甜玉米与糯玉米的染色体核型分析
2014-08-28刘继琳刘彩霞郭佳静杨国强黎杰强
刘继琳, 刘彩霞, 郭佳静, 杨国强, 黎杰强
(广州市华南师范大学生命科学学院,广州 510631)
玉米是一年生禾本科草本植物,是重要的粮食作物和饲料来源.玉米按其籽粒形状、胚乳性质及稃壳有无,可分为9个亚种或类型[1],其中的甜玉米和糯玉米是新兴的特种类型玉米.甜玉米(又称水果玉米)因其籽粒在乳熟期含糖量较高而得名,其原产于美洲,是一类胚乳隐性突变体,由su1,su2,sh1,sh2,sh4,se,bt1,bt2等系列隐性突变基因控制[2].根据遗传基础的不同,甜玉米可分为普甜(susu)、超甜(sh2sh2/sh2sh2sh2)、脆甜(bt1bt1/bt1bt1bt1和bt2bt2/bt2bt2bt2)和加强甜(su1sesu1se/su1sesu1sesu1se)4种类型,其胚乳各种糖分含量因类型而异[2-3].糯玉米(又称蜡质玉米)为原产于我国西南地区的特殊玉米变异类型,是由玉米第9对染色体短臂上的靠近着丝点处(9s-59)的纯合隐性基因wxwx控制[4],其籽粒淀粉全部为支链淀粉,粘性较强.
每一生物体内特定的染色体组成染色体组型.染色体组型分析(也称核型分析)是对染色体数目、形态、着丝点位置、长度、臂比和随体等进行定性或定量的分析,是研究物种演化、分类以及染色体结构、形态与功能之间的关系所不可缺少的重要手段[5-6].关于玉米染色体核型的研究,Kuwada[7]证实玉米的染色体数目为2n=20.Chen[8]和Filion[9]分别描述了其体细胞染色体的核型.Neuffer[10]提出了玉米体细胞染色体的核型模式图.张赞平等[11]分析比较了玉米8个亚种、2个亚型和2个杂交品种染色体核型,发现玉米各亚种的核型进化,基本符合由对称向不对称发展趋势.宗成志[12]对杂交玉米华甜1号与登海9号的染色体核型分析,认为两者染色体的臂长、臂比和形态既有相似之处,也存在一定差异.染色体核型的变化是了解植物系统发育和进化的一个重要方面,可为植物的分类鉴定提供细胞学依据[13].甜玉米和糯玉米除了糖份含量、营养成分、籽粒形状、口感等存在差异外,其起源、株形、叶形等均存在一定的差异,对它们细胞学水平的分析(包括核型分析)报道少.本研究采用常规压片技术,对甜玉米和糯玉米进行核型分析,旨在为探讨特种玉米的进化和系统发育提供细胞学依据.
1 材料与方法
1.1 材料、试剂及仪器
甜玉米采用L-11株系作为材料, 由美国引进的超甜玉米经过8代自交选育而成的二环系;糯玉米采用L-70株系作为材料, 由京科糯玉米经过6代自交选育而成的二环系.材料选育地点:华南师范大学生物园,材料田间农艺性状表现稳定.纤维素酶(美国)、果胶酶(丹麦);显微镜Nikon80i(尼康)及相关的照相系统.
1.2 染色体制片方法和核型分析
选择均匀一致、干燥的甜玉米和糯玉米种子分别置于水中浸泡1 h和2 h,种子吸胀后转到培养皿中,加少许水,30 ℃下培养.根长1.0~1.5 cm时,将根尖切下置于w(秋水仙素)=0.1%溶液常温处理3 h.用蒸馏水冲洗2~3次,移入新配制的固定液(V(甲醇)∶V(冰醋酸)=3∶1)中,4~15 ℃下固定4 h.用φ(乙醇)=90%冲洗2次后转入70%乙醇中,置4 ℃冰箱中保存待用.将根尖置于2.5%纤维素酶和果胶酶等量混合液中,25 ℃酶解3 h,吸掉酶液,蒸馏水冲洗2~3次,改良品红染液,染色10 min,压片法观察.用Nikon 80i 显微镜在10×40倍镜下拍照染色体形态好且分散的细胞.取分散良好、着丝点清晰的染色体中期分裂相图片30张进行染色体计数,统计材料的染色体数目.
核型分析按照李懋学等[14]的标准,核型分类按 Stebbins[15]的分类标准,按 Arano[16]方法计算核型不对称系数(As.k%),按 Kuo S R 等[17]的方法确定染色体相对长度系数(Index of relative length),采用Levan[18]着丝点命名系统.每种材料观察 30个以上中期分裂相进行染色体计数,并选取5个分散良好的中期分裂相,用尼康显微镜(80i)在100倍油镜下拍照.采用手工剪分,取其平均值供核型分析,用Excel绘制核型模式图[19].
2 结果与分析
2.1 甜玉米的染色体核型分析
结果表明,甜玉米(L-11株系)的染色体数目为2n=20(图1),与张赞平等[11,20]对玉米的8个亚种核型分析所得的染色体数目相符合(2n=20),由甜玉米的染色体核型(图2)、核型模式(图3)和染色体参数(表1)得出的甜玉米(L-11株系)核型公式:2n=2x=20=2M+12m+6sm(2Sat),即其染色体组成是由1对正中着丝粒染色体、6对中着丝粒染色体和3对近中部着丝粒染色体组成,且在3对近中部着丝粒染色体中有1对具随体.甜玉米最长与最短的染色体比值为2,臂比大于2的染色体比例为20%,核型不对称系数为59.66%,核型属2B型.染色体相对长度变化范围为3.44%~6.86%;染色体相对长度系数组成为2n=20=2L+4M2+12M1+2S,由1对长染色体(No.1),2对中长染色体(No.2,3),6对中短染色体(No.4-9)和1对短染色体(No.10)组成.

图1 甜玉米的有丝分裂中期染色体(×400)

图2 甜玉米的染色体核型

图3 甜玉米的染色体核型模式图

序号相对长度/%(短臂+长臂=全长)相对长度系数臂比着丝点指数类型13.00+3.86=6.861.371.2943.75m22.58+3.00=5.581.121.1746.15m32.14+3.43=5.571.121.6038.46m42.58+2.58=5.161.031.0050.00M52.36+2.58=4.940.991.1047.83m61.50+3.43=4.930.992.2930.44sm71.72+2.79=4.510.901.6338.10m81.29+3.00=4.290.862.3330.00sm91.29+2.15=3.440.691.6737.50m101.72+3.00=4.720.941.7536.36sm*
*具随体染色体,随体长度未计算在内,下表同
2.2 糯玉米的染色体核型分析
糯玉米(L-70株系)的染色体数目为2n=20(图4).这与张赞平等[11,20]对糯玉米材料根尖细胞染色体数目一致(2n=20).

图4 糯玉米的有丝分裂中期染色体(×400)
由糯玉米的染色体核型(图5)、核型模式(图6)和染色体参数(表2)可以得出糯玉米的核型公式为:2n=2x=20=2M+8m(2Sat)+10sm,即其染色体组成是由1对正中着丝粒染色体、4对中着丝粒染色体和5对近中部着丝粒染色体组成,且在4对中着丝粒染色体中有1对是具随体的.最长与最短的染色体比值为1.777 8,臂比大于2的染色体比例为10%,核型不对称系数为60.41%,核型属2A型.染色体相对长度变化范围为3.67%~6.54%;染色体相对长度系数组成为2n=20=2L+8M2+6M1+4S,由1对长染色体(No.1),4对中长染色体(No.2-5),3对中短染色体(No.6-8)和2对短染色体(No.9,10)组成.
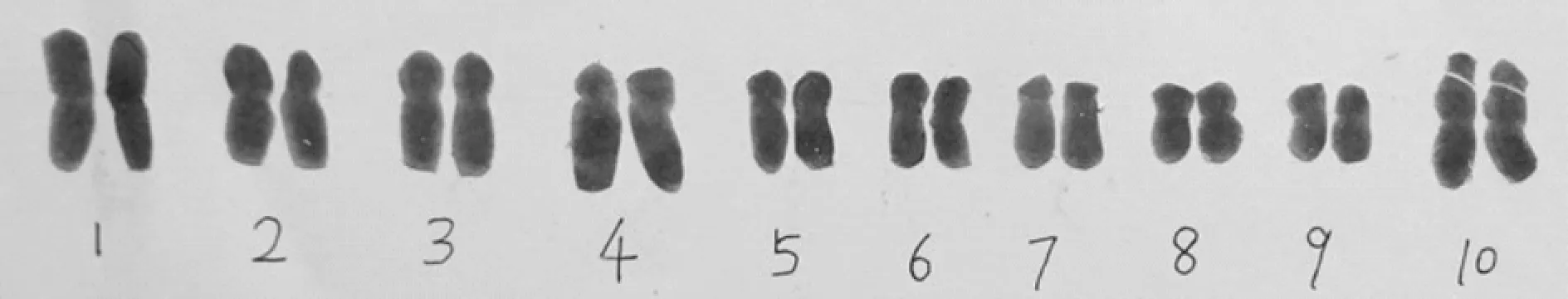
图5 糯玉米的染色体核型

图6 糯玉米的染色体核型模式图

序号相对长度/%(短臂+长臂=全长)相对长度系数臂比着丝点指数类型13.27+3.27=6.541.311.0050.00M22.45+3.27=5.721.141.3342.86m32.04+3.67=5.711.141.8035.71sm41.63+4.08=5.711.142.5028.57sm51.84+3.27=5.111.021.7836.00sm62.24+2.45=4.690.941.0947.83m71.43+2.86=4.290.862.0033.33sm81.63+2.04=3.670.731.2544.44m91.22+2.45=3.670.732.0033.33sm102.04+2.86=4.900.981.4041.67m*
2.3 甜玉米与糯玉米的核型比较
表3显示,甜玉米与糯玉米的核型和染色体参数有相同点:(1)它们的染色体都是20条,两者的核型组成都含有正中着丝粒、中部着丝粒和近中部着丝粒染色体,且都有1对染色体带有随体,与对玉米的各亚种核型结果基本[20]相同.(2)2种材料的染色体相对长度变化幅度接近,甜玉米是3.44%~6.86%,糯玉米是3.67%~6.54%.(3)核不对称系数也接近,甜玉米是59.66%,而糯玉米则是60.41%,均属于较不对称核型.根据Stebbins[13]的理论,不对称程度越高的生物,其染色体变异越大,进化程度越高.因此,甜玉米(L-11株系)与糯玉米(L-70株系)的核型在系统演化上属于较进化的类群.

表3 甜玉米与糯玉米的核型和染色体参数比较Table 3 Comparison of karyotypes and chromosome parameters of sweet corn and waxy corn
甜玉米与糯玉米的核型存在差异:(1)按照Stebbins[15]标准核型分类,甜玉米的核型是2B型,糯玉米的核型是2A型.(2)比较两者核型公式,甜玉米由1对正中着丝粒染色体、6对中着丝粒染色体和3对近中部着丝粒染色体组成,且在3对近中部着丝粒染色体中有1对具随体,臂比>2的比率为20%.糯玉米由1对正中着丝粒染色体、4对中着丝粒染色体和5对近中部着丝粒染色体组成,且在4对中着丝粒染色体中有1对是具随体的,臂比>2的比率为10%.随体分布位置的不同,可见这2个物种之间存在明显的细胞学分化.(3)比较两者染色体长度组成,L、M1、M2、S所占的比例有所不同,甜玉米为1∶2∶6∶1,而糯玉米则是1∶4∶3∶2.综合比较两者核型数值,根据Stebbins[15]的理论,在生物的进化过程中,染色体核型是由对称性向非对称性演化的.Stebbins通过对植物核型资料分析,将染色体核型按对称性程度的高低分为12种类型(1A-4A,1B-4B,1C-4C).核型对称性程度越高的生物,其染色体变异越小,进化程度也越低;而非对称性程度越高的生物,其染色体变异越大,进化程度越高.按照这一观点,甜玉米的核型为2B型,在进化上应属于程度较高的类型;而糯玉米的核型为2A型,应属于进化程度较低的种类.所以,甜玉米的进化程度较高.
通过以上比较,说明玉米的不同种类之间既有种属间的相似性,同时也存在一定的变异性.可能与遗传中的DNA序列在进化过程中歧化相关,从而导致类型品种之间产生一定差异并扩大,差异积累到一定程度,该变化就可能引起染色体某些部位结构的变化,从而反映到染色体形态的变化[12].
3 讨论
甜玉米与糯玉米是属于不同类型的2个玉米品种.两者的起源不同,甜玉米主要起源于美洲,而糯玉米是我国西南的特殊变异类型,对环境的适应性更强.甜玉米引入后经过长时间的驯化种植、基因提纯、选择,对国内的环境(包括土壤、气侯等)适应能力逐步加强,也能很好地适应国内的生产种植.但它们的起源演化过程的不同,可能导致它们核型的差异.另外,栽培居群的不同也可能导致核型的差异.由于不同的生长地点在气候条件、土壤性质等生境方面的差异,同种植物很有可能因适应不同环境的结果而形成核型多态性[21],在其他物种的染色体研究中也有不同居群核型差异的报道[22-25].此外,在人工杂交育种栽培实践中,引种方法、栽培技术等因素也可能会引起变异的发生,染色体数目和结构也会发生改变[26-27].
张赞平[20]指出甜玉米的核型公式为2n=10m(2SAT) +10sm,由5对中着丝粒染色体和5对近中着丝粒染色体组成,其中第6染色体短臂具有随体;最长染色体与最短染色体的比值为2.05,臂比大于2的染色体比例为20%,核型不对称系数为59.78%,核型为2B型.除核型公式外,其它指标数值与本实验的甜玉米染色体核型数值基本相符,说明来源较接近.糯玉米的核型公式为2n=14m+4sm(2Sat)+2st,由7对中着丝粒染色体、2对近中着丝粒染色体和1对近端着丝粒染色体组成,其中第6染色体短臂具有随体;最长染色体与最短染色体的比值为2.17,臂比大于2的染色体比例为20%,核型不对称系数为59.24%,核型为2B型.糯玉米核型为2A型,说明两者的材料来源差异较大,根据Stebbins[15]的理论,本实验的糯玉米是相对进化程度稍低的材料.说明目前甜玉米筛选的资源较窄,糯玉米资源较广.其是长期自然突变与人工选择的结果.核型分析等细胞学资料多应用于讨论种和种以下类群的划分,大多数情况对说明各属是同质还是异质、是近亲还是远亲有很大的价值[13].所以,本实验对甜玉米和糯玉米的核型分析结果为甜、糯玉米这两个类型间的亲缘关系以及分类提供有效的细胞学依据.
对材料预处理、压片技术等,可能导致核型参数产生差异,比如染色体长度在预处理过程中受药剂种类、浓度及处理时间的影响,可能会明显缩短[25,28].玉米染色体较小,给制片与核型分析带来一定的影响.因此要获得准确的结果,合适的预处理浓度和时间、制作清晰、高质量的染色体玻片标本是关键,运用染色体分带技术作核型分析,更能准确反映核型上的差异.G-带是染色体本身特征的本质反应,结合G-带分析可有效地提高了核型分析的精确度[12].同样,植物染色体的C-带显示的部位主要是组成型异染色质,它们主要是由分布在染色体末端和着丝粒附近的DNA重复序列组成,不同物种的染色体C-带带型具有明显的差异,根据染色体的C-带可以进行物种的鉴别[29].原位杂交技术、分子遗传标记等技术的应用也为物种的起源、亲缘关系的准确分析提供了更广阔的前景.
参考文献:
[1] 玉米遗传育种学编写组.玉米遗传育种学[M].北京:科学出版社,1979:5-7.
[2] 刘纪麟.玉米育种学[M]. 北京: 农业出版社,1991:342-348.
[3] 罗高玲,吴子恺.甜玉米和糯玉米遗传基因的研究进展[J].广西农业科学,2003(3):24-26.
Luo G L,Wu Z K.Advances about the research of sweet and glutinous maize[J].Guangxi Agriculture Science,2003(3):24-26.
[4] Collins G N.A new type of Indian corn of China[J]. Bureau of Plant Industry,1909(161):1-31.
[5] 李懋学,张敩方.植物染色体研究技术[M].哈尔滨:东北林业大学出版社,1991:1-20.
[6] Hu J Y,Su Z X,Yue B L,et al.Progress of karyotype analysis method in plants research[J].Journal of Sichuan Teachers Collage: Natural Science,2002,23(3): 239-244.
[7] Kuwada Y.Meiosis in the pollen mother cells ofZeamaysL[J]. Botanical Magazine (Tokyo),1911,25: 163-181.
[8] Chen C C.The somatic chromosomes of maize[J].Canadian Journal of Genetics and Cytology, 1969,11:752-754.
[9] Filion W G,Walden D B.Karyotype analysis:The detection of chromosomal alterations in the somatic karyotype ofZeamaysL[J].Chromosoma,1973,41:183-194.
[10] Neuffer M G.Handbook of geneties.(Ed.R.C.king)[M].New York:Plenum Press,1974.
[11] 张赞平,李懋学.玉米8个栽培亚种(类型)的核型和C-带带型的比较研究[J].遗传学报,1990,17(2):86-93.
Zhang Z P,Li M X.A comparative study on the karyotypes and C-banding patterns of 8 subspecies (types) of corns [J]. Journal of Genetics,1990,17(2):86-93.
[12] 宗成志,李姝婧,吴士筠.杂交玉米华甜1号与登海9号染色体G-显带核型分析[J].现代农业科学,2008(10):1-3.
Zong C Z,Li S J,Wu S Y.Chromosome G- banding karyotype analysis of hybrid corns, Huatian No.1 and Denghai No.9 [J].Modern Agricultural Science,2008(10):1-3.
[13] 吴甘霖.核型分析在细胞分类学中的应用[J]. 生物学杂志,2006,23(1):39-42.
Wu G L. Application of karyotype analysis on cytotaxonomy [J]. Journal of Biology, 2006, 23(1):39-42.
[14] 李懋学,陈瑞阳.关于核型分析的标准化问题[J].武汉植物学研究,1985,3(4):297-302.
Li M X, Chen R Y. The standard problems about karyotype analysis [J].Wuhan Botanical Research,1985,3(4):297-302.
[15] Stebbins G L.Chromosome evolution in hingher plants[M].London: Edward Arnoldy Ltd, 1971:88-93.
[16] Arano H L.Cytological studies in subfamily Carduoideae (Compositae) of Japan IX[J]. Botanical Magazine (Tokyo),1963,76:32-39.
[17] Kuo S R,Wang T T,Huang T C.Karyotype analysis of some Formosan gymnosperms[J]. Taiwania,1972,17(1):66-80.
[18] Levan A,Fredga K,Sandberg A A.Nomenclacture for centromeric position on chromosomes [J].Hereditas,1964,52(2):197-201.
[19] 乔永刚,宋芸.利用EXCLE制作核型模式图[J].农业网络信息,2006(10):97-98.
Qiao Y G, Song Y. Making karyotype model charts by EXCLE [J].The Agricultural Network Information,2006(10):97-98.
[20] 张赞平.玉米各亚种的核型研究[J].华北农学报,1995,10(3):60-67.
Zhang Z P.Karyotype studis on each subspecies of maize[J].North China Agricultural Journal,1995,10(3):60-67.
[21] 李倩,张桂彬,孙崇苹.两种观赏吊兰的核型分析[J].北方园艺,2009(3):202-204.
Li Q,Zhang G B, Sun C P. Karyotype analysis of two kinds of ornamental bracketplants [J]. Northern Horticulture,2009(3):202-204.
[22] 戴小红,张延龙,牛立新.百合属4种植物的核型研究[J].西北植物学报,2006,26(1): 50-56.
Dai X H, Zhang Y L, Niu L X. A karyotype study on 4 species of genus Lilium [J].Northwest Botany Journal,2006,26(1):50-56.
[23] 刘华敏,智丽,赵丽华,等.四种野生百合核型分析[J].植物遗传资源学报,2010,11(4): 469-473.
Liu H M,Zhi L, Zhao L H, et al. Karyotype analysis of four Wild Lilium species [J]. Journal of Plant Genetic Resources,2010,11(4):469-473.
[24] 魏玉杰,何庆祥,张金文,等.中国不同地区罂粟的核型比较[J].草业学报,2012,21(2): 171-179.
Wei Y J,He Q X,Zhang J W, et al.Karyotype comparisons of poppies of different regions in China[J].Grassland Journal,2012,21(2):171-179.
[25] 张芹,李保会,张凡.3种补血草属植物的核型分析[J].河北农业大学学报,2013,36(2):65-68.
Zhang Q,Li B H,Zhang F.Karyotype analysis of 3 species of Limonium plant[J].Journal of Hebei Agricultural University,2013,36(2):65-68.
[26] 詹园凤,党选民,曹振木,等.两个茄子品种的核型分析[J].植物遗传资源学报,2009, 10(2): 283-285.
Zhan Y F,Dang X M,Cao Z M, et al. Karyotype analysis of two eggplant varieties[J].Journal of Plant Genetic Resources,2009,10(2):283-285.
[27] 祝海燕,罗双霞,陈雪平,等.几种茄科蔬菜的核型分析和比较[J].河北农业大学学报,2013, 36(1):21-24.
Zhu H Y,Luo S X,Chen X P, et al.The karyotype analysis and comparison of several solanaceous vegetable[J].Journal of Hebei Agricultural University,2013,36(1):21-24.
[28] 王桃银,郭巧生,汪涛.21个药用菊花栽培类型的核型分析[J].南京农业大学学报,2012,35(6): 13-18.
Wang T Y,Guo Q S,Wang T. Karyotype analysis of 21 cultivars of medicinal chrysanthemum[J]. Journal of Nanjing Agricultural University,2012,35(6):13-18.
[29] 陈全站,杨平.4种栽培大麦染色体组型和C-带带型的比较分析[J].南京晓庄学院学报,2007(3): 57-61.
Chen Q Z,Yang P.The comparative analysis of chromosome karyotypes and C-banding patterns of 4 kinds of cultivated barley [J].Journal of Nanjing Xiaozhuang University,2007(3):57-61.
