绩溪光皮桦次生林的种群动态1)
2014-08-02万志兵程国源丁强强方乐金
万志兵 程国源 丁强强 方乐金
(黄山学院,黄山,245041) (安徽绩溪县镇头林场) (黄山学院)
绩溪光皮桦次生林的种群动态1)
万志兵 程国源 丁强强 方乐金
(黄山学院,黄山,245041) (安徽绩溪县镇头林场) (黄山学院)
对绩溪光皮桦次生林种群进行了调查,以种群生命表及生存分析理论为基础,以林木径级结构代表龄级结构,编制了绩溪光皮桦种群静态生命表,分析绩溪光皮桦次生林种群结构的动态变化特点。结果表明:光皮桦种群结构呈纺锤型,幼龄个体数量偏少,中龄个体数居多,而老龄个体则相对数量较少,说明种群有衰退的趋势;在第Ⅴ龄级时该种群出现死亡高峰,其种群存活曲线属 Deevey-Ⅱ型;绩溪光皮桦次生林种群具有前期种群数量少,中期稳定,末期衰退的特点。
光皮桦;龄级;静态生命表;种群结构;种群动态
光皮桦(BetulaluminiferaH. winkl.)为落叶高大乔木,又名亮皮桦、亮叶桦,俗称山桃,属桦木科(Betulaceae)桦木属(BetulaL.)落叶乔木[1-2],是中国特有的珍贵经济树种[3]。光皮桦分布广泛、生长快、材质优良、用途广泛,具有很高的经济价值和开发前景[2,4]。光皮桦在秦岭、淮河流域以南的10多个省(自治区)均有天然分布[1],分布范围广、跨度大。光皮桦地理分布区域正是我国最大的亚热带阔叶林区,地带性植被以常绿阔叶林为主。现存的光皮桦林分均以壳斗科为基本建群种,次生性强。在原生森林类型中光皮桦重要值很小,只是在原生类型遭到破坏后的演替过程中,光皮桦取而代之,重要值增高,成为亚热带主要落叶阔叶林类型之一[3]。光皮桦群落主要由高位芽植物组成,以中型叶为主,与亚热带林区植物的研究基本一致;群落内部结构层次明显,光皮桦位于乔木层的第一层,各层内植物种类、数量依各地自然条件及人为干扰强度而各有不同。光皮桦与优势伴生树种相比竞争系数较大,林下幼苗逐渐不见,仅留成熟大树和其它侵入树种所致[5-6]。光皮桦林分优势树种间联结程度低、不显著,因为光皮桦群落物种丰富、成分复杂且群落结构不完善不稳定,处于变化和演替中[7-10]。目前主要对广西、福建闽北的光皮桦群落开展了种群生态学研究,研究认为人为干扰破坏了光皮桦的天然更新,干扰较轻的光皮桦群落的树木物种多样性指数及重要值高于人为干扰较重的群落[7,11]。本研究以绩溪光皮桦种群为对象,以种群生命表及生存分析理论为基础,通过编制静态生命表,对种群生存函数、种群的动态和年龄结构进行分析,期望了解绩溪光皮桦种群的自然演替情况和动态变化规律,为光皮桦的保护和合理利用提供参考依据。
1 研究区概况
研究区位于绩溪县,三十八桥村,地处东经118°46′944″~118°46′782″,北纬30°17′120″~30°17′89″,海拔645~739 m,系黄山山脉向东北延伸之支脉,皖南山区变层岩系的中山、低山地区。该地区气候类型属于亚热带湿润类型,年平均气温15.9 ℃,无霜期220~256 d。年平均降水量1 412 mm,但分配不均,夏秋多,冬春少,夏季常呈现暴雨,冬季旱冷,雨水较少。土壤为黄红壤,坡向为阴坡,坡度为缓坡。乔木层为常绿阔叶林,主要有光皮桦、枫香(Liquidambarformosana)、甜槠(Castanopsiseyrei)、杉木(Cunninghamialanceolata)、青冈(Cyclobalanopsisglaica)、苦槠(Castanopsissclerophylla)、马尾松(Pinusmassoniana)等;灌木层植物有小构树(Broussonetiakazinoki)、山牡荆(Vitexquinata)、乌药(Linderaaggregata)、檵木(Loropetalumchinense)、山胡椒(Linderaglauca)等;草本层以千根草(Euphorbiathymiflia)、乌蕨(Stenolomachusanum)等为主。
2 研究方法
2012年11月,在绩溪县三十八桥村光皮桦次生林中,选择光皮桦分布集中,长势较好的地段设置200 m×100 m的样地,该样地的坡度为20°~30°。调查区域林龄为中龄林,郁闭度为0.8。对样地内光皮桦进行全林每木调查,调查指标为胸径及树高。
2.1 龄级结构
有关种群大小级别的划分标准,不同的学者提出了不同的标准[12-15]。在光皮桦年龄结构的分析中,将殷东生等[16-17]的研究成果作为参考标准,并结合光皮桦生活的特点,采用“空间代替时间”法[18],即用树木的径级结构代替年龄结构。以胸径大小为标准将样地内的光皮桦分为6个径级,每径级的间隔距离为5 cm。具体分类标准为:3 cm<胸径≤5 cm为第1径级;5 cm<胸径≤10 cm为第2径级;10 cm<胸径≤15 cm为第3径级,以此类推;然后将第1径级对应第I龄级,第2径级对应第Ⅱ龄级,……如此类推,一一对应,统计各个龄级的植株数量,编制出绩溪光皮桦种群静态生命表,分析研究其动态变化规律。
采用陈晓德[19]的数学模型,推导种群年龄结构的动态指数对绩溪光皮桦种群动态进行定量描述,其计算方法为:

式中:Vn为种群从n级到n+1级的个体数量变化情况;Vpi为整个种群结构的数量变化动态指数;Sn为第n个龄级种群个体数;Sn+1为第n+1个龄级种群个体数。
当考虑未来的外部干扰时,
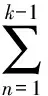
式中:K为种群龄级数量。Vpi与Vn的数值取正、负、零的意义分别反映了种群或相邻龄级个体数量的增长、衰退及稳定的动态关系。
2.2 静态生命表
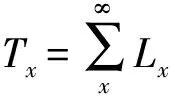
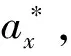
2.3 生存函数
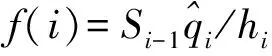
3 结果与分析
3.1 龄级结构
种群结构分析是揭示种群生存现状和更新策略的重要途径之一[25]。对样地内光皮桦的个体数量进行统计,分析该地点的光皮桦种群龄级结构(见表1)。调查样地光皮桦共455株,平均树高12.3 m,平均胸径11.9 cm,最大胸径28.1 cm,最小胸径3.4 cm。由表1可看出,光皮桦在每个龄级都有一定数量的分布,种群龄级结构属纺锤形,表现为衰退型种群。第I龄级个体数是19株,占该群体总数的4.2%,在幼龄阶段个体出现一定程度缺失,可能与种群本身的生理特性有关。第Ⅱ龄级和第III龄级个体数占该群体总数的73.4%,随着径级的增大,个体数量呈下降趋势。老龄个体相对数量较少,第VI龄级个体数只有1株,有可能经受过较严酷的环境选择(自然或人为干扰),导致数量减少。分析数据表明:光皮桦种群的幼龄阶段发展不稳定,幼树数量相对较少。

表1 光皮桦种群龄级结构
根据量化分析方法,绩溪光皮桦种群相邻各级间个体数量变化动态为:V1=-86.986%,V2=-22.340%,V3=141.026%,V4=70.513%,V5=95.652%。该种群龄级结构的动态指数Vpi=64.58%;考虑到种群的外部干扰时,Vpi=10.76%>0,表明现阶段该种群较为稳定。在调查中发现,不少植株上有花絮,说明整个种群中有了一定数量的生殖个体,使得种群整体相对比较稳定。
3.2 静态生命表及存活曲线
静态生命表是根据一定时期样地内一个种群所有个体的年龄数据编制而成的[16],通过静态生命表可分析群体的出生率、死亡率等重要参数,提供更多关于种群年龄结构和数量统计方面的信息。以生存分析理论为基础,依据静态生命表的编制要求,编制了绩溪光皮桦种群静态生命表(表2);首先以该群体的龄级作为横坐标,以计算得到的存活量lx和死亡数dx为纵坐标,绘制得到了该光皮桦种群的存活曲线和死亡曲线(图1)。存活曲线的趋势可以大致反映生存率随年龄的增加而发生变化的情况[26]。从图1可以看出,在第Ⅳ龄级前,该光皮桦种群的存活数量下降非常快;而在第Ⅳ径级以后,该种群的存活数量则表现为稳定下降。出现这种情况,可能是光皮桦在幼年阶段,个体不断增大,种间及种内竞争强度随之逐渐增强,这样就会导致幼龄个体大量死亡;当光皮桦达到成熟阶段,种间及种内竞争强度会逐渐降低,光皮桦种群的存活数量则保持在稳定水平。
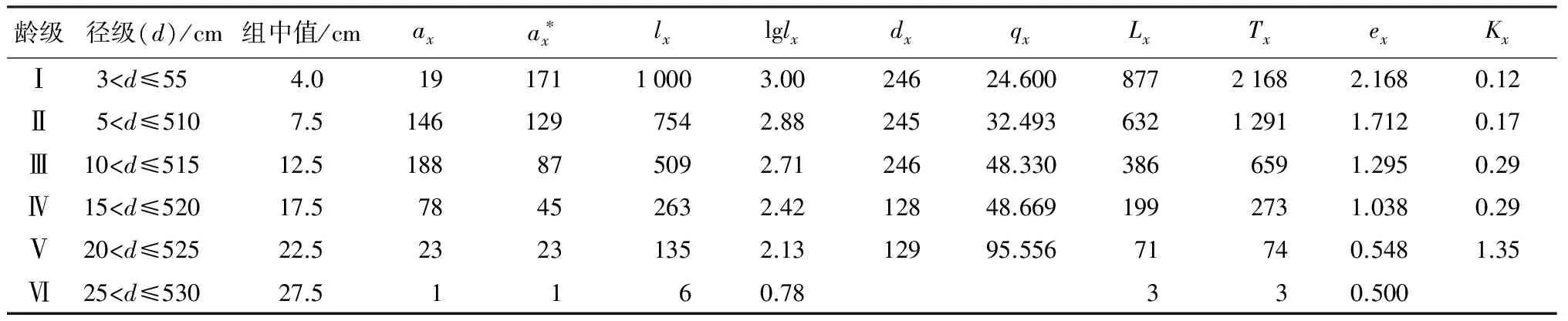
表2 光皮桦种群静态生命表
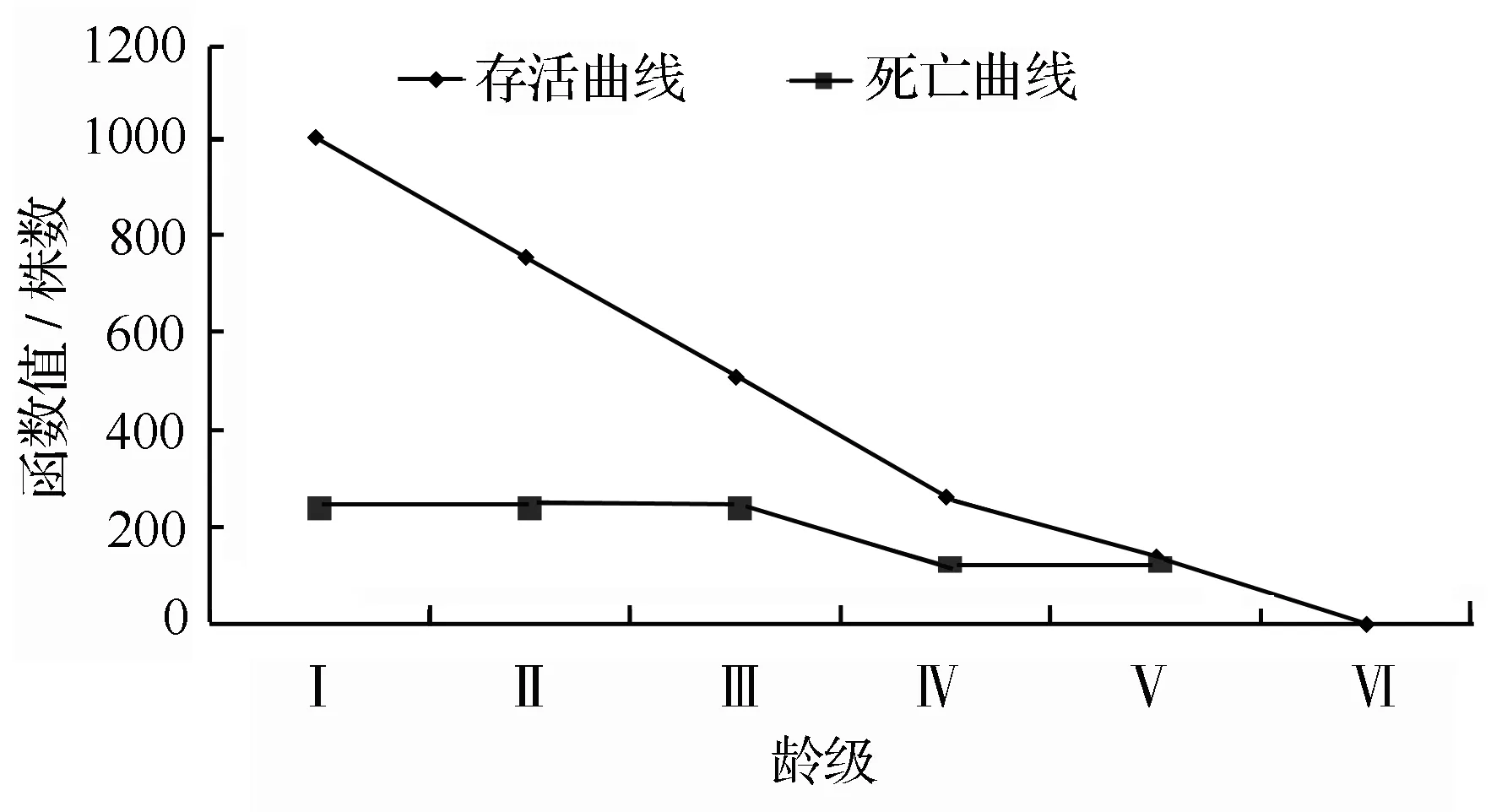
图1 种群的存活曲线(lx)和死亡曲线(dx)
生命期望值表现的是个体的平均生存能力大小[27]。由表2可知,光皮桦种群生命期望值在第I级达到最大,说明这个阶段光皮桦林的生存状况是最好的,处在生理活动的旺盛阶段。从第Ⅱ~Ⅵ级,期望值(ex)都是下降趋势。前期下降可能是植株竞争较强,受到环境筛选和外界干扰的影响,后期下降可能是由于光皮桦种群个体的生理衰退及生存质量下降所致。
Deevey[28]划分了3种类型的存活曲线:I型(呈凸型),采用指数方程式Nx=N0e-bx来描述,表示种群中绝大数植株都能达到平均生理寿命,而且在达到平均寿命后,几乎同时死亡;II型(呈对角线型),用直线方程式Nx=N0+bx描述,表示各径级植株个体死亡率大约相等;III型(呈凹型),以幂函数式Nx=N0x-b描述,含义是植株的幼体死亡率高,随着植株的生长,其死亡率有所降低,且比较稳定。选用以上三种数学模型对光皮桦种群的存活曲线类型进行检验,建立其相应模型,即Nx=10.664e-0.222x(F=7.914,r=0.664);Nx=8.479-0.897x(F=15.421,r=0.794);Nx=8.757x-0.528(F=3.585,r=0.473),检验结果表明绩溪光皮桦种群存活曲线为Deevey-Ⅱ型。
3.3 死亡率和亏损率曲线
以光皮桦种群的龄级为横座标,分别以死亡率(qx)和亏损率(Kx)为纵座标,绘制得到了该光皮桦种群死亡率和亏损率曲线,由图2可以看出,光皮桦种群的死亡率和亏损率曲线的变化趋势基本是一致;在第I、Ⅱ、III、Ⅳ龄级,个体会出现大量死亡;当光皮桦种群从Ⅳ龄级向Ⅴ龄级转化时出现了一个死亡率高峰。
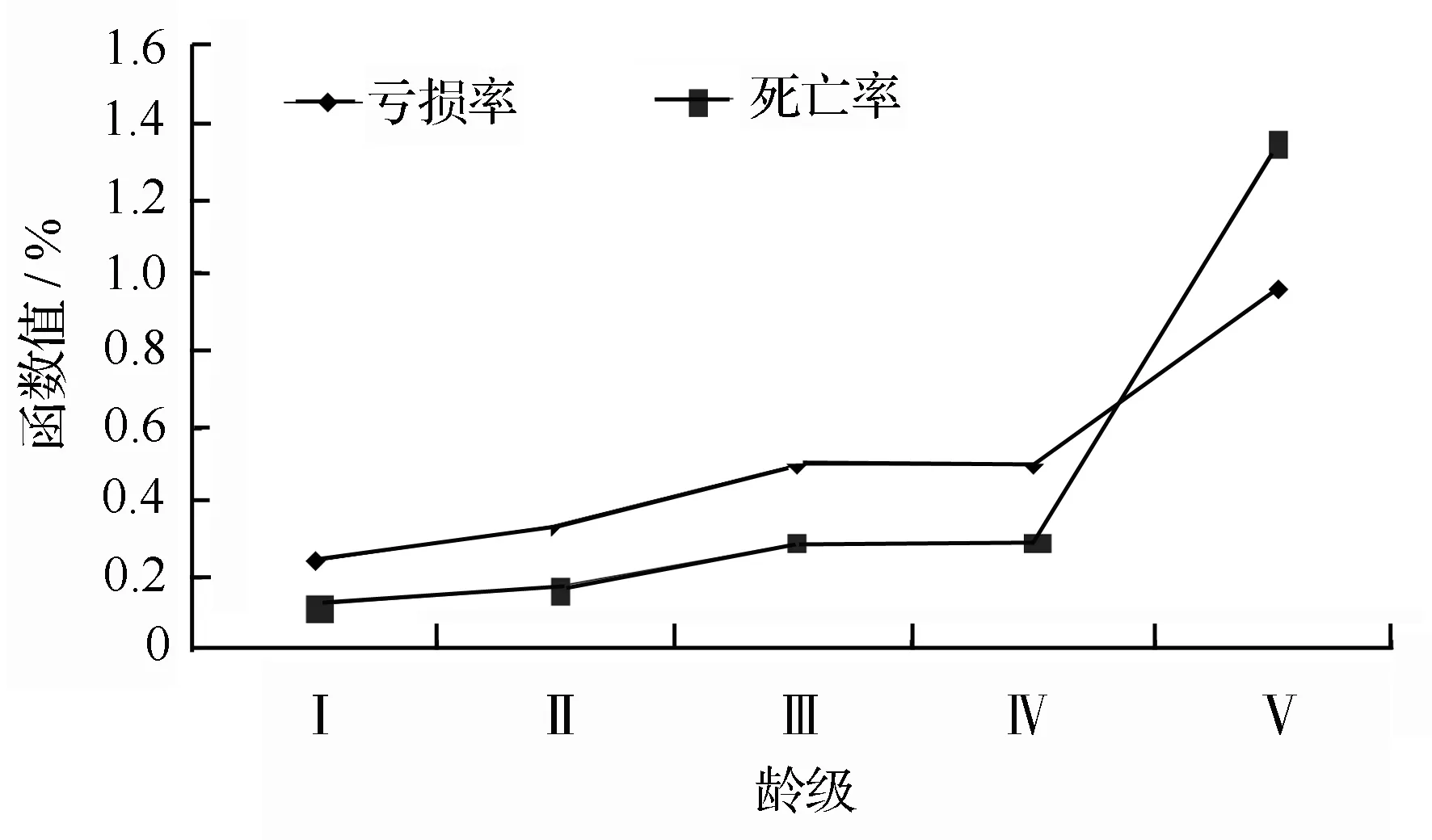
图2 绩溪光皮桦种群亏损率(qx)和死亡率(Kx)曲线
3.4 种群生存函数分析
绩溪光皮桦种群生存分析函数(见表3)。通过绩溪光皮桦种群生存分析函数绘制得到光皮桦的生存函数曲线(图3)可以看出,在第Ⅴ龄级前该种群的生存率曲线和积累死亡率曲线变化幅度都较大;但从第Ⅴ龄级开始,种群生存率曲线和积累死亡率曲线的趋势则变缓;第Ⅴ龄级以后,种群进入生理衰老期,呈现衰退症状。分析结果表明,该光皮桦种群的生存率和累积死亡率变化情况为前期变化幅度较大而后期变化幅度较小,说明在生长前期该种群数量很不稳定,到达后期其数量才趋向稳定。
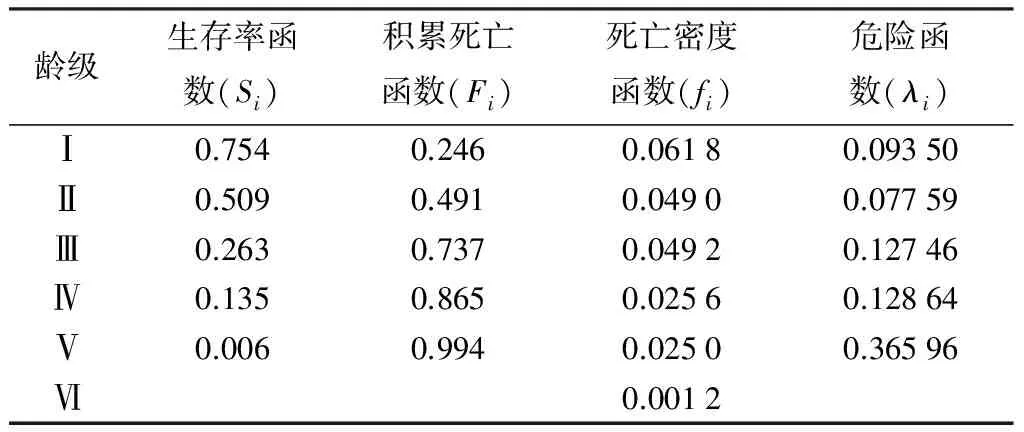
表3 绩溪光皮桦种群生存分析函数
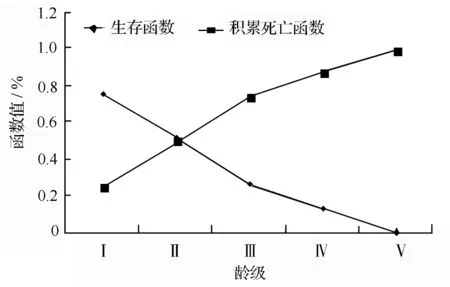
图3 光皮桦种群生存函数和累积死亡率函数曲线
通过死亡密度曲线和危险率曲线(图4)可以看出,光皮桦种群的死亡密度曲线和危险曲线变化表现出明显的反差,死亡密度曲线逐渐减低。危险曲线只是在第I龄级向第Ⅱ龄级转化时从6.18%减低到4.90%,从第Ⅱ龄级以后都是递增趋势。而且第Ⅳ龄级向第Ⅴ龄级转化时增幅比较大,从12.864%增加到36.596%。
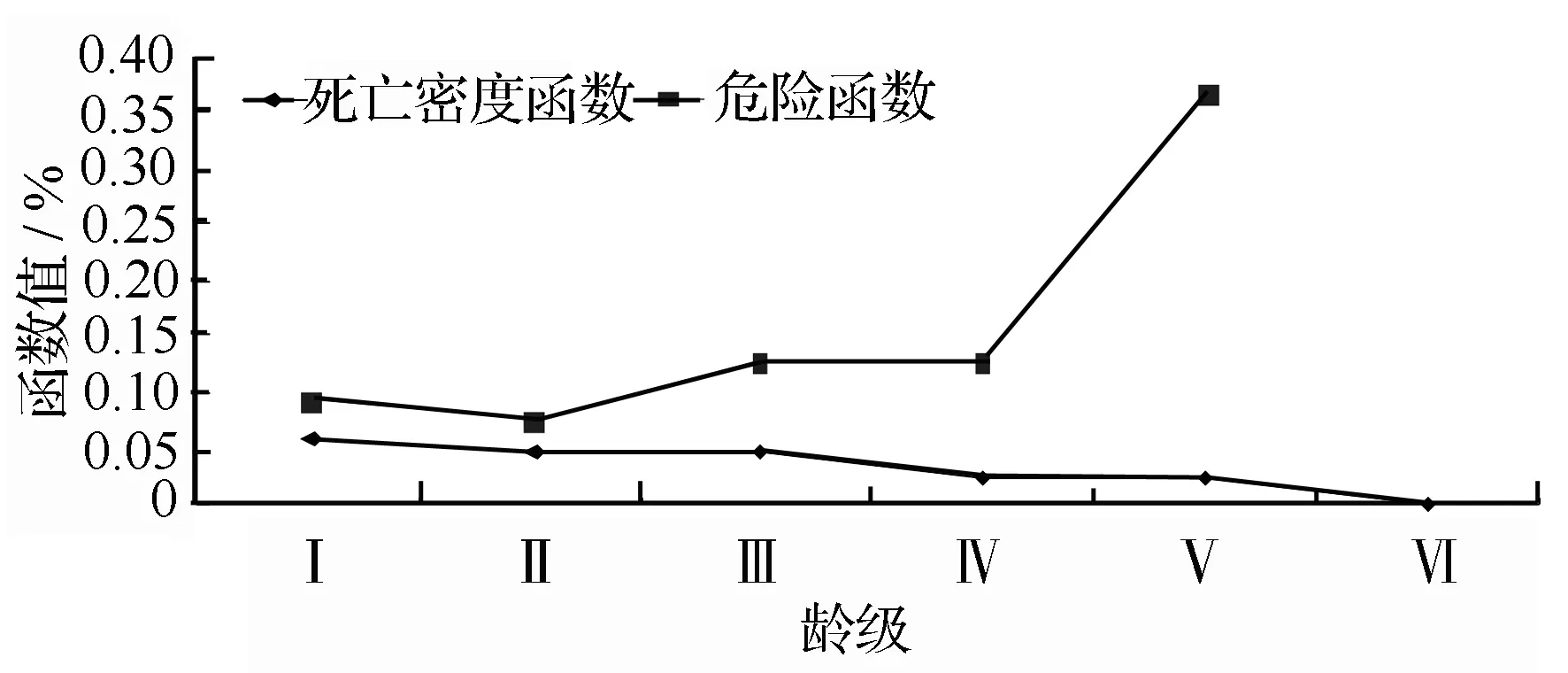
图4 光皮桦种群死亡密度和危险率曲线
4 讨论与结论
本研究调查地点位于山体中上部,以光皮桦为优势的落叶阔叶林,属地带性植被和珍贵的乡土树种。在调查中没有发现幼苗幼树的存在,种群整体上以中龄林数量相对较多,种群龄级结构属纺锤形,数量动态指数为正值,为稳定性种群,但数值偏低,结构增长性很低,有可能进一步衰退。种群生命表反映了绩溪光皮桦种群早期死亡率低,后期死亡率高的动态变化特征,存活曲线趋于Deevey-II型。后期死亡率高的原因是种内竞争,调查中发现部分树高10 m左右的光皮桦,相距1 m左右,调查区域内光皮桦是优势树种,部分区域光皮桦占到80%以上,种内竞争很激烈。每个个体侵占彼此对方的植株生长空间,个体与个体之间相互排挤,对营养空间的需求也逐渐增大,群体内的光照、水分、养分和空间等因子不能满足植株生长的要求,植株间竞争加大,自疏作用增强,个体间的分化现象增加,导致群体死亡率增高,而且山地土层过薄造成根系相互缠结,争夺养分,调查中发现两棵树的根部已经融合一体了。孟广涛等[11]对云南富源光皮桦种群研究表明种群存活曲线属Deevey-III型,种群处于中、幼龄林阶段。光皮桦种群更新能力好于绩溪种群,主要原因是绩溪光皮桦所在林分郁闭度达到0.8,高于富源县光皮桦种源的林分,光皮桦是中性偏阳树种,没有充分的阳光,幼苗很难生长。在郁闭度较大的林分中,仅有成熟的光皮桦与其他阔叶树混生,林下无光皮桦更新层和演替层[29]。其次,受皖南山区气候影响,有的年份很少有光皮桦种子[30-31]。光皮桦种子发育成幼体后,在适宜的环境情况下可以稳定生长,随着植株个体的生长,进入中期后,将受到种内、种间树种对光照、水分、养分和空间生态位的竞争,环境筛选的强度增大,导致光皮桦死亡量增高。在生长到一定树龄时,种群开始衰退,光皮桦种群在第Ⅳ和第Ⅴ龄级的年龄阶段出现死亡高峰。研究的4个生存函数也充分体现了光皮桦的生存规律,累积死亡率单调增加,生存率单调下降,其增加或下降幅度是后期高于前期。
[1] 郑万钧.中国树木志:第2卷[M].北京:中国林业出版社,1985.
[2] 陈存及,陈伙法.阔叶树种栽培[M].北京:中国林业出版社,2000.
[3] 吴子诚,王乐辉.光皮桦选择群体特征和改良技术[J].四川林业科技,1996,17(4):17-28.
[4] 潘新建.光皮桦资源的开发利用与发展前景[J].资源开发与市场,2000,16(4):220-221.
[5] 吴大荣.罗卜岩保护区闽楠等优势植物种群竞争初步研究[J].南京林业大学学报,1998,22(3):35-38.
[6] 余爱华,宋丁全,刘建峰.用改单纯形法对光皮桦种群Logistic模型及改进模型的拟合[J].江西农业大学学报,2004,26(1):59-62.
[7] 李建明,陈存及,赵大洲,等.光皮桦天然林群落优势种群的种间联结性研究[J].应用生态学报,2001,12(2):168-170.
[8] 李建明,谢芳,张思玉,等.不同干扰强度下光皮桦群落树木物种多样性比较[J].浙江林学院学报,2001,18(4):359-361.
[9] 宋丁全,姜志林,郑作孟,等.卫闽光皮桦种群优势增长动态的初步研究[J].山东农业大学学报:自然科学版,1998,29(4):465-468.
[10] 宋丁全,姜志林,郑作孟,等.光皮桦林优势树种间的竞争[J].南京林业大学学报,2000,24(4):26-28.
[11] 孟广涛,柴勇,方向京,等.云南富源光皮桦种群的生存状况分析[J].东北林业大学学报,2008,36(9):19-21.
[12] 江洪.云杉种群生态[M].北京:中国林业出版社,1992.
[13] 赵学农,刘伦辉.版纳青梅种群结构动态与分布格局[J].植物学报,1993,35(7):552-560.
[14] Brodie C, Howle G, Fortin M J. Development of aPopulusbalasamiferaclone in subaretic Quebec reconstructed from spatial analyses[J]. Journal of Ecology,1995,83(2):309-320.
[15] 苏建荣,张志钧,邓疆,等.云南红豆杉种群结构与生命表分析[J].林业科学研究,2005,18(6):651-656.
[16] 吴承祯,洪伟,谢金寿,等.珍稀濒危植物长苞铁杉种群生命表分析[J].应用生态学报,2000,11(3):333-336.
[17] 殷东生,张海峰,王福德.小兴安岭白桦种群径级结构与生命表分析[J].林业科技开发,2009,23(6):40-43.
[18] 吴承桢,吴继林.珍稀濒危植物秃杉种群生命表分析[J].应用生态学报,2000,11(3):333-336.
[19] 陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217.
[20] 陈远征,马祥庆,冯丽贞,等.濒危植物沉水樟的种群生命表和谱分析[J].生态学报,2006,26(12):4267-4272.
[21] 刘任涛,毕润成,闫桂琴,等.山西南部翅果油树种群动态与谱分析[J].植物研究,2007,27(5):550-555.
[22] 周建云,李荣,张文辉,等.不同间伐强度下辽东栎种群结构特征与空间分布格局[J].林业科学,2012,48(4):149-155.
[23] 冯士雍.生存分析Ⅲ[J].数学的认识与实践,1983(1):70-76.
[24] 韩路,王海珍,彭杰,等.塔里木河上游灰胡杨种群生存分析[J].林业科学,2010,46(1):131-135.
[25] 丁国民,刘兴明,倪自银,等.甘肃祁连山青海云杉种群数量动态的初步研究[J].植物资源与环境学报,2005,14(4):36-41.
[26] 关锐敏,王孝安,郭华.黄土高原马栏林区顶级群落的种间关联分析[J].干旱区研究,2009,26(4):569-573.
[27] 祝宁.植物种群生态学导论[M].哈尔滨:东北林业大学出版社,1982.
[28] Deevey E S. Life table for natural population of animals[J]. Q Ver Biol,1947,22(4):283-314.
[29] 胡灶新.光皮桦采种育苗技术[J].安徽林业,2004(3):13-14.
[30] 宋廷茂.林木种子[M].北京:中国林业出版社,1984.
[31] 李建民.光皮桦天然林群落特征研究[J].林业科学,2004,36(2):122-124.
Population Structure Dynamics ofBetulaluminiferaSecondary Forest in Jixi County of Anhui Province, China/
Wan Zhibing(Huangshan University, Huangshan 245041, P. R. China); Cheng Guoyuan(Zhentou Forest Farm of Jixi County); Ding Qiangqiang, Fang Lejin(Huangshan University)//Journal of Northeast Forestry University.-2014,42(1).-27~30,60
Betulaluminifera; Age class; Static life table; Population structure; Population dynamics
1) 省级优秀青年人才基金项目(2012SQRL185)。
万志兵,男,1980年12月生,黄山学院生命与环境科学学院,博士研究生,副教授。E-mail:wanzb626@hsu.edu.cn。
方乐金,黄山学院生命与环境科学学院,教授。E-mail:flj@hsu.edu.cn。
2013年5月5日。
S792.15
责任编辑:王广建。
The experiment was conducted to determine population structure dynamic and quantitative statistics ofBetulaluminiferain a secondary forest of Jixi county, Anhui Province. By the population life table and survival analysis, the life table ofBetulaluminiferapopulation was constructed and age classes were replaced with size-classes, therefore, the population structure dynamic ofB.luminiferawas explored. The results of the population structure illustrates a spindle type with a gradually decline in population. The individual plants of class I and class II are not enough. The survival curve in the population is a Deevey-Ⅱ type with a peak of mortality in size class V. The population size ofB.luminiferadeclines sharply in early stage, stabilizes in middle stage and falls rapidly in last stage.
