2)、pPIC9K-HSP70-PreS2B(18-24)-PreS2Th(37-53)-HBcAg(18-27)(PIC9/P3)、pPIC9K-HSP70-PreS2B(18-24)-Th(830-843)-HBcAg(18-27)(PIC9/P4)[2]。将上述构建的质粒进行 XhoI和EcoRI双酶切和测序鉴定。
1.2.2 重组蛋白的表达、纯化与鉴定 按照Invitrogen公司的Pichia pastoris expression system表达系统手册,用SacI酶线性化各质粒,电转化宿主菌GS115[2],重组转化子分别标识为GS115/P1、GS115/P2、GS115/P3、GS115/P4,MD平板选择性培养,PCR方法快速筛选每种阳性转化子6~10株。将宿主菌培养在OD600为1.0的BMMY培养基中,用0.5%的甲醇诱导表达3d。将表达上清液用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)进行电泳后,考马斯亮蓝染色观察有无目的条带。然后将蛋白转移到聚偏氟乙烯(PVDF)印迹膜上,用鼠抗-TB.HSP70单克隆抗体和辣根酶标记的羊抗鼠IgG抗体孵育,蛋白免疫印迹法(Western blot)鉴定表达的目的蛋白[2,5]。将表达量高的菌株接种至1L培养基中诱导表达3d,10 000r/min离心30min,收集的上清液经50×103的滤膜超滤浓缩后,去除小分子和盐,其滤液放置ADP-agarose柱中分离纯化,用1mmol/L的 MgATP预先调节至pH 7.0洗脱,然后在洗脱液中加入乙二胺四乙酸(EDTA)至终浓度2 mmol/L,进行磷酸盐透析得到的洗脱液包含纯度超过95%的目的蛋白 HSP70(P1)、HSP70-HBcAg(18-27)(P2)、HSP70-PreS2B(18-24)-PreS2Th(37-53)-HBcAg(18-27)(P3)、HSP70-PreS2B(18-24)-Th(830-843)-HBcAg(18-27)(P4)[2]。
1.2.3 体外考察重组蛋白对慢性乙型肝炎患者外周血来源的树突状细胞生物学活性的影响 按乙型肝炎治疗指南的标准选取慢性乙型肝炎志愿者(HLA-A2)24例,每组6例,与健康志愿者(CTR)作对照,取20mL外周血,肝素抗凝,常规分离外周血单个核细胞,放置37℃,5%CO2培养箱中用RMIP1640完全培养基(含10%胎牛血清)贴壁培养2~4h。用RMIP1640培养基清洗细胞3次,洗脱未贴壁的细胞(混合淋巴细胞),得到的贴壁细胞在 GM-CSF(1 000U/mL)和IL-4(500U/mL)的作用下诱导成树突状细胞[2]。用RMIP1640培养基重悬树突状细胞,调节细胞浓度为1×106个/mL置于96孔培养板中培养至5d时,分别加入100μg/mL重组蛋白P1、P2、P3各0.1mL(终浓度为10μg/mL),共同孵育2d,以培养基为空白对照,行流式细胞术评估树突状细胞成熟度,包括检测 CD1a、CD40、CD86、HLA-DR表达[6]。重组蛋白负载树突状细胞2d后,ELISA法检测细胞因子分泌情况IL-12p70、IL-1β、TNF-α(pg/mL)。
1.2.4 体外考察重组蛋白诱导淋巴细胞增殖 树突状细胞按照上述方法分离培养5d后,用RMIP1640培养基重悬树突状细胞,调节细胞浓度为1×106个/mL置于96孔培养板中,每孔100.0μL,分别加入100.0μg/mL 重组蛋白P1、P2、P3各0.1mL(终浓度为10.0μg/mL),共同孵育5d。在树突状细胞培养过程中,贴壁孵育后,取未贴壁的细胞(自体淋巴细胞),用RMIP1640完全培养基,含10%的胎牛血清和100U重组人IL-2培养,以2×106个/mL的细胞浓度置于24孔培养板中,分别加入10μg P1、P2、P3刺激,共同培养5d,收集细胞并用无血清1640培养基洗涤2次。负载重组蛋白的树突状细胞以1∶10的比例与洗涤后的自体淋巴细胞混合置于37℃,5%CO2培养箱中共同孵育3d,以培养基为空白对照,每孔加入TdR-3H1μCi,18h后收集细胞,检测cmp值[6]。在细胞共同孵育3d后收集细胞培养上清液,用ELISA检测IFN-γ分泌。
1.2.5 体外考察重组蛋白诱导HBV特异性细胞毒活性 将树突状细胞与10μg重组蛋白P1、P2、P3共同培养5d后,再以1∶10比例与自体淋巴细胞共同孵育7d作为效应细胞,换液,用前24h重组蛋白P1、P2、P3再刺激1次。以HepG2.2.15细胞(表达HBsAg和HBeAg)为靶细胞,将2×106个靶细胞在含20%胎牛血清的1640培养基中用100μCi Na51Cr O4于37℃标记1h,再用无血清的1640培养基洗涤细胞。在1×104个/孔的靶细胞中按不同的效靶比(10∶1、20∶1、40∶1、80∶1)加入效应细胞于37℃10%胎牛血清中孵育4h,取100 μL上清液应用经典51Cr法检测细胞毒活性。杀伤率=100×(实验组释放-自然释放)/[最大释放(0.1mmol HCl)-自然释放]。在所有实验中自然释放小于15%最大释放[6]。
1.3 统计学处理 采用SPSS19.0统计软件进行分析,每次独立的实验重复操作3次,计量资料以x±s表示,采用Student′s t检验,P<0.05表示差异有统计学意义。
2 结 果
2.1 酵母表达质粒的构建与鉴定 构建的质粒经XhoI和EcoRI双酶切后,得到相应的目的片段(图1),同时测序结果也显示表达载体构建成功,表位序列准确地融合在HSP70的C端,质粒分别标识为PIC9/P1、PIC9/P2、PIC9/P3、PIC9/P4。
2.2 重组蛋白的表达、纯化与鉴定 实验表明重组蛋白P1、P2、P3能够顺利地分泌表达(在70×103处均有一条带),而带有破伤风毒素通用辅助性T细胞识别表位Th830-843融合蛋白P4不能分泌表达(图2A),经实验反复证实,但胞内检测有表达,可能Th830-843表位的引入干扰了该载体分泌信号的正确切割。将表达量高的菌株接种至1L培养基中诱导表达3 d,离心,上清液经50×103的超滤浓缩后,用ADP-agarose介质亲和层析得纯度超过95%的目的蛋白,摇瓶实验显示目的蛋白表达量大于120mg/L(图2B)。

图1 表达质粒的酶切鉴定
2.3 重组蛋白刺激慢性乙型肝炎与健康者外周血来源树突状细胞表面分子表达 体外重组蛋白诱导树突状细胞成熟,促进细胞因子分泌,树突状细胞在培养5d时,分别加入P1、P2、P3,浓度为10μg/mL,共同孵育2d,以培养基为对照,行流式细胞术检测树突状细胞CD1a、CD40、CD86、HLA-DR表达,结果见表1。树突状细胞经 P1、P2、P3刺激后,CD1a、CD40、CD86均明显提高,CD1a乙型肝炎患者较健康者低,CD40、CD86二者差异无统计学意义(P>0.05),HLA-DR差异无统计学意义(P>0.05),且 HLA-DR的表达在抗原刺激前后差异无统计学意义(P>0.05),重组蛋白负载树突状细胞2d后,ELISA法检测细胞因子分泌情况(图3),P1、P2、P3均能诱导树突状细胞分泌 Th1型细胞因子(IL-12p70、IL-1β、TNF-α),有利于Th0向Th1的极化。

表1 重组蛋白刺激慢性乙肝与健康者外周血来源树突状细胞表面分子表达(%pos)
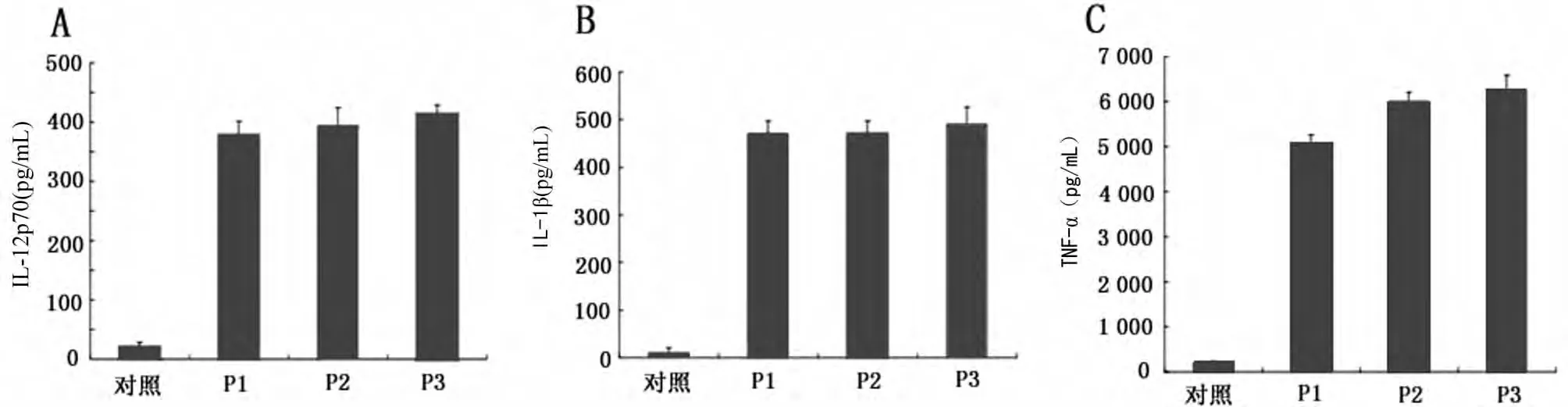
图3 重组蛋白刺激慢性乙型肝炎外周血来源树突状细胞因子分泌情况

图4 重组蛋白负载树突状细胞刺激淋巴细胞增殖和IFN-γ的分泌
2.4 体外重组蛋白诱导淋巴细胞增殖和IFN-γ释放 将负载重组蛋白的树突状细胞按1∶10与自体淋巴细胞孵育3d,TdR-3H掺入法检测淋巴细胞的增殖情况(图4A)和ELISA检测IFN-γ分泌(图4B),结果显示P1、P2、P3明显刺激淋巴细胞增殖和释放IFN-γ。
2.5 体外重组蛋白诱导HBV特异性细胞毒活性 以HepG2.2.15细胞(表达HBsAg和HBeAg)为靶细胞,在体外应用经典51Cr法检测重组蛋白(P1、P2、P3)诱导 HBV特异性细胞毒活性(图5),其结果显示P3能引起较强的细胞毒活性,健康者较乙型肝炎患者强,在效靶比为80∶1时,前者为65%,后者为44%,这可能与乙型肝炎患者T细胞功能缺陷有关,而单纯的TB.HSP70(P1)基本没有细胞毒性。以HepG2和HEN细胞为靶细胞,P3几乎无细胞毒性。
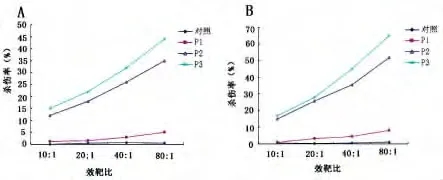
图5 重组蛋白诱导自身淋巴细胞HBV特异性细胞毒活性
3 讨 论
本研究借助TB.HSP70作为表位肽载体,选用了应用最多的 HBcAg HLA-A2限制性 CTL表位 HBcAg(18-27),将HBcAg(18-27)融合到TB.HSP70的C端,并引入Th和B表位,应用Pichia pastoris酵母分泌表达目的蛋白HSP70(P1)、HSP70-HBcAg(18-27)(P2)、HSP70-PreS2B(18-24)-PreS2Th(37-53)-HBcAg(18-27)(P3),体外考察这些疫苗对慢性 HBV感染者外周血淋巴细胞和外周血来源的树突状细胞的作用,表明HSP70可作为HBV HBcAg细胞毒性CTL表位肽载体,提高CTL表位肽的免疫原性,特别是含有B、Th表位的P3能更好地激活HBV特异性免疫应答。
作者之前的研究证实HBcAg(18-27)有较弱的增殖活性,它对淋巴细胞的特异性杀伤作用影响不大[2,7]。而本研究观察了重组蛋白(P1、P2、P3)在体外诱导树突状细胞成熟、淋巴细胞增殖以及HBV特异性细胞毒活性情况。其结果显示P1、P2、P3能上调CD1a、CD40、CD86表达,诱导未成熟的树突状细胞分泌大量的IL-12p70、IL-1β、TNF-α,有利于 Th0向 Th1的极化,促进树突状细胞成熟,增强树突状细胞抗原递呈能力,同时促进淋巴细胞增殖和释放IFN-γ,而且P3最能有效激活自体淋巴细胞成为细胞毒活性细胞,而单独的HSP70没有杀伤活性,不能有效引起机体特异性免疫应答。这些实验结果表明HSP70作为表位肽载体大大增强了CTL表位肽的免疫原性,特别是引入B、Th表位,可更有效地诱导HBV特异性免疫应答。其可能的原因为:(1)HSP70(P1)和融合蛋白(P2、P3)结合树突状细胞能上调CD1a、CD40、CD86表达,诱导树突状细胞分泌产生高水平的 Th1 型细胞因子(IL-12p70、IL-1β、TNF-α),其中IL-12在诱导Th1极化中起关键作用,另外诱导释放的IFN-γ也有利于Th0向Th1的极化,从而促进树突状细胞成 熟,增 强 其 免 疫 活 性[2,8-9]。(2)HSP70特 异 性 结 合CD40,作用于单核细胞、树突状细胞等激活免疫系统,促进树突状细胞释放CC趋化因子如CCL3(MIP-1a)、CCL4(RANTES)。通过CD40信号可以上调抗原呈递能力,增加共刺激分子CD80、CD86和CD58的表达,使树突状细胞能向CTL启动[10]。因此树突状细胞通过刺激T细胞、促进CD40表达,以及与CD40L的交联作用诱导 MHC-I限制性抗原特异性CD8+T细胞反应[3,11]。
本研究通过体外考察表明TB.HSP70可作为HBV HB-cAg细胞毒性CTL表位肽载体,提高CTL表位肽的免疫原性,而且含有B和Th表位的P3能更好地激活HBV特异性免疫应答,这为CAH的免疫治疗开辟新的途径。Floto等[12]研究表明TB.HSP70通过CCR5受体作用于树突状细胞,诱导树突状细胞聚集,促进树突状细胞与T细胞形成免疫应答,而P3是否也通过CCR5受体作用于树突状细胞,这将在后续研究中进一步探讨。
[1]Brooks J,Gelson W,Rushbrook SM.Therapeutic advances in the management of chronic hepatitis B infection[J].Ther Adv Chronic Dis,2013,4 (4):157-166.
[2]Peng M,Chen M,Ling N,et al.Novel vaccines for the treatment of chronic HBV infection based on mycobacterial heat shock protein 70[J].Vaccine,2006,24(7):887-896.
[3]Moffat JM,Cheong WS,Villadangos JA,et al.Hepatitis B virus-like particles access major histocompatibility classⅠandⅡantigen presentation pathways in primary dendritic cells[J].Vaccine,2013,31 (18):2310-2316.
[4]del Guercio MF,Sidney J,Hermanson G,et al.Binding of apeptide antigen to multiple HLA alleles allows defnition of an A2-like supertype[J].J Immunol,1995,154(2):685-693.
[5]彭明利,凌宁,许红梅,等.结核杆菌热休克蛋白70在毕赤酵母中的分泌表达与鉴定 [J].生物工程学报,2003,19(3):286-290.
[6]彭明利,陈敏,任红.结核杆菌热休克蛋白质70作为乙型肝炎核心抗原细胞毒性T淋巴细胞表位肽载体的研究[J].中华肝脏病杂志,2006,14(4):406-407.
[7]彭明利,任红,许红梅,等.结核杆菌热休克蛋白70-乙型肝炎表面抗原诱导特异性免疫反应[J].中华肝脏病杂志,2003,11(5):271-274.
[8]Tavakoli S,Schwerin W,Rohwer A,et al.Phenotype and function of monocyte derived dendritic cells in chronic hepatitis B virus infection[J].J Gen Virol,2004,85(Pt 10):2829-2836.
[9]Ge C,Xing Y,Wang Q,et al.Improved efficacy of therapeutic vaccination with dendritic cells pulsed with tumor cell lysate against hepatocellular carcinoma by introduction of 2tandem repeats of microbial HSP70peptide epitope 407-426and OK-432[J].Int Immunopharmacol,2011,11(12):2200-2207.
[10]Caux C,Massacrier C,Vanbervliet B,et al.Activation of human dendritic cells through CD40cross-linking[J].J Exp Med,1994,180 (4):1263-1272.
[11]Borges TJ,Wieten L,van HMJ,et al.The anti-inflammatory mechanisms of Hsp70[J].Front Immunol,2012,3:95.
[12]Floto RA,MacAry PA,Boname JM,et al.Dendritic cell stimulation by mycobacterial Hsp70is mediated through CCR5[J].Science,2006,314(5798):454-458.
