啮齿动物捕食和搬运蒙古栎种子对种群更新的影响
2014-08-11张晶虹刘丙万
张晶虹,刘丙万
(东北林业大学野生动物资源学院, 哈尔滨 150040)
啮齿动物捕食和搬运蒙古栎种子对种群更新的影响
张晶虹,刘丙万*
(东北林业大学野生动物资源学院, 哈尔滨 150040)
2010—2011年,在东北林业大学城市林业示范基地研究了啮齿动物对蒙古栎种子的捕食和搬运。结果表明,蒙古栎种子捕食率在年际间无显著差异,但2010年的蒙古栎种子搬运率显著高于2011年,存留率显著低于2011年。啮齿动物在胡桃楸林、樟子松林、水曲柳林和白桦林内对蒙古栎种子的总捕食率和总搬运率分别达到(5.7±13.5)%和(27.1±37.1)%。蒙古栎种子在樟子松林内的捕食率和搬运率均最高,胡桃楸林内蒙古栎种子存留率最高;2010年蒙古栎种子在与蒙古栎林边缘距离0, 10, 20, 30, 40, 50m组间的捕食率无显著差异,搬运率和存留率有显著差异,但2011年蒙古栎种子在不同距离组间的捕食率呈显著差异,搬运率和存留率无显著差异。与蒙古栎林边缘距离20m处蒙古栎种子的捕食率最高。2010年和2011年,分别有(37.8±49.7)%和(27.7±49.8)%的蒙古栎种子被啮齿动物利用,这表明啮齿动物是林业示范基地内蒙古栎地表种子的主要捕食者。冬季食物匮乏秋季贮藏种子是造成啮齿动物对蒙古栎种子有较大捕食、扩散压力的主要原因。因此,啮齿动物对蒙古栎种子的捕食和搬运影响了蒙古栎林的种群更新。
啮齿动物;蒙古栎;捕食;搬运;种群更新
壳斗科(Fagaceae)栎属(Quercus)的植物有着极低的自然更新率。因此很多学者都非常关注栎属植物种群更新的研究。有关壳斗科植物更新的研究国内外已有很多,涉及动物对种子的贮藏、捕食、扩散、多次扩散[1- 4]和种子库和幼苗建成[5- 7]等。但对蒙古栎(Quercusmongolica)种子的捕食和搬运及动物对蒙古栎幼苗更新的作用却很少受到关注。种子扩散是保证植物种群繁衍增殖的基本途径[8],而动物是植物自然更新的重要参与者[9]。如林木种子是森林内啮齿动物赖以生存的重要食物来源[10],它们在消耗大量种子的同时,对植物或植物种子本身会产生有益的影响[11]。许多啮齿动物分散贮藏植物种子,促进种子的进一步扩散(即次级扩散)[12- 13],扩大了植物的分布范围,减少母树下种子密度,从而减少植物种内竞争。被分散贮藏的种子也降低与其他捕食者遭遇的风险[14]。此外,埋藏在土壤基质中的种子,可能由于温湿度适宜,微环境有利,并最终逃脱动物的捕食而萌发成苗,促进植物更新。因此,啮齿动物对植物种群更新和扩散具有重要意义[15]。
蒙古栎也称柞树,隶于壳斗科栎属,是我国北方林区的主要阔叶树种之一,具有较强的繁殖能力。栎类因种子较大、且富含淀粉等营养物质而成为昆虫和鸟兽等动物的重要食物来源[16]。前人相关研究指出,尽管蒙古栎每年都有种子散落,但林下多为1年生幼苗,多年生幼苗极少,种子库也极为短暂[17]。而动物捕食是地表种子损失的主要原因[18- 20]。自然界中,鸦类(Corvidae)和啮齿动物能分散贮食[21]。据观察及相关研究表明,林业示范基地内主要的鸟类有大斑啄木鸟(Dendrocoposmajor),灰喜鹊(Cyanopicacyana)等,并没有松鸦(Garrulusglandarius)的存在[22]。因此,本研究通过比较不同林分类型,不同距离组间啮齿动物对蒙古栎种子的捕食和搬运的格局,测定啮齿动物捕食是影响蒙古栎种子命运的主要因子,并探讨啮齿动物捕食和搬运对蒙古栎种群的潜在作用。
1 研究方法
1.1 研究地点概况
东北林业大学城市林业示范基地,位于黑龙江省哈尔滨市,N45°20′—45°25′,E127°30′—127°34′。海拔136—140m,气候属于中温带大陆性季风气候,冬长夏短,全年平均降水量569.1mm,四季分明,年平均温度3.6℃,≥10℃年积温2757℃。20世纪50年代末到60年代初造人工纯林。示范基地内现有30块人工纯林,蒙古栎林面积约为0.28 hm2(40m×70m),其他林型面积约为0.5hm2(70m×70m)。基地内分布的树种除蒙古栎外,还包括樟子松(Pinussylvestnis)、白桦(Betulaplatyphylla)、水曲柳(Fraxinusmandshurica)、胡桃楸(Juglansmandshurica)等,胡桃楸林、樟子松林、水曲柳林和白桦林与蒙古栎林相邻,且在上述4个林分中分布有野生的蒙古栎幼苗。
蒙古栎的花期是4—5月,果期在9月,种子成熟后会遭到啮齿动物等的捕食和搬运。在研究地区,分布的主要啮齿动物有松鼠(Sciurusvulgaris)、花鼠(Eutamiassibiricus)、褐家鼠(Rattusnorvegicus)、黑线姬鼠(Apodemusagrarius)等常见种。据观察,这些啮齿动物均可捕食蒙古栎种子,而贮藏种子的啮齿动物(如松鼠)也可通过分散贮藏来扩散蒙古栎种子[22]。
1.2 研究方法
以花生为饵,采用笼捕法调查啮齿动物种类和数量。在胡桃楸林、樟子松林、水曲柳林和白桦林内分别设置3条平行样线,样线间距为13m,每条样线置放5个鼠笼,笼间距为13m,次日检查进笼情况,连续设笼3d。捕获的个体记录其种类,用(捕获的啮齿动物个体数量/笼日数)×100%来表示捕获率,并以捕获率表示种群相对密度。在2010年和2011年的8、9、10月份,在蒙古栎林、胡桃楸林、樟子松林、水曲柳林和白桦林内每周定期进行一次啮齿动物行为观察,每次不少于2h,选择啮齿动物日活动高峰时段,如9:00—11:00,14:00—16:00等,记录啮齿动物种类、数量和行为等。
2010年和2011年8月中旬种子雨开始下落时起,在蒙古栎林内设置6×3,共18个1 m×1 m的样方收集框(图1),每1—2d调查1次,直到9月中旬没有种子降落,记录各样方收集框内蒙古栎种子壳斗数和蒙古栎种子数,以此估算蒙古栎种子产量。
在蒙古栎种子成熟的季节,从地面或母树上收集若干蒙古栎种子,从中选取饱满的种子用于研究啮齿动物对蒙古栎种子命运的影响。以蒙古栎林4边中点为准,向胡桃楸林、樟子松林、水曲柳林、白桦林4个林分内延伸4条垂直平行线,以各林分边缘为起始点,每条样线分别设立间距为10m 的6个样点,每一个样点作为一个距离组,分别设为0, 10, 20, 30, 40, 50m(图1)。分别于2010年和2011年的9月21日至9月30日,共10d,每天6:30在各距离组放置10粒饱满未萌发的蒙古栎种子,并于当日15:30检查,记录蒙古栎种子被捕食(predation)、被搬运(removal)和存留(remaining)的格局。
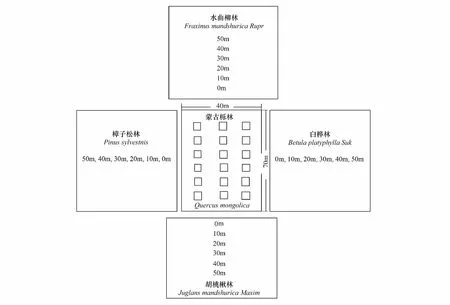
图1 蒙古栎种子产量及命运研究样方设置示意图Fig.1 Sketch map of plots setting of Mongolian oak seed corps and fates胡桃楸林,樟子松林,水曲柳林和白桦林与蒙古栎林间分别有一条宽5m, 4m, 8m, 6m的小路 There is a width of 5m, 4m, 8m and 6m path between Manchurian walnut and Mongolian oak, scotch pine and Mongolian oak, northeast China ash and Mongolian oak, white birch and Mongolian oak
采用SPSS17.0软件包作统计分析,运用Chi-Square检验分析各林分内啮齿动物种类和数量是否存在显著差异。运用配对T检验分析2010年和2011年蒙古栎种子产量差异。应用一般线性分析的Two-way ANOVA检验各林分组和各距离组释放点蒙古栎种子被捕食、搬运和存留情况的差异及二者的交互作用,同时比较蒙古栎种子产量差异对种子命运是否有显著影响。
2 研究结果
2.1 啮齿动物的种类及捕获率
调查期间共捕获啮齿动物4种,15只,总捕获率为8.3%(表 1)。4种林分类型中,啮齿动物的捕获率以水曲柳林内最高,其次是白桦林和胡桃楸林,樟子松林内没有捕获啮齿动物(表1)。Chi-Square检验表明,胡桃楸林、水曲柳林、白桦林啮齿动物捕获率之间无显著差异(X2=0.500,df=2,P>0.05)。但林型间的鼠种组成存在差异:黑线姬鼠是白桦林的优势种;而褐家鼠是水曲柳林的优势种。
2010年多在樟子松林、水曲柳林和胡桃楸林内观察到松鼠和花鼠觅食,平均每次4只左右,且各观察过1次褐家鼠和黑线姬鼠搬运蒙古栎种子;而在2011年则很少观察到啮齿动物活动,仅在水曲柳林内观察到1次松鼠和1次花鼠觅食。
表1 2011年4种林分内捕获啮齿动物的种类数量及捕获率
Table 1 Species and trap success in the four stands: Manchurian walnut, scotch pine, northeast China ash and white birch in 2011 (n=180 trap nights)

种类Species胡桃楸林Juglansmandshurica樟子松林Pinussylvestnis水曲柳林Fraxinusmandshurica白桦林Betulaplatyphylla黑线姬鼠Apodemusagrarius2—24褐家鼠Rattusnorvegicus1—31松鼠Sciurusvulgaris——1—花鼠Eutamiassibiricus——1—捕获率Trapsuccess6.7—15.611.1
2.2 蒙古栎种子产量及其对种子命运情况的影响
2010年壳斗数(t=8.380,df=17,P<0.05)和种子产量(t=8.126,df=17,P<0.05)均显著高于2011年。2010年蒙古栎种子产量较高,壳斗数和种子数分别达到(55.6±26.0)个/m2和(39.8±19.4)个/m2,而2011年壳斗数和种子数仅为(4.6±1.9)个/m2和(3.4±1.3)个/m2。蒙古栎种子捕食率在年间无显著差异(F1,478=1.103,P>0.05);但2010年蒙古栎种子搬运率(F1,478=11.558,P<0.05)显著高于2011年,存留率(F1,478=7.178,P<0.05)显著低于2011年(表2)。2010年和2011年,分别有(37.8±49.7)%和(27.7±49.8)%的蒙古栎种子被啮齿动物利用,说明啮齿动物是蒙古栎种子主要的捕食群体。

表2 2010年和2011年蒙古栎种子命运的比较
2.3 释放点蒙古栎种子的命运
由于啮齿动物捕食和搬运,两年中蒙古栎种子在胡桃楸林、樟子松林、水曲柳林和白桦林内的总捕食率和总搬运率分别达到(5.7±13.5)%和(27.1±37.1)%。蒙古栎种子在释放点处的命运包括原地取食和原地存留。2010年种子在各林分内的捕食率(F3,236=8.417,P<0.05),搬运率(F3,236=24.300,P<0.05)和存留率(F3,236=26.849,P<0.05)均呈显著差异。2011年蒙古栎种子在各林分内的捕食率(F3,236=4.866,P<0.05),搬运率(F3,236=6.040,P<0.05)和存留率(F3,236=6.877,P<0.05)也均呈显著性差异(图2)。两年的结果表明,蒙古栎种子在樟子松林的捕食率和搬运率最高,胡桃楸林内蒙古栎种子存留率最高。
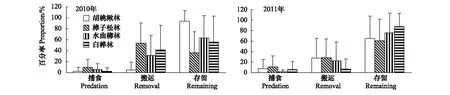
图2 4种林分蒙古栎种子的命运Fig.2 Fates of Mongolian oak seeds in four stands: Manchurian walnut, scotch pine, northeast China ash and white birch
2010年蒙古栎种子在不同距离组间的捕食率(F5,234=1.863,P>0.05)无显著差异,搬运率 (F5,234=4.823,P<0.05)和存留率(F5,234=5.188,P<0.05)呈显著性差异;2011年蒙古栎种子在不同距离组间的捕食率(F5,234=3.173,P<0.05)呈显著差异,而搬运率 (F5,234=0.738,P>0.05)和存留率 (F5,234=0.455,P>0.05)均无显著差异(图3)。两年的实验结果表明,20m处蒙古栎种子的捕食率最高,40m处的搬运率最高,0m处的存留率最高。
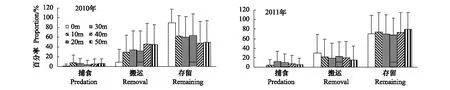
图3 不同距离组蒙古栎种子命运的比较Fig.3 Fates of Mongolian oak seeds among different distance groups:0m, 10m, 20m, 30m, 40m and 50m
2010年和2011年,种子的搬运率和存留率在林分与距离组间的交互作用均呈显著差异(2010:F15,216=3.834,P<0.05,F15,216=3.395,P<0.05;2011:F15,216=3.608,P<0.05,F15,216=3.136,P<0.05),2010年种子捕食率在林分与距离组间的交互作用呈显著差异(F15,216=2.462,P<0.05),而2011年无显著差异(F15,216=1.258,P>0.05)。2010年,樟子松林10m距离组的捕食率(平均22%)最高,白桦林40m距离组的搬运率最高(平均81%),在胡桃楸林0,20m和白桦林0m距离组的存留率最高(100%);2011年胡桃楸林20m距离组的捕食率最高(平均23%),10m距离组的搬运率最高(平均60%),40m距离组的存留率最高(平均100%)。
3 讨论
3.1 种子产量与蒙古栎种子命运的关系
壳斗科植物的种子产量在年间有很大的变化,种子产量的丰年一般每隔两至数年发生一次[6,23- 29]。种子雨产量的大幅变动,可能有助于部分种子逃脱动物的捕食[4]。两年内,蒙古栎种子产量并不稳定。2010年种子产量较高,饱满完好的种子占主要部分,虫蛀种子、霉变种子和败育种子比例低,啮齿动物活动频繁,使得2010年种子搬运率显著高于2011年,种子在更大空间的分散贮藏,可能导致了2011年蒙古栎幼苗数量的大幅度上升,促进了蒙古栎种群的更新;而2011年蒙古栎种子产量极低,虫蛀和霉变的蒙古栎种子比例也较高,被啮齿动物捕食和搬运的种子数量减少,导致2011年蒙古栎种子存留率显著高于2010年。蒙古栎种子壳斗数多于种子数,且在调查过程中对种实残留物及种皮碎片外观形状的观察,可以确定,部分蒙古栎种子被啮齿动物和昆虫捕食。种子产量的年间变化,同时也影响着地面和林冠层中种子捕食者的数量。例如,在2010年和2011年同一时期进行的啮齿动物行为观察中发现,2010年期间,松鼠、花鼠多在樟子松林和水曲柳林中活动觅食,每次可观察到2—3只,多则达5只,在蒙古栎林,胡桃楸林和白桦林中也偶尔见到,且各观察过一次褐家鼠和黑线姬鼠搬运蒙古栎种子;而在2011年研究期间,只在水曲柳林中看见两次啮齿动物,分别是松鼠和花鼠,且各只有1只,这可能是2011年蒙古栎种子捕食和搬运率低于2010年的原因之一。
3.2 啮齿动物对蒙古栎种子命运及种群更新的作用
存活的种子或处于休眠状态,或发芽,或衰老后死亡,或被动物取食、搬运等,它们的不同命运对种群更新和群落演替非常重要[30]。啮齿动物通常会取食较大的种子或果实,因其富含营养物质,且种子或果实体积大小适宜,易于贮藏。蒙古栎种子没有休眠特性,种子库短暂而断续,又因为橡实中含丰富营养,是多种动物取食的对象[31]。通过对被捕食过的蒙古栎种子残片的观察得知,取食蒙古栎种子的动物主要是啮齿动物。在调查地区的4类林分中,啮齿动物对蒙古栎种子均有较大的捕食压力,因为在2010年和2011年,分别有37.8%和27.8%的蒙古栎种子被啮齿动物取食或搬运到它处取食或贮藏,这表明啮齿动物是蒙古栎种子扩散的媒介,并且主要依靠松鼠和花鼠等的捕食和搬运来传播其种子。在秋季,蒙古栎是研究地内主要的结种子树种,种子产量和可供动物利用的种子都极为有限,且由于面临冬季,食物匮乏而使种子有较大被捕食和被扩散的压力。其他研究也表明,食物资源丰富使种子存活增加,并降低捕食压力,而食物资源匮乏则增加动物对种子的捕食压力[32- 35]。在本研究的4种林分类型中,樟子松林内啮齿动物对蒙古栎种子有着更大的捕食压力,因为在两年的调查中,由于松鼠的主要活动,使得樟子松林内的蒙古栎种子拥有最高的捕食率和搬运率,而存留率最低。蒙古栎种子在4个林分内的平均存留率有显著差异,可能与两个因素有关,一是4个林分内啮齿动物组成有差异,二是林下灌丛的盖度差异。实验是下午检查释放的种子,松鼠和花鼠是昼行性动物[36],且多在樟子松林内活动,偶见在胡桃楸林和水曲柳林,松鼠贮食量大,所以种子在樟子松林内的利用率最高。其他啮齿动物虽然是昼夜活动型,但以夜间活动为主,导致其他林分内种子利用率较低。樟子松林下的灌丛层次鲜明,以蒙古栎幼苗,樟子松幼苗居多,且由于林冠层有樟子松种子存在,更易吸引松鼠和花鼠较大啮齿动物前来觅食;胡桃楸林下灌木层稀少,偶见裸地,啮齿动物活动较少;而水曲柳林和白桦林下的灌木层最茂密,食物不宜被发现。两年的调查结果还表明,蒙古栎种子在0m存留率最高,可能是因为该距离点位于两林分类型之间,人为干扰最严重,影响了啮齿动物正常觅食。而20m处捕食率最高,可视为比较安全的捕食地带;40m处种子的搬运率最高,也进一步证明了啮齿动物可能是蒙古栎种子扩散的媒介。由于各林分生境类型及林分内啮齿动物组成不同,使得林分与距离组间的交互作用呈现显著差异。
综上所述,种子的潜在价值决定动物是否取食、搬运和贮藏[14]。动物一方面在种子密度较大的资源处(如母树附近或距离组附近等)快速消耗部分种子以满足当前需要,另一方面将剩余的大量可利用种子收集起来搬运到贮藏点以备所需,以致种子雨结束时,仅有极少数散落在母树下的种子得到保留。与此同时,啮齿动物在选择种子时会挑选完好饱满的种子,种子一经被扩散,可能由于微环境适宜并最终逃脱动物的捕食而发芽生长成苗,在研究地区的四个林分内,均有蒙古栎幼苗和成树散布,这也就解释了为什么蒙古栎林下蒙古栎幼苗数量较少,而其他林分内又有较多蒙古栎幼苗。因而证实了啮齿动物是蒙古栎地表种子的主要捕食者,是影响蒙古栎种子命运的主要因子,它们对蒙古栎种子的捕食和搬运可对野生蒙古栎种群的更新产生重要的影响。
致谢:中国科学院动物研究所肖治术副研究员提供部分相关文献和实验指导,特此致谢。
[1] Vander Wall S B. Masting in animal-dispersed pines facilitates seed dispersal. Ecology, 2002, 83(12): 3508- 3516.
[2] Vander Wall S B, Kuhn K M, Beck M J. Seed removal, seed predation, and secondary dispersal. Ecology, 2005, 86(3): 801- 806.
[3] Xiao Z S, Zhang Z B, Wang Y S, Cheng J R. Acorn predation and removal ofQuercusserratain a shrubland in Dujiangyan Region, China. Acta Zoologica Sinica, 2004, 50(4): 535- 540.
[4] Xiao Z S, Zhang Z B. Hoarding behavior of rodents and plant seed dispersal. Acta Theriologica Sinica, 2004, 24(1): 61- 70.
[5] Xiao Z S, Wang Y S, Zhang Z B. Seed bank and the factors influencing it for three Fagaceae species in Dujiangyan Region, Sichuan. Biodiversity Science, 2001, 9(4): 373- 381.
[6] Tanouchi H, Sato T, Takeshita K. Comparative studies on acorn and seedling dynamics of fourQuercusspecies in evergreen broad-leaved forest. Journal of Plant Research, 1994, 107(6): 153- 159.
[7] Li H J, Zhang Z B. Relationship between animals and plant regeneration by seed Ⅱ. Seed predation, dispersal and burial by animals and relationship between animals and seedling establishment. Biodiversity Science, 2001, 9(1): 25- 37.
[8] Howe H F, Smallwood J. Ecology of seed dispersal. Annual Review of Ecology and Systematics, 1982, 13: 201- 228.
[9] Li X S, Peng M C. Research progress on natural regeneration of plants. Chinese Journal of Ecology, 2007, 26(12): 2081- 2088.
[10] Xiao Z S, Zhang Z B. Small mammals consuming tree seeds in Dujiangyan forest. Acta Theriologica Sinica, 2004, 24(2): 121- 124.
[11] Xiao Z S, Zhang Z B, Wang Y S. Observations on tree seed selection and caching by Edward′s long-tailed rat (Leopoldamysedwardsi). Acta Theriologica Sinica, 2003, 23(3): 208- 213.
[12] Vander Wall S B, Longland W S. Diplochory: are two seed dispersers better than one. Trends in Ecology and Evolution, 2004, 19: 155- 161.
[13] Vander Wall S B, Kuhn K M, Gworek J R. Two-phase seed dispersal: linking the effects of frugivorous birds and seed-caching rodents. Oecologia, 2005, 145: 282- 287.
[14] Vander Wall S B. Food hoarding in animals. Chicago: University of Chicago Press, 1990.
[15] Kollmann J, Schill H P. Spatial patterns of dispersal, seed predation and germination during colonization of abandoned grassland byQuercuspetreaandCorylusavellana. Vegetatio, 1996, 125: 193- 205.
[16] Wang X, Xiao Z S, Zhang Z B, Pan H C. Insect seed predation and its relationships with seed crop and seed size ofQuercusmongolica. Acta Entomologica Sinica, 2008, 51(2): 161- 165.
[17] Chen B H, Feng Z W, Ju X. Oak forest in the southern slope of Xiao Hinggan Ling. Forestry bulletins, 1959, (4): 61- 86.
[18] Fei S M, Peng Z H, Yang D S, Zhou J X, He Y P, Wang P, Chen X M, Jiang J M. Seed rain and seed bank ofCastanopsisdelavayipopulations in mountainous area of southwest Sichuan. Scientia Silvae Sinicae, 2006, 42(2): 49- 55.
[19] Liu T, Zhou Z Q. Seed rain and soil seed bank ofQuercusmongolicapopulations. Journal of Northeast Forestry University, 2007, 35(5), 22- 23.
[20] Pan C L, Lai J Y, Li X D, Shi H M. Seed rain and natural regeneration ofKmeriaseptentrionalisin Mulun of Guangxi. Chinese Journal of Ecology, 2008, 27, 2235-2239.
[21] Lu C H. Seed dispersal-the role of animal. Journal of Northeast Forestry University, 2003, 70- 88.
[22] Hao H J, Wu J P. The role ofSquirrelin the Dispersal ofMongolianOakin the Northeast Forestry University Forestry Farm in Harbin. Territory and natural resources study, 2012, 4: 84- 85.
[23] Shaw M W. Factors affecting the natural regeneration of sessile oak (Quercuspetraea) in North Wales: 1. A preliminary study of acorn production, viability and losses. Journal of Ecology, 1968, 56(3): 565- 583.
[24] Sork V L, Bramble J, Sexton O. Ecology of mast-fruiting in three species of north American Deciduous Oaks. Ecology, 1993, 74(2): 528- 541.
[25] Sork V L. Evolutionary ecology of mast-seeding in temperate and tropical oaks (Quercusspp.). Vegetatio, 1993, 107/108: 133- 147.
[26] Koenig W D, Mumme R L, Carmen W J, Stanback M T. Acorn production by oaks in central coastal California: variation within and among years. Ecology, 1994, 75(1): 99- 109.
[27] Kelly D. The evolutionary ecology of mast seeding. Trends in Ecology and Evolution, 1994, 9(12): 466- 470.
[28] Harmer R. Natural regeneration of broad-leaved trees in Britain: Ⅱ. Seed production and predation. Forestry, 1994, 67(4): 275- 286.
[29] Crawley M J, Long C R. Alternate bearing, predator satiation and seedling recruitment inQuercusroburL. Journal of Ecology, 1995, 83(4): 683- 696.
[30] Harper J L. Population biology of plants. London: Academic press, 1977, 83- 147, 458- 482.
[31] Sun S C, Chen L Z. The effects of animal removal and groundcover on the fate of seeds ofQuercusliaotungensis. Acta Ecologica Sinica, 2001, 21(1): 80- 85.
[32] Lu J Q, Zhang Z B. Effects of high and low shrubs on acorn hoarding and dispersal of Liaodong oakQuercusliaotungensisby small rodents. Acta Zoologica Sinica, 2005, 51: 195- 204.
[33] Xiao Z S, Zhang Z B, Wang Y S. The effects of seed abundance on seed predation and dispersal by rodents inCastanopsisfargesii(Fagaceae). Plant Ecology, 2005, 177: 249- 257.
[34] Xiao Z S, Wang Y S, Harris M K, Zhang Z B. Spatial and temporal variation of seed predation and removal of sympatric large-seeded species in relation to innate seed traits in a subtropical forest, Southwest China. Forest Ecology and Management, 2006, 222: 46- 54.
[35] Xiao Z S, Zhang Z B, Wang Y S. Impacts of scatter-hoarding rodents on restoration of oil teaCamelliaoleiferain a fragmented forest. Forest Ecology and Management, 2004, 196: 405- 412.
[36] Su H J, Ma J Z, Zong C. Behavior Comparison between four diurnal Animals on Seed Consumption and Hoarding of Korean Pine. Chinese Journal of Zoology, 2001, 42(2): 10- 16.
参考文献:
[3] 肖治术, 张知彬, 王玉山, 程瑾瑞. 都江堰地区灌丛内枹栎橡子的捕食和搬运. 动物学报, 2004, 50(4): 535- 540.
[4] 肖治术, 张知彬. 啮齿动物的贮藏行为与植物种子的扩散. 兽类学报, 2004, 24(1): 61- 70.
[5] 肖治术, 王玉山, 张知彬. 都江堰地区三种壳斗科植物的种子库及其影响因素研究. 生物多样性, 2001, 9(4): 373- 381.
[7] 李宏俊, 张知彬. 动物与植物种子更新的关系 Ⅱ. 动物对种子的捕食、扩散、贮藏及与幼苗建成的关系. 生物多样性, 2001, 9(1): 25- 37.
[9] 李小双, 彭明春, 党承林. 植物自然更新研究进展. 生态学杂志, 2007, 26(12): 2081- 2088.
[10] 肖治术, 张知彬. 都江堰林区小型兽类取食林木种子的调查. 兽类学报, 2004, 24(2): 121- 124.
[11] 肖治术, 张知彬, 王玉山. 小泡巨鼠对森林种子选择和贮藏的观察. 兽类学报, 2003, 23(3): 208- 213.
[16] 王学,肖治术,张知彬,潘红春. 昆虫种子捕食与蒙古栎种子产量和种子大小的关系. 昆虫学报, 2008, 2, 51(2): 161- 165.
[17] 陈炳浩,冯宗炜,鞠岘. 小兴安岭南坡的柞林. 林业集刊. 1959, (4): 61- 86.
[18] 费世民, 彭镇华, 杨冬生, 周金星, 何亚平, 王鹏, 陈秀明, 蒋俊明. 四川西南山区高山锥种群的种子雨和种子库. 林业科学, 2006, 42(2), 49- 55.
[19] 刘彤, 周志强. 蒙古栎种群种子雨和土壤种子库. 东北林业大学学报, 2007, 35(5), 22- 23.
[20] 潘春柳, 赖家业, 黎向东, 石海明. 广西木论地区单性木兰种子雨和自然再生. 生态学杂志, 2008, 27, 2235- 2239.
[21] 鲁长虎. 种子传播—动物的作用. 东北林业大学出版社, 2003, 70- 88.
[22] 郝宏娟, 吴建平. 东北林业大学实验林场蒙古栎扩散与松鼠的关系. 国土与自然资源研究, 2012, 4: 84- 85.
[31] 孙书存, 陈灵芝. 动物搬运与地表覆盖物对辽东栎种子命运的影响. 生态学报, 2001, 21(1): 80- 85.
[32] 路纪琪, 张知彬. 高低灌丛内小型啮齿动物对辽东栎贮藏和扩散的影响. 动物学报, 2005, 51: 195- 204.
[36] 粟海军, 马建章, 宗诚. 四种昼行性动物取食和贮藏红松种子的行为比较. 动物学杂志, 2007, 42(2): 10- 16.
Patterns of seed predation and removal of Mongolian oak (Quercusmongolica) by rodents
ZHANG Jinghong, LIU Bingwan*
CollegeofWildlifeResources,NortheastForestryUniversity,Harbin150040,China
During 2010 and 2011, seed predation and removal of Mongolian oak (Quercusmongolica) were studied in the City forestry demonstration base of Northeast Forestry University, Harbin, Heilongjiang Province. The results showed that: there was no significant difference in Mongolian oak seed predation rate among the two years. But Mongolian oak seed removal rate in 2010 was significantly higher than 2011, and the remaining rate was significantly lower than that of 2011. The total seed predation rate and removal rate of Mongolian oak by rodents respectively were (5.7±13.5)% and (27.1±37.1)%. Mongolian oak seed predation rate and removal rate were the highest in scotch pine forest because of predation and removal. Seed remaining rate was the highest in Manchurian walnut forest. Mongolian oak seed predation rate had not significantly different in 0m, 10m, 20m, 30m, 40m, 50m distance group that were apart from edge of Mongolian oak forest, but removal rate and remaining rate had significant difference in 2010. Seed predation rate had significant difference at different distance group, removal rate and remaining rate had not significantly different in 2011. Mongolian oak seed predation rate was the highest at 20m. There were (37.8±49.7)% and (27.7±49.8)% Mongolian oak seed were used by rodents in 2010 and 2011, respectively. This showed that the rodents were the main ground seeds predators of the City forestry demonstration base, winter food shortages and autumn storage seeds could be the main reason that caused Mongolian oak seed have large predation and spread stress by rodents. Therefore, seed predation and removal of Mongolian oak by rodents could affect the fates of Mongolian oak seeds and population regeneration.
rodents; mongolian oak (Quercusmongolica); predation; removal; population regeneration
中央高校基本科研业务费专项资金(DL13CA08); 哈尔滨市科学技术局科技创新人才研究专项资金资助项目(RC2009QN002149)
2012- 10- 12;
2013- 03- 04
10.5846/stxb201210121412
*通讯作者Corresponding author.E-mail: liubw1@sina.com
张晶虹,刘丙万.啮齿动物捕食和搬运蒙古栎种子对种群更新的影响.生态学报,2014,34(5):1205- 1211.Zhang J H, Liu B W.Patterns of seed predation and removal of Mongolian oak (Quercusmongolica) by rodents.Acta Ecologica Sinica,2014,34(5):1205- 1211.
