中龄和老龄杉木人工林细根序级形态和呼吸特征
2014-08-22熊德成黄锦学杨智杰陈光水杨玉盛
熊德成,黄锦学,杨智杰,陈光水,杨玉盛
(1.福建省湿润亚热带山地生态国家重点实验室——省部共建国家重点实验室培育基地,福州350007;2.福建师范大学地理研究所,福州350007)
细根 (直径≤2 mm),是树木根系的组成部分,具有重要的生理和生态学功能[1]。它作为提供植物养分和水分的“源”和消耗碳的“汇”,已成为生态系统生态学及全球变化研究中最受关注的热点[2]。近来研究表明,根系是由不同的分支等级构成,它们在形态、结构和功能上存在显著差异,若简单地把直径<2 mm(或<1 mm)的根视为细根,则忽视了细根形态结构的异质性[3-4]。这一观点已被国外自Pregitzer等[3,5]有关根序的论文发表以来一系列以根序为角度开展根系及功能的研究所证实[6-7]。近年来,国内有关细根异质性方面的文章从温带到亚热带再到热带,从阔叶树种到针叶树种都屡有报道[8-13],同时以往对细根序级的研究主要还集中在对单一林龄树种及不同树种根序之间的比较,有关林龄对细根序级相关特征的影响仍鲜有报道。目前中国亚热带有关阔叶林树种细根根序特征的研究已有一定报道,而有关针叶林的研究还相对比较缺乏,那么针叶林随细根根序的变化形态和生理特征有何差异,林龄又是如何影响这些特征均缺乏论证。
杉木 (Cunninghamia lanceolata)是中国南方重要的造林和用材树种,在中国人工林中占有重要地位。基于以上原因本研究在福建省南平市王台镇溪后村安曹下选取19年生和91年生杉木人工林为研究对象,重点揭示林龄对细根序级形态及呼吸特征的影响。这为细根周转估算以及杉木人工林在不同生长期碳分配格局的变化规律提供一定的参考,同时也对揭示林龄对森林碳收支影响具有一定作用。
1 材料与方法
1.1 试验地概况
试验地选择在福建省南平市王台镇溪后村安曹下 (26°28'N,l17°57'E),本区属武夷山系南伸支脉,属亚热带季风气候,平均海拔200 m左右,多年平均气温19.3℃,多年平均降水量1 669 mm,降水多集中在3~8月,年均蒸发量1 413 mm,年均相对湿度为83%;土壤是由燕山晚期白云母化中细粒花岗岩发育的山地暗红壤。土壤厚度在100 cm以上,土壤表层疏松,但均含有一定量的石砾,质地为砾质轻壤土。
本研究选取的2个林分地理位置上大约相距1 km左右,其中19年生杉木人工林为1991年造林而成,灌木层以杜茎山 (Maesa japonica)、寒莓 (Rubus buergeri)、粗叶榕 (Ficus hirta)等为主,草本层以金星蕨 (Parathelypteris glanduligera)、长江蹄盖蕨 (Athyrium iseanum)、鳞毛蕨 (Dryopteris austrtiaca)、狗脊 (Woodwardia japonica)等为主,盖度约90%;91年生杉木人工林为1919年插条造林而成,是世界上现存杉木人工林年龄和单位面积蓄积量最大的林分之一,具有极大的科研价值,林下灌木层以杜茎山 (Maesa japonica)、寒莓 (Rubus buergeri)、粗叶榕 (Ficus hirta)等为主,草本层以芒萁 (Dicranopteris dichotoma)、水龙骨 (Polypodiodes niponica)、狗脊 (Woodwardia japonica)为主,林下植被盖度约100%。2个林分的本底条件参见表1(2006年调查数据):

表1 2个林分的基本特征及表层土壤性质Table1 Forest characteristics and surface soil properties of two study sites
2 研究方法
2.1 根系采样与处理
细根采样集中在2010年7月份,采样时分别在19年生和91年生杉木林中选取胸径大小相近的5棵成年树,根据Guo等[4]的完整土块法在每棵树树干基部1~1.5 m范围用铁铲挖取1个20 cm×20 cm×20 cm大小的土块,然后将里面的所有根段全部取出,并放置在湿纱布中以保持其活性,并立即带回实验室,随即用低温去离子水分别清理掉根系表面上的土壤和杂质,装进贴有标签的自封袋中并装入冷藏箱内以保持其活性。根据Pregitze等[3]的分级方法,每个土块选取1个完整根段将其分成5级,最远端的根尖定为1级根,1级根交汇处为2级根,依次类推到5级根,其余所有根系根据同样的方法进行分级处理,不同等级的根经处理后分别装入已标记好的玻璃皿中,并放入冷藏箱保存。
2.2 根系形态指标测定
细根分级完成后将处理好的样品使用数字化扫描仪Espon scanner分别进行扫描 (各序级均进行3次重复实验),扫描完成后用Win RHIZO(Pro 2005b)根系图像分析软件对扫描后的细根图像进行形态指标的分析,而后将所有根系样品分别放入65℃的烘箱48 h烘至恒重,以计算出各个序级的干重。通过扫描分析可以得出细根的直径、根长、体积等数据,并通过进一步的计算得出比根长,组织密度等。相关计算公式为:根长 (cm)=总长度 (cm)/细根数量;比根长=根长 (m)/干重 (g);组织密度(g·cm-3)=干重 (g)/体积 (cm-3)。
2.3 根系呼吸测定
进行根系扫描时选取一部分根系同步对不同序级细根的呼吸进行测定。测定时将分离好的2个林分的各个土块的各级细根分别选取大约0.5 g放入Li-6400便携式光合测定仪的叶室中,1、2级根由于体积和直径较小可直接放进叶室测定,待低级根测定完成后立即对较高级根进行测定。实验前用一体积较大的塑料瓶与Li-6400出气口相连接以作为气流的缓冲装置。测定过程中主要是大约5~10 min后待气体流动稳定后开始测定根系释放的CO2浓度,主要是进行手动读数,每次测定读取5个数值,每个土块的各级细根均进行3次重复测定,共测得25个数值,然后取其平均值,各个序级5个土块共得到5组平均值,然后用这5组平均值进行比根呼吸的计算最后求得平均比根呼吸。整个实验都是在有空调控温的室内进行测定 (25℃),根系释放的水汽通过Li-6400便携式光合测定仪的干燥剂吸收,使叶室中的空气湿度保持稳定,并且整个细根呼吸的测定基本控制在细根取出后2 h内完成。根系呼吸速率的计算以根系干重为基础计算每克根系干物质在每秒钟释放的CO2,即比根呼吸速率,比根呼吸计算公式为:比根呼吸 (μgC·g-1·s-1)=细根呼吸 (μgC·s-1)/干重 (g) ×0.006×12。
2.4 数据分析
利用SPSS13.0软件采用单因素方差法分析2个林分同一根序间的细根形态指标和比根呼吸的差异,并分别对序级、林龄以及序级和林龄的交互作用对2个林分细根相关形态指标以及呼吸的影响进行分析;采用回归分析方法分析2个林分细根形态特征及呼吸与序级之间的关系。相关图表用Excel完成。
3 研究结果
3.1 2个林分细根不同序级形态特征
随着序级的升高,2个林分细根直径、根长和组织密度均升高,而比根长则降低,2个林分细根直径4级之间和5级之间具有显著差异 (P<0.05),其余指标间在各级均没有显著差异 (图1)。91年生杉木细根直径的变化从0.550~0.989 mm,19年生杉木从0.570~0.923 mm(图1);细根直径随序级的升高呈有规律的变化,2个林分均可用三次函数表征 (表2)。细根根长的变化91年生从1.054~6.565 cm,19年生从1.491~5.80 cm(图1);根长随序级的升高也具有规律的变化,91年生可用三次函数表征,19年生可用幂函数表征 (表2)。2个林分细根比根长随序级降低,91年生12.382~0.846 m·g-1,19年生8.989~1.144 m·g-1(图1),细根比根长随序级的变化2个林分均可用三次函数表征 (表2)。2个林分细根组织密度91年生变化于0.346~1.581 g·cm-3,19年生变化于0.452~1.321 g·cm-3(图1),组织密度随序级的变化91年生可用指数函数进行表征,19年生可用三次函数表征 (表2)。
林龄仅对细根直径有显著影响 (P<0.01),而对根长、比根长及组织密度的影响均不显著;林龄和序级的交互作用对细根直径和比根长有显著影响 (P<0.05),对根长和组织密度的影响不显著;序级对2个林分细根直径、根长、比根长、组织密度的影响均达到极显著水平 (P<0.01)(表3)。
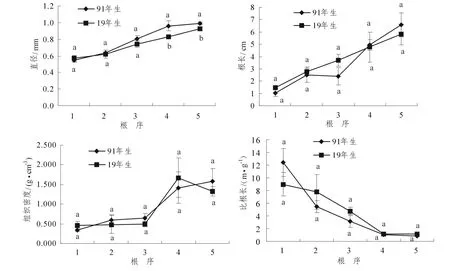
图1 2个林分1~5级根平均直径、平均根长、平均比根长、平均组织密度Figure1 Average diameter,length,specific root length,tissue density of the first five root branch orders for the two forest stands

表3 林龄、序级及林龄与序级的交互作用对细根形态和比根呼吸的影响Table3 Effects of stand age,root order and the interaction between stand age and root order on the fine root morphology and specific root respiration
3.2 2个林分细根不同序级呼吸特征
2个林分细根比根呼吸随序级变化均表现出一致的变化规律,均随序级的升高呈极显著下降(P<0.01)(图2),比根呼吸仅在5级根之间具有显著差异 (P<0.05);91年生杉木林细根比根呼吸变化幅度为 1.269 ~ 0.177 μgC·g-1·s-1,19 年生为 1.297 ~ 0.112 μgC·g-1·s-1,同时,2个林分同一序级比根呼吸之间也均没有显著差异 (图2)。
通过对2个林分细根比根呼吸与序级之间的回归分析发现:比根呼吸均随序级变化呈现出系统性的变化,其中91年杉木林细根比根呼吸随序级升高呈指数函数变化,而19年生比根呼吸可用三次函数表征(表2)。方差分析发现:年龄对细根各序级比根呼吸的影响不显著,序级对2种树种细根比根呼吸的影响均达到极显著水平 (P<0.01);而林龄与序级的交互作用对比根呼吸具有极显著影响 (P<0.01)(表3)。
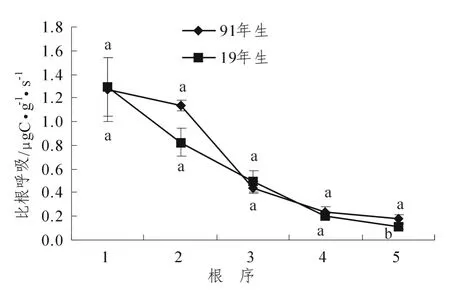
图2 2个林分细根比根呼吸随序级变化情况Figure2 Specific root respiration of the first five root branch orders for the two forest stands
4 讨论
4.1 2个林分细根形态的比较
本研究中2片杉木林细根直径、根长和组织密度均表现出随序级升高而增大,比根长随序级增加而减小的规律。这与Pregitizer等[3]对北美5个针叶树种前3级根构型的研究,Guo等[4]对美国长叶松细根直径、根长和比根长的研究,刘佳等[10]对湖南会同杉木林树种细根直径、比根长的研究也具有类似的结果。同时与以上相关研究中有关阔叶树以及熊德成等[14]有关本区阔叶树的研究具有相似的规律,这也表明木本植物细根在形态特征方面存在一定的异质性。同时本研究中91年生杉木细根除1级根较19年生杉木更大外其余序级均小于19年生,这与Claus& George[15]和Børja等[16]研究发现年轻的林分细根比根长比年老的林分更大相似。有研究表明,细根形态除了受树种本身的遗传因子控制外,主要受土壤资源有效性的影响[17-18];同时,Wang等[13]对东北兴安落叶松和水曲柳的研究发现土壤深度和季节对直径、组织密度都有影响。Comas等[19]对6种温带树种的研究以及Ostonen等[20]对欧洲北方森林3种树种的研究发现,细根的比根长还受到细根生长环境以及纬度位置,气候条件和生物因素影响。然而目前有关林龄对细根序级形态特征的影响如何还有待论证,本研究中从林龄对细根序级形态特征影响的角度出发,分析发现林龄仅对根系直径有显著影响,对其余形态指标影响并不显著,而林龄与序级的交互作用也仅对直径和比根长有显著影响,这在一定程度上表明林龄和序级的交互作用在分析细根序级形态特征时应作为主要的考虑因素。但本研究仅选取2个林龄跨度较大的林分进行研究,要进一步说明林龄对根序特征的影响还需选取较多的年龄序列进行对比研究,以深入揭示这一问题。另外,本研究中细根直径、根长、比根长、组织密度与序级之间均呈现出较好的三次函数,指数函数和幂函数关系,表明序级可以对杉木细根形态特征进行很好的表征。
4.2 2个林分细根呼吸的比较
目前有关细根呼吸的研究已有大量报道,但已有的研究都是以细根直径大小为基础,通过划分不同直径等级的细根来进行,忽视了细根分枝特点对根系功能的作用。本研究发现2个林分细根比根呼吸均随序级增大而降低,也直接表明细根呼吸在不同序级间存在一定的异质性,这与贾淑霞等[11]对落叶松和水曲柳的研究结果相似,也与熊德成等[14]先前对本区6种阔叶树种细根序级呼吸的研究结果相似[14]。同时,与李又芳等[21]、Ryan等[22]、Makita等[23]对根系不同径级呼吸的研究结果发现比根呼吸随着根茎的增大而减小类似。本研究中2个林分细根比根呼吸均可通过与序级的三次函数、指数函数来很好地表达,也进一步说明细根序级结构可以有效表征细根的功能异质性。2个林分比根呼吸随序级增大而降低的机理,可能与众多有关根系呼吸随径级增大而降低及有关根序呼吸随序级升高而降低的机理相似,即随径级或序级的变化,根系的代谢组织减少,导致代谢活动减弱,呼吸作用降低。也有研究表明这与低级根中具有较高的N含量有关,植物细胞中90%的N以蛋白质形式存在,这些蛋白质的存在是细胞发挥正常生理功能的基础[24],这些蛋白质的周转和修复就占到维持呼吸23% ~30%[25]。此外,2个林分之间同一序级细根呼吸仅在5级根之间有显著差异,方差分析发现林龄对比根呼吸并未有显著影响,这可能主要是因为2个林分虽然林龄不同,但是在细根部分根系生理活动并未受到较大林龄的影响,但要进一步说明这一问题同样需要选取较多的年龄序列进行对比研究;而林龄与序级的交互作用对比根呼吸有显著影响,这也表明林龄和序级的交互作用在分析细根序级、比根呼吸时应重点考虑。目前有关不同根序细根呼吸的研究还十分有限,已往细根功能异质性的研究主要通过形态结构[3]、不同根序组织中N浓度[4]或者解剖特征间接进行推测[26-27],同时有关细根呼吸的测定方法仍是极大的难题,尤其是原位测定细根呼吸特别是不同根序的呼吸一直以来没有得到有效的解决。采用离体测定的方法是目前较多采用的用于测定不同序级细根呼吸速率最常见的方法。本研究中所采取的测定方法与本研究组先前对阔叶树细根序级呼吸测定的方法相同,所测得的呼吸同样也只能相对衡量细根序级呼吸的变化规律,而在测定过程中也只能尽量缩短细根从离体到测定的时间,同时保证测定过程中周围环境 (主要是温度和湿度)的相对稳定。因此从精度和长远的角度考虑,探索离体根呼吸有效合理的方法并尝试原位测定细根呼吸的方法仍将是细根研究中重点应该解决的问题。
[1]Ruess R W,Hendrick R L,Burton A J,et al.Coupling fine root dynamics with ecosystem carbon cycling in black spruce forests of interior Alaska[J].Ecological Monographs,2003,73:643-662.
[2]Copley J.Ecology goes underground[J].Nature,2000,406:452-454.
[3]Pregitzer K S,Deforest J L,Burton A J,et al.Fine root architecture of nine North American trees[J].Ecological Monographs,2002,72:293-309.
[4]Guo D L,Mitchell R J,Hendricks J J.Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest[J].Oecologia,2004,140:450-457.
[5]Pregitzer K S,Kubiske M E,Yu C K,et al.Relationships among root branch order,carbon,and nitrogen in four temperate species[J].Oecologia,1997,111:302-308.
[6]Wells C E,Glenn D I,Eissenstat D M.Changes in the risk of fine root mortality with age:A case study in peach,Prunus persica(Rosaceae)[J].American Journal of Botany,2002,89:79-87.
[7]Withington J M,Reich P B,Oleksyn J,et al.Comparisons of structure and life span in roots and leaves among temperate trees[J].Ecological Monographs,2006,76(3):381-397.
[8]Wang Xiang-rong,Gu Jia-cun,Mei Li,et al.Fine root order morphology and proportion between mother roots and daughter roots inFraxiuus mandshuricaandLarix gmeliniiplantations[J].Acta Ecologica Sinica,2006,26(6):1686-1692.[王向荣,谷加存,梅莉,等.水曲柳和落叶松细根形态及母根与子根比例关系 [J].生态学报,2006,26(6):1686-1692.]
[9]Shi Wei,Wang Zheng-quan,Liu Jin-liang,et al.Fine root morphology of twenty hardwood species in Maoershan natural secondary forest in northeastern China[J].Journal of Plant Ecology,2008,32(6):1217-1226.[师伟,王政权,刘金梁,等.帽儿山天然次生林20个阔叶树种细根形态 [J].植物生态学报,2008,32(6):1217-1226.]
[10]Liu Jia,Xiang Wen-hua,Xu Xiao,et al.Analysis of architecture and functions of fine roots of five subtropical tree species in Huitong,Hunan Province,China[J].Journal of Plant Ecology,2010,34(8):938-945.[刘佳,项文化,徐晓,等.湖南会同5个亚热带树种的细根构型及功能特征分析 [J].植物生态学报,2010,34(8):938-945.]
[11]Jia Shu-xia,Zhao Yan-li,Ding Guo-quan,et al.Relationship among fine-root morphology,anatomy,tissue nitrogen concentration and respiration in different branch root orders inLarix gmeliniiandFraxinus mandshurica[J].Bulletin of Botany,2010,45(2):174-181.[贾淑霞,赵妍丽,丁国泉,等.落叶松和水曲柳不同根序细根形态结构、组织氮浓度与根呼吸的关系 [J].植物学报,2010,45(2):174-181.]
[12]Xu Yang,Gu Jia-cun,Dong Xue-yun,et al.Fine root morphology,anatomy and tissue nitrogen and carbon contents of the first five orders in four tropical hardwood species in Hainan Island,China [J].Chinese Journal of Plant Ecology,2011,35(9):955-964.[许旸,谷加存,董雪云,等.海南岛4个热带阔叶树种前5级细根的形态、解剖结构和组织碳氮含量 [J].植物生态学报,2011,35(9):955-964.]
[13]Wang Z Q,Guo D L,Wang X R,et al.Fine root architecture,morphology,and biomass of different branch orders of two Chinese temperate tree species[J].Plant and Soil,2006,288(1/2):155-171.
[14]Xiong De-cheng,Huang Jin-xue,Yang Zhi-jie,et al.Fine root architecture and morphology among different branch orders of six subtropical tree species[J].Acta Ecologica Sinica,2012,32(6):1888-1897.[熊德成,黄锦学,杨智杰,等.亚热带6种树种细根序级结构和形态特征分析[J].生态学报,2012,32(6):1888-1897.]
[15]Claus A,George E.Effect of stand age on fine-root biomass and biomass distribution in three European forest chronosequenc-es[J].Canadian Journal of Forest Research,2005,35:1617-1625.
[16]Børja I,De Wit H A,Steffenrem A,et al.Stand age and fine root biomass,distribution and morphology in a Norway spruce chronosequence in southeast Norway [J].Tree Physiology,2008,28:773-784.
[17]Fitter A H.Characteristics and functions of root systems[M] //Waisel Y,Eshel E,Kafkafi U.Plant roots:The hidden half,2nd edition.New York:Dekker.1996:1-20.
[18]Hodge A.The plastic plant:Root responses to heterogeneous supplies of nutrients[J].New Phytologist,2004,162:9-24.
[19]Comas L H,Eissenstat D M.Patterns in root trait variation among 25 co-existing North American forest species[J].New Phytologist,2009,182(4):919-928.
[20]Ostonen I,Lõhmus K,Helmisaari H S,et al.Fine root morphological adaptations in Scots pine,Norway spruce and silver birch along a latitudinal gradient in boreal forests[J].Tree Physiology,2007,27(11):1627-1634.
[21]Li You-fang,Gao Ren,Li Ying,et al.Root respirations of different diameter classes for Chinese firs in relation to root parameters[J].Journal of Subtropical Resources and Environment,2008,3(2):19-24.[李又芳,高人,李莹,等.不同径级杉木根参数与离体根呼吸[J].亚热带资源与环境学报,2008,3(2):19-24.]
[22]Ryan M G,Hubbard R M,Pongracic S,et al.Foliage,fine-root,woody-tissue and stand respiration in pinus radiata in relation to nitrogen status[J].Tree Physiology,1996,16:333-343.
[23]Makita N,Hirano Y,Dannoura M,et al.Fine root morphological traits determine variation in root respiration ofQuercus serrata[J].Tree Physiology,2009,29(4):579-585.
[24]Lambers H,Chapin F S,Pons T L.Plant Physiological Ecology[M].New York:Springer-Verlag.1998:122-133.
[25]Scheurwater I,Dunnebacke M,Eising R,et al.Respiratory costs and rate of protein turnover in the roots of a fast-growing(Dactylis glanerataL.)and a slow-growing(Festuca ovinaL.)grass species[J].Journal of Experimental Botany,2000,51:1089-1097.
[26]Hishi T,Takeda H.Dynamics of heterorhizic root systems:Protoxylem groups within the fine-system ofChamaecyparis obtuse[J].New Phytologist,2005,167:509-521.
[27]Guo D L,Xia M X,Wei X,et al.Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species[J].New Phytologist,2008,180:673-683.
