siRNA干扰碱性成纤维细胞生长因子表达对肺腺癌A549增殖和克隆特性的影响
2014-09-12白小燕孟又胜王秀问王亚伟
白小燕 孟又胜 王秀问 王亚伟
(成都市第五人民医院呼吸内科,四川 成都 611130)
肿瘤干细胞理论认为肿瘤干细胞是肿瘤发生和复发的根源,它们具有无限的自我更新、增殖、多潜能分化及抗拒化放疗等特点,针对肿瘤干细胞的治疗才是根本。碱性成纤维细胞生长因子(bFGF)在人体正常组织和多种肿瘤广泛表达,bFGF信号通路在肿瘤的发生发展及维持干细胞特性方面起重要作用。因此,本研究应用RNA干扰(RNAi)技术抑制bFGF表达,观察bFGF信号通路对A549的增殖和自我更新等特性的影响。
1 材料与方法
1.1细胞株与试剂 A549细胞系购自中国科学院上海细胞生物研究所;RPMI-1640购自Hyclone公司产品;胎牛血清(FBS)购自TBD公司; bFGF抗体购自SANTA CRUZ公司;β-actin及OCT-4抗体系Abcam公司产品;辣根酶标记山羊抗兔IG购自中杉金桥公司;RIPA蛋白裂解液、BCA蛋白定量试剂盒均购自碧云天公司;蛋白Marker购自Fermentas公司;ECL发光液购自北京博奥森公司; FGF2-RNA质粒载体购自上海吉凯基因化学技术有限公司;质粒抽提试剂盒购自北京TIANGEN公司;Nano Fectin转染试剂购自SBI公司;Trizol购自Invitrogen;引物购自上海博尚生物公司。
1.2主要仪器 CO2细胞培养箱(美国Heraus公司)、SWCJ型超净工作台(苏州净化设备公司)、ELx800酶标仪(美国Bio-TEK公司)、倒置显微镜及荧光显微镜(日本Olympus公司)、凝胶分析仪(江苏捷达科技公司)、PTC-200 DNA扩增仪(美国MJ公司)、紫外分光光度计(美国Bekman公司)、低温高速离心机(美国Dupont公司)、台式高速离心机及移液器(德国eppendorf公司)、恒压恒流电泳仪(北京六一仪器厂)。
1.3方法
1.3.1细胞培养 A549细胞置于含10%胎牛血清,青、链霉素各100 U/mL的RPMI-1640培养液中,37 ℃、体积分数为5%的CO2、湿度为100 %的孵箱中培养,每2~3 d传代。
1.3.2bFGF干扰质粒的构建 以bFGF编码基因(GenBank ID:NC_000004)的799~819区域为靶序列,设计siRNA干扰序列: 5′-ACTACAATACTTACCGGTCAA-3′,Loop结构为CTCGAG,命名为pGCsiU6/ puro/GFP-bFGF-siRNA,以无关序列5′-TTCTCCGAACGTGTCACGT-3′作为阴性对照(NC),此序列不与任何人类基因序列同源,命名为pGCsiU6/ puro/GFP-NC-siRNA。质粒构建及测通等具体工作由上海吉凯基因化学技术有限公司完成。
1.3.3质粒转染和细胞分组 转染前1 d制备A549单细胞悬液接种于6孔板,当细胞生长达50%~80%汇合时,按NanoFectinTM说明书步骤分别进行pGCsiU6/puro/GFP-bFGF-siRNA和pGCsiU6/puro/GFP-NC-siRNA质粒转染。转染48 h后,按1∶5传代,细胞贴壁良好后加入1 mg/ml嘌呤霉素进行筛选,2 w后荧光显微镜下挑选多个表达绿色荧光蛋白(GFP)的单一抗性克隆扩增培养,得到稳定表达pGCsiU6/puro/GFP-bFGF-siRNA的A549细胞(干扰质粒组)和稳定表达pGCsiU6/puro/ GFP-NC-siRNA的A549细胞(阴性对照组)。以未转染的A549细胞作为空白对照组。
1.3.4Real-time PCR检测bFGF mRNA表达 引物序列设计:bFGF上游引物为5′-AGTGTGTG CTAACCGTTACCT-3′,下游引物为5′-ACTGCCCAGTTCGTTTCAGTG-3′;以GAPDH为内参对照,上游引物为5′-ATGGGGAAGGTGAAGGTCG-3′,下游引物5′-GGGGTCATTGATGGCAACAATA-3′。收集各组细胞,按Trizol说明书提取总RNA,测纯度、浓度。按照PrimeScriptTM反转录试剂盒合成cDNA,反应条件为37 ℃ 15 min,85 ℃ 5 s。然后按SYBR Green反应条件进行实时PCR反应,条件设定为:95 ℃预变性3 min,95 ℃变性30 s,58 ℃退火30 s,72 ℃延伸30 s,循环35次,72 ℃再延伸5 min。扩增结束后绘制熔解曲线及扩增曲线,以SDS2.0软件分析Ct值,将各bFGF Ct值减去GAPDH Ct值得ΔCt值,ΔCt越高,mRNA表达越低,以2-△△CT法计算mRNA相对表达量。
1.3.5Western印迹检测OCT-4表达 收集用预冷的PBS洗涤3次的细胞,加适量RIPA及PMSF混合液裂解30 min,4 ℃离心5 min,取上清得细胞总蛋白,BCA法测蛋白浓度。SDS-PAGE:蛋白样品加上样缓冲液混匀,95 ℃ 5 min变性后取50 μg蛋白上样,压缩胶70 V 20 min,浓缩胶100 V 60 min电泳,转膜后,5%脱脂奶粉封闭2 h,加一抗(OCT-4的浓度为1∶800,内参β-actin为1∶1 000)4 ℃过夜,PBST洗膜3次,每次10 min,加二抗(1∶5 000)室温孵育90 min,洗膜后进行ECL法发光、显影、定影。捷达凝胶系统分析各条带积分光密度值(IOD),以OCT-4的IOD/β-actin的IOD,计算蛋白相对表达量。
1.3.6CCK-8实验检测细胞增殖能力 收集细胞制备细胞悬液,调整细胞浓度为1×104/ml,取100 μl接种7块96孔板,边缘孔以PBS填充,同时设置空白孔(培养基和CCK-8,无细胞),对照孔(培养基和细胞),置37 ℃,5%CO2孵育过夜,每孔加入10 μl CCK-8溶液,4 h后测定490 nm各孔的吸光度值(A值),连测7 d,以时间为横轴,每天的A值为纵轴绘制生长曲线。实验重复3次。
1.3.7克隆形成实验 制备单细胞悬液计数,取100个细胞种于35 mm培养皿中,置于37 ℃、5% CO2孵箱中培养2 w,吸除培养液,甲醇固定15 min,吉姆萨染色液染色10 min,PBS洗涤,于显微镜下计数克隆数并记录。克隆形成率(%)=克隆形成数/接种细胞数×100%。
2 结 果
2.1转染效率的鉴定 转染48 h后用荧光显微镜观察转染效率,大于80%。见图1。
2.2RT-PCR结果 以空白对照组A549细胞的bFGF mRNA表达水平为参照,质粒干扰组及阴性对照组的mRNA相当表达分别为(0.209±0.044)倍及(1.029±0.215)倍,干扰组明显低于阴性及空白对照组(P<0.01);阴性对照组和空白对照组间差异无统计学意义。

图1 荧光显微镜下观察A549细胞的转染效率
2.3OCT-4蛋白的表达水平 质粒干扰组、阴性对照组和空白对照组OCT-4蛋白相对表达分别为0.440±0.046、0.748±0.102、0.726±0.019,与阴性对照组和空白对照组相比,质粒干扰组的OCT-4蛋白表达明显下降(P<0.01)。见图2。
2.4细胞生长曲线图 第1天各组细胞增殖没有差别,第2~7天质粒干扰组细胞生长明显受抑,增殖活力明显低于阴性对照组和空白对照组(各P<0.01),阴性对照组和空白对照组间差异无统计学意义。见图3。

1:空白对照;2:阴性对照组;3:质粒干扰组
2.5抑制bFGF表达对细胞克隆形成能力的影响 图中单个紫色小点为一个克隆集落,质粒干扰组、阴性对照组及空白对照组细胞克隆形成率分别为(12.25±2.97)%、(42.75±4.50)%、(44.25±3.31)%,质粒干扰组克隆形成能力明显低于阴性对照组和空白对照组(各P<0.01),而阴性对照组和空白对照组之间差异无统计学意义。见图4。

与质粒转染组比较:1)P<0.05,2)P<0.01
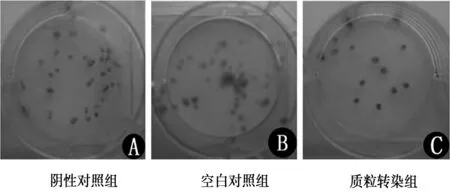
图4 转染后各组A549细胞的克隆集落
3 讨 论
bFGF作为bFGF/FGFR信号通路的关键蛋白,由定位于染色体4q 26~27基因编码,因其上游起始密码子的不同,可产生分子量在18~34 kD之间的多种异构体,这些异构体通过自分泌、旁分泌或者内分泌的形式分泌,与细胞表面的受体接触后通过PKC、Ras /Raf/MEK /ERK、PI3K等途径将信号传递至胞内。
肿瘤的发生、发展很多都和细胞信号通路传导异常密切相关,bFGF/FGFR信号通路作为众多重要信号通路之一,不仅在维持胚胎干细胞〔1〕、骨髓间充质干细胞〔2〕、神经干细胞〔3〕等正常干细胞自我更新、多潜能分化等特性方面起重要作用外,而且在促进肿瘤细胞增殖、侵袭、耐药、抗凋亡及肿瘤新生血管形成等方面起重要作用。已有研究表明,bFGF在脑胶质瘤〔4〕、乳腺癌〔5〕、黑色素瘤〔6〕、肝癌等〔7〕多种肿瘤中过表达。Zhao等〔8〕对肺癌组织及正常肺组织行免疫组化发现,80.9%肺癌组织表达bFGF,明显高于正常肺组织,而且bFGF过表达预示着更差的总生存,说明bFGF在肺癌发生发展中发挥重要作用。
转录因子OCT-4已被证实可作为胚胎干细胞的标志物〔9〕,其在干细胞高表达,一旦干细胞分化,其表达迅速下降或不表达。OCT-4对肿瘤形成及发展也起重要作用,这种作用在乳腺癌〔10〕、肺癌〔11〕、前列腺癌〔12〕、口腔癌〔13〕等肿瘤干细胞中得到证实。Meng等〔14〕研究发现A549细胞系中约有45%的为肿瘤始发细胞,它们具有自我复制、成瘤等特性。Vera等〔15〕对多种人癌细胞系进行化疗药物处理,发现存活细胞富集肿瘤干细胞。本研究检测A549表达OCT-4。
由于bFGF容易变性降解、半衰期极短,限制了bFGF单克隆抗体的使用。而RNA干扰作为一种重要的基因转录后沉默技术,其核心原理是将特异性同源双链RNA导入细胞,导致目的基因不表达或表达下降,由于其不影响DNA的复制和转录,与反义寡核苷酸干扰技术相比,RNAi具有高度特异性、抑制效率高、可持久传代等优点,已广泛应用于基因治疗研究〔16〕。本研究通过RNA干扰可以有效抑制肺癌A549细胞中bFGF在mRNA水平的表达。Greber等〔17〕在人胚胎干细胞中发现bFGF通过调节TGF-β受体信号通路而维持Oct-4、Nanog和Sox2的表达,促进干细胞的自我更新,本研究发现bFGF表达抑制后,A549细胞的增殖能力及自我更新能力降低,,推测可能bFGF被抑制后下调OCT-4表达,影响肺癌A549中的干细胞的特性有关。总之,本研究通过应用RNAi技术,不仅有效抑制肺癌A549细胞中bFGF的表达,而且证明bFGF信号通路被抑制可能通过下调OCT-4,抑制肺癌细胞的增殖及自我更新能力等干细胞特性。
4 参考文献
1Dvorak P,Dvorakova D,Hampl A.Fibroblast growth factor signaling in embryonic and cancer stem cells〔J〕.FEBS Lett,2006;580(12): 2869-74.
2Schmidt A,Ladage D,Schinklthe T,etal.Basic fibroblast growth factor controls migration in human mesenchymal stem cells〔J〕.Stem Cells,2006;24(7):1750-8.
3王占尧,曹 磊,姬西团.bFGF和BDNF对小鼠神经干细胞体外增殖分化的影响〔J〕.中华神经外科疾病研究杂志,2010;9(5):435-7.
4Wang DL,Wang YF,Shi GS,etal.Correlation of hTERT expression to maspin and bFGF expression and their significance in glioma〔J〕.Ai zheng,2007;26(6):601-6.
5Fuksiewicz M,Kowalska M,Kotowicz B,etal.Serum soluble tumour necrosis factor receptor type I concentrations independently predict prognosis in patients with breast cancer〔J〕.Clin Chem Lab Med,2010;48(10):1481-6.
6Fontijn D,Bosch LJ,Duyndam MC,etal.Basic fibroblast growth factor- mediated overexpression of vascular endothelial growth factor in IF6 human melanoma cells is regulated by activation of PI3K and p38 MAPK〔J〕.Cell Oncol,2009;31(3):179-90.
7Wang XH,Sun X,Meng XW,etal.Beta-catenin siRNA regulation of apoptosis- and angiogenesis-related gene expression in hepatocellular carcinoma cells:potential uses for gene therapy〔J〕.Oncol Rep,2010;24(4):1093-9.
8Zhao M,Gao FH,Wang JY,etal.JAK2/STAT3 signaling pathway activation mediates tumor angiogenesis by upregulation of VEGF and bFGF in non-small-cell lung cancer〔J〕.Lung Cancer,2011;73: 366-74.
9Lengner CJ,Welstead GG,Jaenisch R.The pluripotency regulator Oct-4:a role in somatic stem cells〔J〕?Cell Cycle,2008;7(6):725-8.
10Bussolati B,Grange C,Sapio A,etal.Endothelial cell differentiation of human breast tumor progenitor cells〔J〕.J Cell Mol Med,2009; 13(2):309 -19.
11Chen YC,Hsu HS,Chen YW,etal.Oct-4 expression maintained CD133-positivrcell 〔J〕.PLoS One,2008;3(7):e2637.
12Gu G,Yuan J,Wills M,etal.Prostate cancer cells with stem cell characteristics reconstitute the original human tumor in vivo 〔J〕.Cancer Res,2007;67(10):4807-15.
13Chiou SH,Yu CC,Huang CY,etal.Positive correlation of Oct-4 and Nanog in oral cancer stem-like cells and high-grade ora squamous cell carcinoma 〔J〕.Clin Cancer Res,2008;14(13): 4085-95.
14Meng X,Wang X,Wang Y.More than 45%of A549 and H446 cells are cancer initiating cells:evidence from cloning and tumorigenic analyses 〔J〕.Onco Rep,2009; 21(4):995-100.
15Vera Levina,Adele M,Marrangoni A,etal.Drug-selected human lung cancer stem cells:cytokine network,tumorigenic and metastatic properties 〔J〕.PloS One,2008;3(8):e3077.
16Kim DH,Rossi JJ.Strategies for silencing human disease using RNA inter- ference〔J〕.Nat Rev Genet,2OO7; 8(3): 173-84.
17Greber B,Lehrach H,Adjaye J.Fibroblast growth factor 2 modulates transforming growth factor beta signoling in mouse embryonic fibroblasts and human ESCs(hESCs)to support hESC self-renewal〔J〕.Stem Cells,2007;25(2):455-64.
