部分野生杜鹃杂交授粉后花粉管生长状况分析
2014-09-21耿兴敏张超仪尹增芳
耿兴敏,张超仪,尹增芳
(1.南京林业大学 风景园林系,江苏南京 210037;2.南京林业大学森林资源与环境学院,江苏 南京 210037)
我国野生杜鹃花资源丰富,占世界杜鹃花资源的半数以上[1],但我国对于杜鹃花资源的应用与研究,尤其是新品种选育方面,还相当薄弱,仅限于张长芹[2]、刘晓青[3]等的报道.国外有关杜鹃花杂交育种的研究起步较早,杂交育种相关的研究也有大量报道,到2002年为止,登记的杜鹃花新品种已经有28 000种[4].杜鹃花种间杂交障碍存在着受精前与受精后障碍.杜鹃种间杂交授粉后,花粉基本在柱头都可以萌发,但柱头表面可能会产生胼胝质阻止花粉管进入花柱,在进入花柱后,花粉管也可能发生顶端膨大、变形、停止生长等现象,或花粉管无法进入胚珠以及花粉管在胚囊过度生长等[5-7].花粉管的生长受阻可能发生在不同的部分,不同杂交组合中花粉管生长受阻的部位存在种间差异[6].有报道称蒙导授粉和正己烷处理可以克服杜鹃受精前障碍[8].受精后障碍主要表现为胚珠停止发育,种子不萌发或萌发率很低以及后代出现白化苗等现象[9-11].胚拯救是克服受精后障碍的有效技术手段,在杜鹃育种中也有报道[12].在常绿杜鹃与黄花落叶杜鹃杂交过程中,常出现白化苗现象,Ureshino等[13-14]利用三重杂交、回交和倍性杂交等有效克服了受精后障碍并获得绿色幼苗.
近年来,各地纷纷展开了野生杜鹃的引种驯化工作,但由于野生杜鹃大多分布在高山地区,生长发育要求冷凉的环境条件,对高温的忍耐力不强,杜鹃属植物,尤其是常绿品种的引种栽培至今没有成功[15-16].高温胁迫严重影响了杜鹃花在我国城市中的推广应用.
因此该研究以培育耐热性强、生态适宜范围广的杜鹃新品种为目的,以南京地区栽培的耐热性较好的映山红(Rhododendron simsii)和锦绣杜鹃(Rhododendron pulchrum)为母本,自交的同时,与高海拔地区引种的杜鹃属其他各种进行杂交.利用荧光显微镜观察这些杂交组合授粉后花粉管的生长状况,分析其远缘杂交亲和性的同时,进一步明确杜鹃杂交存在的种间杂交受精前、后障碍,为我国野生杜鹃的杂交育种提供理论依据.
1 材料与方法
1.1 实验材料
以南京地区栽培的映山红(R.simsii)、锦绣杜鹃(R.pulchrum)为母本,自交的同时,以表1所列野生杜鹃种为父本(花粉来自中科院昆明植物园),配置杂交组合.

表1 各杜鹃野生种的学名、分类及编号Tab.1 Latin name and classification of wild Rhododendron and their abbreviations
1.2 试验方法
2011年4月中旬开花前3 d对母本进行去雄套袋,待柱头分泌大量黏液时选择于当天上午9:00—12:00进行常规人工授粉,各组合至少重复15朵花.授粉24、48、72 h后采集花柱及子房,每个杂交组合取5朵花.参照Ureshino等[5]的方法,将柱头等用FAA液固定24 h后,置于70%的酒精内,于4℃条件下保存,用于解离、染色和观察.用3 mol·L-1的NaOH溶液45℃水浴解离3 h,解离前后都要用蒸馏水冲洗,然后用0.1%的苯胺蓝(0.1 mol·L-1K3PO4·7H2O缓冲液)染色3 h,压片后使用Leica DM500B荧光显微镜观察、拍照.统计花粉管生长数量及其异常花粉管的比率.
2 结果与分析
2.1 不同杂交组合花粉管异常生长状况分析
杂交授粉后花粉管生长状况如图1所示.图1中:(1)映山红×白花杜鹃,花粉管在柱头萌发,并向下开始生长(箭头所示);(2)映山红×白花杜鹃,花粉管在柱头上部生长,部分花粉管出现膨大、扭曲生长(箭头所示);(3)映山红×白花杜鹃,花粉管在柱头下部生长,胼胝质规则分布(箭头所示);(4)映山红×白花杜鹃,杂交授粉3 d后花粉管进入胚囊;(5)锦绣杜鹃×薄皮杜鹃,授粉5 d后,大多花粉管在柱头缠绕(箭头所示),没有进入花柱道,花粉管有胼胝质不规则沉积(箭头所示);(6)映山红×薄皮杜鹃,花粉管粗细不均(箭头所示);(7)锦绣杜鹃×滇南杜鹃,花粉管在柱头表面打结(箭头所示);(8)映山红×滇南杜鹃,花粉管在花柱内螺旋、打结(箭头所示);(9)映山红×露珠杜鹃,花粉管破裂(箭头所示);(10)锦绣杜鹃×薄皮杜鹃,花粉管螺旋生长、粗细不均(箭头所示);(11)锦绣杜鹃×红露珠杜鹃,先端沉积胼胝质塞(箭头所示);(12)锦绣杜鹃×睫毛杜鹃,花粉管卷曲、杂乱生长(箭头所示);(13)锦绣杜鹃×露珠杜鹃,花粉管尖端膨大成气球状(箭头所示);(14)映山红×滇南杜鹃,杂交后子房未见膨大,但杂交24 h后花粉管就进入胚囊(箭头所示);(15)映山红×美容杜鹃,花粉管在子房内螺旋生长(箭头所示);(16)映山红×映山红(昆明),花粉管正常生长.
总之,该试验所配置的杂交组合中,所有花粉在授粉1 d后都能在柱头萌发,杂交亲和的花粉在花柱上部花粉管也部分出现异常,但大多数花粉仍能继续生长进入胚囊(图1(1)~1(4)).杂交亲和性较差的组合,如图1(5)所示,锦绣杜鹃×薄皮杜鹃在授粉5 d后大多数花粉还在柱头盘旋,没有进入花柱.所有杂交组合花粉管生长状况的观察结果表明,花粉管异常现象主要表现为花粉管粗细不均、螺旋、破损、尖端膨大、杂乱生长、胼胝质沉积等异常情况(图1(6),图1(9)~1(15)),少数组合出现花粉管打结现象(图1(7)和1(8)),如映山红×大字杜鹃、映山红×薄皮杜鹃、锦绣杜鹃×美容杜鹃、锦绣杜鹃×薄皮杜鹃等.还有一些杂交组合,如锦绣杜鹃×薄皮杜鹃、映山红×薄皮杜鹃出现较多的异常类型.
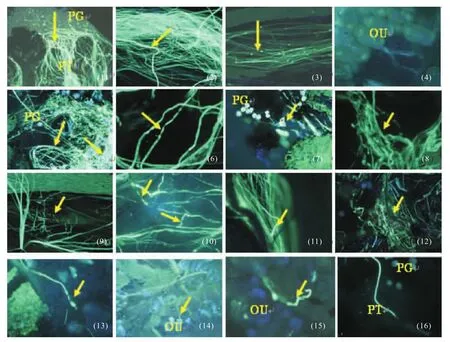
图1 杂交授粉后花粉管生长状况分析Fig.1 Pollen tube growth after pollination
2.2 不同杂交组合花粉管生长状况
2.2.1 映山红杂交组合
如图2所示,以映山红作为母本时,所有杂交组合的花粉在授粉24后,映山红自交、映山红×映山红(昆明)、映山红×薄皮杜鹃以及映山红×滇南杜鹃组合的花粉管已穿过花柱进入子房.其他杂交组合,其父本花粉也都能够在柱头表面萌发,但进入花柱通道内的花粉管数量及异常花粉管的比率,因杂交组合不同而有所不同.进入花柱的花粉管数量大于50根的父本花粉,包括睫毛杜鹃、白花杜鹃、红花露珠杜鹃以及映山红自交花粉,但红花露珠杜鹃以及映山红自交花粉有70%以上出现花粉管异常现象.大白杜鹃、美容杜鹃、马缨杜鹃及大字杜鹃萌发花粉中,进入花柱的花粉管数量较少,不到20根,但异常花粉管比例也有所降低.另外,映山红×露珠杜鹃的杂交花粉,进入花柱的花粉管数量较少,但异常花粉管的比例很高.

图2 授粉后花粉管在映山红柱头中生长状况及异常花粉管比例Fig.2 Pollen tube growth on stigmas and into the style of R.simsii after pollination and frequency of abnormal pollen tubes
授粉72 h后,大部分杂交组合进入花柱中的花粉管数量均多于授粉后24 h,但映山红×马缨杜鹃组合进入花柱的花粉管数量未见增加,仍少于20.并且随着进入花柱花粉管数量的增加,部分杂交组合,如映山红×睫毛杜鹃和映山红×大白杜鹃异常花粉管的比例也在增加.除了映山红×露珠杜鹃、映山红×大字杜鹃、映山红×马缨杜鹃、映山红×白花杜鹃以及映山红×窄叶杜鹃以外,大部分杂交组合在授粉72 h后,花粉管都伸入了子房.
授粉120 h后,各组合花粉管进入花柱的数量也有所增加(>20根),且各进入花柱的花粉管均能到达花柱底部.但个别组合,如映山红×薄皮杜鹃、映山红×滇南杜鹃异常花粉管的比例增高,主要表现为较严重的胼胝质沉积、花粉管破裂等现象.其他组合也存在花粉管异常现象,但比例较低,如映山红×睫毛杜鹃、映山红×露珠杜鹃、映山红×大字杜鹃、映山红×马缨杜鹃、映山红×美容杜鹃.
2.2.2 锦绣杜鹃杂交组合
授粉后花粉管在锦绣杜鹃柱头中生长状况及异常花粉管比例如图3所示.

图3 授粉后花粉管在锦绣杜鹃柱头中生长状况及异常花粉管比例Fig.3 Pollen tube growth on stigmas and into the style of R.pulchrum after pollination and frequency of abnormal pollen tubes
由图3可知,以锦绣杜鹃作为母本时,不同种类父本各花粉虽然亦能在柱头表面萌发,但花粉粒附着数量、萌发数量与花粉管长度等却存在显著差异.授粉24 h后,大部分组合花粉在柱头的附着量都较多,但白花杜鹃、窄叶杜鹃和滇南杜鹃花粉在母本柱头的附着量较少,并且除锦绣杜鹃×滇南杜鹃以外各组合花粉管均能通过柱头进入花柱.有部分组合萌发的花粉管数量在50根以上,如锦绣杜鹃×映山红、锦绣杜鹃×大字杜鹃与锦绣杜鹃×睫毛杜鹃,但花粉管异常的比率高达31% ~100%.与映山红为母本的杂交组合相比,大部分组合花粉管数量都比较少,并且可以发现,除了锦绣杜鹃×薄皮杜鹃、锦绣杜鹃×美容杜鹃、锦绣杜鹃×马银花与锦绣杜鹃×白花杜鹃以外,较多花粉管存在胼胝质过量沉积或者粗细不一现象.同时,仅部分组合的花粉管能够在24 h内达到花柱底部,如锦绣杜鹃×映山红、锦绣杜鹃×大白杜鹃与锦绣杜鹃×白花杜鹃.
与授粉第24 h相比,授粉72 h后,大部分杂交组合中进入花柱的花粉管数量都有所增加,但是穿过花柱进入子房的杂交组合数量并未增多,仅部分组合的花粉管继续向下生长,如锦绣杜鹃×红花露珠杜鹃、锦绣杜鹃×大字杜鹃、锦绣杜鹃×马缨杜鹃、锦绣杜鹃×窄叶杜鹃、锦绣杜鹃×马银花、锦绣杜鹃×滇南杜鹃,其中锦绣杜鹃×红花露珠杜鹃、锦绣杜鹃×大字杜鹃、锦绣杜鹃×马银花与锦绣杜鹃×滇南杜鹃等组合的花粉管的70%都存在异常现象.
授粉后120 h后,包括自交在内的14个杂交组合当中,有8个杂交组合的花粉管进入花柱底部,其中锦绣杜鹃自交、锦绣杜鹃×映山红(昆明)、锦绣杜鹃×白花杜鹃有较多花粉管到达花柱底部(>50根),异常花粉管数量较少(<30%),其他5个组合进入花柱底部的花粉管数量较少,并且锦绣杜鹃×红花露珠杜鹃、锦绣杜鹃×马缨杜鹃及锦绣杜鹃×美容杜鹃3个组合的异常花粉管比例较高.
3 讨论
亲和性授粉在花柱道和胎盘凹槽生长期间,花粉管是直的、末端半透明(称为帽区),在帽区后边相隔一定距离有规则的胼胝质沉积[6].映山红、锦绣杜鹃自交、杂交后,花粉管的生长出现扭曲、螺旋、膨大、断裂和胼胝质过量沉积等异常情况.Williams等[6]在杜鹃远缘杂交时还发现受精后花粉管末端渐缩、有尖刺等其他花粉管生长异常现象.花粉管异常情况出现的类型与杂交亲和性无直接联系,即使自交的组合也发现花粉管螺旋、破损、胼胝质过量沉积等异常的情形,部分组合如:锦绣杜鹃×薄皮杜鹃杂交组合,除以上异常状况外,还发现花粉管打结、杂乱生长、先端膨大等异常,在所有异常情况中较多组合出现花粉管胼胝质过量沉积、螺旋,而花粉管打结现象较少.
根据Williams等[6]在杜鹃花科远缘杂交研究中的发现,花粉管的生长受阻可能发生在7个不同的部分:①花粉在柱头表面无法萌发;②花粉粒萌发,产生花粉管,但花粉管在柱头表面发生异常;③花粉管进入花柱道,但在花柱道内出现异常;④花粉管无法进入子房;⑤花粉管进入子房,但不能进入珠孔;⑥花粉管进入珠孔,但无法进入胚囊;⑦花粉管无法在胚囊内释放精子.不同杂交组合中花粉管生长受阻的部位存在种间差异.
该研究以映山红作为母本杂交,各父本花粉都能够在映山红柱头上附着并萌发,并且各父本花粉管基本上都能到达花柱下部,虽然不同杂交组合花粉管的生长速度、到达花柱基部的比率及异常花粉管的比率等存在显著差异(图2).个别组合,如映山红×马缨花、映山红×滇南杜鹃杂交2个月后子房未见膨大[17],但在研究中也观察到有花粉管进入胚珠(图1(14)).解玮佳等[18]在高山杜鹃与大喇叭杜鹃(R.excellens)杂交时也有类似发现,即有花粉管进入胚囊,但未见子房膨大.花粉管进入胚囊,并不一定能够完成受精作用,可能会出现胚囊不亲和现象,即花粉管进入胚囊并释放出精子,但精子却不能有效地与雌配子结合实现双受精,这在玉米等作物的远缘杂交中也有报道[19].在杜鹃杂交研究中发现,花粉管进入胚囊后,会因出现过度生长等情况,而无法完成受精[7].该研究还发现部分杂交组合在萌发初期,或花柱中、上部出现异常花粉管,但在下部恢复正常,进入胚囊.Williams等[6]发现异常的花粉管也能伸入胚囊,但是否能顺利完成受精,还需要进行进一步胚胎发育学研究.
锦绣杜鹃作为母本杂交时,仅种内及映山红组内杂交72 h后,有花粉管进入花柱下部(图3),这几个杂交组合也获得了种子[17].杜鹃亚属、马银花亚属以及同一亚属不同组的大字杜鹃花粉管没能到达花柱基部,在花柱通道内花粉管的生长受阻,或因其生长速度过慢,在授粉72 h后花粉管仍未能达到花柱底部;并且这些杂交组合中异常花粉管的比率也较高(图3).这些杂交组合在杂交后均未获得杂交种子,虽然部分杂交组合,如锦绣杜鹃×大字杜鹃、锦绣杜鹃×睫毛杜鹃等,杂交2个月后有子房膨大现象,但并未正常发育,形成种子[17],这表明这些杂交组合同时存在受精后障碍.
整体来看,杂交亲和性较好的组合,花粉管生长速度较快,到达花柱底部的花粉管比率较高,如映山红自交及种内杂交.Williams和Rouse[20]的研究表明,花粉管生长速度及进入子房的时间与雌蕊花柱长度有关,父本/母本的柱头长度比(SLR)显著影响种间杂交成功率,比值越接近1,杂交成功率越高.映山红做母本时,花粉管到达花柱底部的杂交组合多于锦绣杜鹃母本,异常花粉管比率低(<30%),这表明作为母本,映山红种间杂交亲和性要优于锦绣杜鹃.这一结果也表明母本的不同可能导致花粉管生长状况不同;同种母本因父本的不同也可能对花粉管的生长情况有不同影响,Ureshino等[5]的研究中也有类似报道.
[1]方瑞征.中国植物志·第57卷[M].北京:科学出版社,1996.
[2]张长芹,冯宝钧,吕元林.杜鹃花属的杂交育种研究[J].云南植物研究,1998,20(1):94-96.
[3]刘晓青,苏家乐,李畅,等.杜鹃花杂交、自交及开放授粉结实性研究[J].上海农业学报,2010,26(4):145-148.
[4]Leslie A C.The international Rhododendron register and checklist[M].2nd ed.U.K.:Royal Botanic Garden Edinburgh,Edinburgh,2002.
[5]Ureshino K,Kawai M,Miyazima I.Factors of intersectional unilateral cross incompatibility between several evergreen azalea species and Rhododendron japonicum f.flavum[J].Japan Soc Hort,2000(36):261 -265.
[6]Williams E G,Knox R B,Rouse J L.Pollination sub -systems distinguished by pollen tube arrest after incompatible interspecies crosses in Rhododendron(Ericaceae)[J].Journal of Cell Science,1982(53):255 - 277.
[7]Williams E G,Kaul V,Rouse J L,et al.Overgrowth of pollen tubes in embryo sacs of Rhododendron following interspecific pollinations[J].Amer J Bot,1986(34):413 - 423.
[8]Lee J S,Ryu B Y.Breaking of interspecific incompatibility in Korea native and introduced azalea[C]∥24th International Horticultural Congress(Abstract),1994:165.
[9]Okamoto A,Suto K.Cross incompatibility between Rhododendron sect.Tsutsusi species and Rhododendron japonicum(A.Gray)J.V.Suringar f.flavum Nakai[J].Journal of the Japanese Society for Horticultural Science,2004,73(5):453-459.
[10]Kita K,Kurashige Y,Yukawa T,et al.Intergeneric hybridization between Menziesia and Rhododendron based on molecular phylogenetic data[J].Journal of the Japanese Society for Horticultural Science,2005a,74(1):51 -56.
[11]Kita K,Kurashige Y,Yukawa T,et al.Plastid inheritance and plastome-genome incompati-bility of intergeneric hybrids between Menziesia and Rhododendron[J].Journal of the Japanese Society for Horticultural Science,2005b,74(1):318-323.
[12]Eeckhaut T,de Keyser E,van Huylenbroeck J,et al.Application of embryo rescue after interspecific crosses in the genus Rhododendron[J].Plant Cell Tissue and Organ Culture,2007,89(1):29 -35.
[13]Ureshino K,Abe T,Akabane M.Relationship between nuclear genome construction and the plastome-genome incompatibility of progenies from intra and inter-ploid cross of evergreen Azaleasx Rhododendron japonicum f.flavum[J].Journal of the Japanese Society for Horticultural Science,2010,79(1):91 -96.
[14]Ureshino K,Miyajima I,Akabane M.Effectiveness of three-way crossing for the breeding of yellow-flowered evergreen azalea[J].Euphytica,1998,140(1):113 -118.
[15]刘晓青,苏家乐,李倩中,等.部分高山杜鹃品种在南京的性状表现及配套栽培技术[J].江苏农业科学,2008(1):119-120.
[16]张春英,戴思兰,张启翔.常绿杜鹃花叶片耐热性指标研究[R].中国园艺学会观赏园艺专业委员会年会:抗性生理,2006:409-412.
[17]耿兴敏,张超仪,罗凤霞,等.中国野生杜鹃花杂交结实性研究[J].江苏农业科学,2013,2:159-161.
[18]解玮佳,李世峰,李树发,等.高山杜鹃与大喇叭杜鹃种间杂交过程的观察研究[J].西北植物学报,2012,32(12):2432-2437.
[19]段桃利.玉米与其近缘种属杂交花粉管行为研究[D].成都:四川农业大学玉米研究所,2008.
[20]Williams E G,Rouse J L.Relationships of pollen size,pistil length and pollen tube growth in Rhododendron and their influence on hybridization[J].Sex Plant Reprod,1990(3):7 -17.
