昼夜开花萱草杂交后代花朵开闭时间1)
2014-07-27贾贺燕高亦珂张启翔
贾贺燕 高亦珂 何 琦 朱 琳 张启翔
(北京林业大学,北京,100083) (国家花卉工程技术研究中心(北京林业大学))
昼夜开花萱草杂交后代花朵开闭时间1)
贾贺燕 高亦珂 何 琦 朱 琳 张启翔
(北京林业大学,北京,100083) (国家花卉工程技术研究中心(北京林业大学))
萱草属(Hemerocallis)植物依据花朵的开放时间分为夜间开花型和白天开花型两类。为研究萱草花朵开放和闭合的规律,对3种夜间开花的萱草种和品种以及4种白天开花萱草品种进行了杂交,设计了4对杂交组合,每个组合包括正反交,共获得F1代386株。此外,研究了16个辅助杂交组合,后代共208株。2012年和2013年夏季对全部植株的花朵开放和闭合时间进行观测。对于花朵开放时间,北黄花菜(Hemerocallislilio-asphodelus)和黄花菜(H.citrina)做亲本的杂交组合后代中99.74%的植株表现为夜间开放;黄花菜品种‘四月花’(H. ‘April Flower’)做亲本的各杂交组合中夜间开花的‘诺米路’ב四月花’杂交组合的后代全部表现为夜间开放;与白天开花型杂交的3个组合后代中夜间开放类型与白天开放类型株数的比例接近1∶1或3∶1。花朵闭合时间变异较大,黄花菜和北黄花菜做亲本的杂交后代主要集中在夜间和白天闭合;‘四月花’杂交后代在一天中各个时段均有闭合。 结果表明:萱草花朵开放与闭合是受不同基因控制的;花朵开放受一个主基因控制,夜间开花性状是显性,白天开花则为隐性性状;控制花朵开放时间的基因属于核基因;萱草花朵闭合时间表现复杂,可能受多基因调控。
萱草;花朵开放时间;花朵闭合时间;基因
Journal of Northeast Forestry University.-2014,42(7).-100~104
There are two types ofHemerocallis, day blooming species and night blooming species. We studied the genetic development of the flower opening and closing time inHemerocallis. We designed the crossing experiments including 4 combinations and 16 auxiliary combinations between nocturnal blooming daylily species and diurnal blooming cultivars. We observed the flowering behavior of all 594 F1hybrids in 2012 and 2013. For flower opening time, 99.74% of the F1hybrids ofH.citrinaandH.lilio-asphodeluswere highly consistent and all of them showed nocturnal blooming. All the hybrids ofH. ‘Naomi Ruth’×H. ‘April Flower’ showed nocturnal blooming, and the ratio of evening flowering and morning flowering among F1hybrids of other combinations ofH. ‘April flower’ was close to 1∶1 or 3∶1. The closing time of F1hybrids ofH.citrinaandH.lilio-asphodeluswas at day or night, while closing time of F1hybrids ofH. ‘April Flower’ was at the whole day. Therefore, the time of flower opening and the start of closing are regulated by different genes and a dominant allele on a single locus largely determines this phenotype. Nocturnal flowering is a dominant trait and the flower opening and closing times are not controlled by plasmagene but by karyogene. The closing times are more complex and probably controled by multi-genes.
KeywordsHemerocallis; Blooming time; Closing time; Gene
在开花植物中,花朵授粉的成功与否依赖于花朵的开放和闭合时间[1]。花朵的开放和闭合时间也决定了观花植物单朵花期的长度,对花卉的观赏价值有重要影响。林奈(Linnaeus)发现植物开花都有确定的时间,由植物的内源生物节律调控,植物的花朵在一天中确定的时间开放,据此于1751年绘制了“花钟”,用植物的这一特点来提醒时间[2]。Bünning研究发现,植物确定的花朵开闭合时间是受基因控制的。可是由于植物材料的限制,对于花朵开放闭合时间相关基因的研究极少[3]。
萱草属植物有10余种,国产3个夜间开花类型种为黄花菜、北黄花菜、小黄花菜,其余国产种都为白天开花类型[4-6]。不同种萱草的单朵花开放时长约10 h。萱草的品种也分为白天开花和夜间开花两种类型,其中,黄花菜品种多为夜间开花型。萱草属中白天、晚上开花类型间杂交可育[7],因此,萱草属植物是研究昼间开花类型和夜间开花类型的开花习性以及植物花朵开放和闭合基因调控机制的好材料[8],将为萱草的单花长花期育种提供理论依据,是一个植物学和园艺学研究的重点与热点。
Hasegawa et al.[9]在对萱草(H.fulva)和黄花菜天然杂交后代和人工杂交F1代的花朵开放和闭合时间的研究中认为是一个显性主基因控制着花朵开放时间,夜间开花对于白天开花是隐性性状。夜间开花与白天开花的显隐型关系与2008─2009年课题组对萱草白天开花和夜间开花杂交后代的开花时间观察结果相反。为进一步研究萱草白天开花和夜间开花性状的遗传规律,2010年设计了更多的萱草白天开花和夜间开花类型之间的杂交组合,对其F1代植株的花朵开放时间和闭合时间进行了观测,并基于观测结果进一步分析了萱草花朵开放闭合时间的遗传规律和基因调控。
1 材料与方法
夜间开花类型:北黄花菜(H.lilio-asphodelus)、黄花菜(H.citrina)和黄花菜品种‘四月花’(H. ‘April flower’)。品种‘贝拉’(H. ‘Bela Lugosi’)、‘诺米路’(H. ‘Naomi Ruth’)、‘红酒’(H. ‘Red Rum’)、‘蓝光’(H. ‘Blue Sheen’)经多次观察也为夜间开始开放,也归为夜间开花类型。
白天开花类型:‘儿童节’(H. ‘Children’s Festival’)、‘红色海盗’(H. ‘Crimson Pirate’)、‘紫泉’(H. ‘Purple water’)、‘凯乌德’(H. ‘Catherine Woodbury’)、‘盛夏酒红’(H. ‘Summer Wine’)、萱草(H.fulva)。
所有的植物材料都种植于位于北京市昌平区小汤山镇的国家花卉工程技术研究中心基地(北纬40°15′,东经116°44′)。
杂交试验:2009年和2010年夏季分别进行了昼夜开花类型间的杂交,杂交前一天对所有花朵进行严格去雄套袋,授粉后再进行套袋。杂交组合包括:‘四月花’ב儿童节’共42朵,获得11个果实;‘儿童节’ב四月花’共37朵,获得10个果实;北黄花菜ב儿童节’共65朵,获得24个果实;‘儿童节’×北黄花菜共35朵,获得12个果实;‘四月花’ב红色海盗’共32朵,获得9个果实;‘红色海盗’ב四月花’共30朵,获得8个果实;黄花菜ב红色海盗’共53朵,获得18个果实;‘红色海盗’×黄花菜共46朵,获得16个果实,共计得到386棵杂种苗。16个辅助杂交组合,包括北黄花菜做母本,‘凯乌德’、‘紫泉’、‘贝拉’、‘诺米路’、‘红酒’、‘蓝光’分别做父本的6个杂交组合和北黄花菜自交组合,黄花菜做母本,‘盛夏酒红’、‘紫泉’、‘贝拉’、‘诺米路’、‘儿童节’、‘凯乌德’分别做父本的6个杂交组合;黄花菜做父本的组合包括萱草×黄花菜,‘四月花’做亲本的杂交组合(‘四月花’ב紫泉’、‘诺米路’ב四月花’),共获得208株杂交后代。杂交后约40 d采收种子,播种于温室中,次年春季定植到大田中。2009年杂交所获F1代于2011年夏季首次开花,2010年杂交所获F1代于2012年夏季首次开花。2013年夏季(7─8月份)对所有F1代以及亲本植株的花朵开放时间和花朵闭合时间进行观察记录。
花朵开放闭合的6个阶段:利用亲本品种‘小酒杯’的一个即将开放的花蕾观察了萱草花朵的开放闭合过程(图1)。整个开放至闭合的过程可以分为6个阶段(或时间点),不同的萱草品种及F1代植株的花朵开放闭合过程均具有这6个阶段,只是各阶段持续时间不完全一致。

1.花朵开始开放时间:萱草花蕾尖端从闭合状态转为开裂状态的时间,从该时间点开始,花朵逐步开放;2.内被片打开时间:此时内被片打开,雌雄蕊可见;3.花朵完全展开阶段:花朵的内外被片均展开至最大角度,为花朵授粉的主要时间段,可持续数小时;4.花朵开始闭合时间:内外被片从完全伸展状态转为开始向内部收缩的时间,从该时间点起,花朵逐步闭合、收缩;5.内被片闭合时间:此时内被片闭合,部分品种雄蕊不可见,但雌蕊伸长可见;6.花朵完全闭合阶段:内外被片全部闭合,花朵逐渐萎蔫脱落。
图1 萱草花朵开放至闭合的过程
花朵开放闭合时间的测定:对花朵开放闭合过程的6个阶段出现的时间进行记录,并选择花朵开始开放时间和花朵开始闭合时间进行分析,因为,在该时间点花朵发生了质的变化(从该时间点开始花朵逐步达到开放状态或闭合状态),因此能更好地揭示花朵内部由基因控制的生理变化。在花期对所有杂交组合后代共594株以及亲本的花朵开放闭合时间进行观测,每15 min观测1次,每株于不同日期共观察3朵花作为重复,取其均值。由于花朵开放闭合时间受阴雨天气影响,故分析时删除了阴雨天的记录数据。用软件R2.15.1对F1代植株花朵的开放闭合时间绘制了时钟图(图2),图中某时间点对应的黑柱表示在该时间开放或闭合的花朵数量。其他数据分析用软件SPSS18.0(PASW18.0)完成。
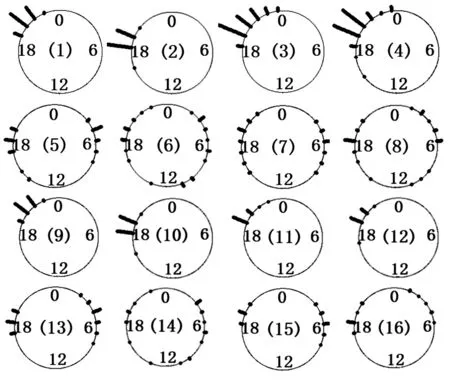
(1).黄花菜ב红色海盗’F1代花朵开放时间;(2).黄花菜ב红色海盗’F1代花朵闭合时间;(3).北黄花菜ב儿童节’F1代花朵开放时间;(4).北黄花菜ב儿童节’F1代花朵闭合时间;(5).‘四月花’ב儿童节’F1代花朵开放时间;(6).‘四月花’ב儿童节’F1代花朵闭合时间;(7).‘四月花’ב红色海盗’F1代花朵开放时间;(8).‘四月花’ב红色海盗’F1代花朵闭合时间;(9).‘红色海盗’×黄花菜F1代花朵开放时间;(10).‘红色海盗’×黄花菜F1代花朵闭合时间;(11).‘儿童节’×北黄花菜F1代花朵开放时间;(12).‘儿童节’×北黄花菜F1代花朵闭合时间;(13).‘儿童节’ב四月花’F1代花朵开放时间;(14).‘儿童节’ב四月花’F1代花朵闭合时间;(15).‘红色海盗’ב四月花’F1代花朵开放时间;(16).‘红色海盗’ב四月花’F1代花朵闭合时间。
图2 萱草白天、夜间开花类型杂交F1代花朵开放闭合时钟图
2 结果与分析
2.1 亲本花朵开放和闭合时间
萱草的花朵开放闭合时间虽然可以分为两大类,但是具体的开闭时间不完全相同(表1),对于夜间开花亲本黄花菜、北黄花菜和四月花,它们的花朵均在傍晚开始开放,2~3 h后达到盛开状态,之后这种盛开状态一直持续到次日凌晨,之后逐渐闭合,整个过程持续12 h左右。由于这3个亲本的花朵达到盛开状态的时间是在夜间,并且花朵的盛开状态也主要持续在夜间,因此将它们归为夜间开花类型。白天开花亲本儿童节和红色海盗则是在凌晨开始开放,3~4 h后达到盛开状态,盛开状态发生在白天,故归为白天开放类型。对于F1代,按照同样的标准,也可将其分为两类:傍晚开始开放,夜间达到盛开状态的归为夜间开花类型;凌晨开始开放,白天达到盛开状态的归为白天开花类型。
表1 4个萱草杂交组合中的亲本花朵开放闭合时间及单朵花持续开放时间
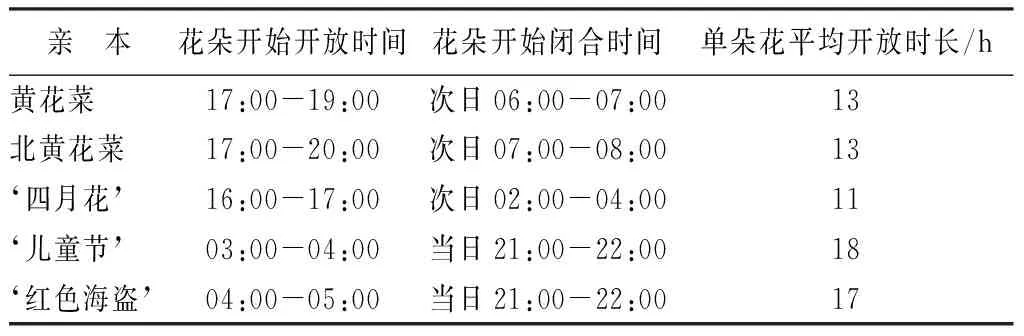
亲 本花朵开始开放时间花朵开始闭合时间单朵花平均开放时长/h黄花菜17:00-19:00次日06:00-07:0013北黄花菜17:00-20:00次日07:00-08:0013‘四月花’16:00-17:00次日02:00-04:0011‘儿童节’03:00-04:00当日21:00-22:0018‘红色海盗’04:00-05:00当日21:00-22:0017
2.2 白天开花和夜间开花萱草杂交F1代花朵开放和闭合时间
杂交组合黄花菜ב红色海盗’和北黄花菜ב儿童节’杂交F1代共227株,其花朵开放时间和闭合时间表现出高度的一致性,均为夜间开花(夜间开花类型植株的比例均为100%),并持续开放约24 h,次日晚上闭合;其中,黄花菜ב红色海盗’F1代花朵开放时间分布在19:10─22:45,在20:15有明显峰值,北黄花菜ב儿童节’F1代花朵开放时间分布在18:15─23:45,在19:00有明显峰值(图2)。
杂交组合‘四月花’ב儿童节’F1代共48株,其中26株夜间开放,22株白天开放,比例未偏离1∶1(χ2=0.333,P=0.564);反交组合‘儿童节’ב四月花’F1代共36株,其中16株夜间开放,20株白天开放,比例未偏离1∶1(χ2=0.444,P=0.505);‘四月花’ב红色海盗’F1代共42株,其中24株夜间开放,18株白天开放,分离比例未偏离1∶1(χ2=0.857,P=0.355);反交组合‘红色海盗’ב四月花’F1代共35株,其中19株夜间开放,16株白天开放,结果同样未偏离1∶1(χ2=0.257,P=0.612)。‘四月花’后代花朵开放的峰值时间与亲本一致,但主峰不太明显,有部分植株花朵开放时间与亲本‘四月花’不一致。
对于各杂交组合后代的花朵闭合时间,当黄花菜或北黄花菜为亲本时,F1代闭合时间高度一致,均为次日晚上闭合,峰值时间与其白天开花亲本接近,单朵花开放时长达24 h左右,超过其亲本植株。而当‘四月花’为亲本之一时,F1代花朵闭合时间变异较大,几乎在一天的各个时段都有分布,缺乏明显的峰值。
以上4个组合的反交组合花朵开放闭合时间与4个正交组合具有相似的分布和峰值,差异不显著,这表明萱草科植物的花朵开放闭合时间主要由核基因控制,受细胞质基因影响较小。
2.3 辅助杂交组合F1代群体花朵开放闭合时间
辅助杂交组合中新增加的亲本中白天开放型:‘盛夏酒红’,03:00─04:30开放,20:00─21:30闭合;‘凯乌德’,03:00─05:00开放,20:00─21:00闭合;萱草,03:30─05:30开放,19:00─20:30闭合;‘紫泉’,04:00─05:00开放,20:00─21:00闭合。夜间开放型:‘贝拉’,21:00─23:00开放,次日21:30─23:00闭合;‘蓝光’,21:00─23:00开放,次日21:30─22:30闭合;‘红酒’,21:30─23:00开放,次日21:00─23:00闭合;‘诺米路’,22:00─23:30开放,次日18:00─20:00闭合。
夜间开放型×夜间开放型8个杂交组合中的7个组合的杂交后代全部表现为夜间开花(表2),一个例外组合是黄花菜ב贝拉’8株后代中7株夜间开花,1株表现为白天开花,但亲本‘贝拉’是4倍体,推测杂交后代的倍性会影响杂交后代的基因显隐关系;北黄花菜自交共获得18株后代,全部后代都表现为夜间开花,花朵开放的峰值时间为19:00,与亲本北黄花菜一致;‘诺米路’ב四月花’的后代也全部表现为夜间开花。

表2 辅助杂交组合F1代群体花朵开放闭合各类型株数
白天开放型×夜间开放型杂交组合12个及北黄花菜做亲本的3个组合的杂交后代全部表现为夜间开花;黄花菜做亲本的6个组合后代也都表现为夜间开花类型;亲本‘四月花’的后代表现则不同,‘四月花’ב紫泉’后代27株中有22株表现为夜间开花,5株表现为白天开花,二者的比例未偏离3∶1(χ2=0.605,P=0.437)。
对于花朵闭合时间,组合北黄花菜ב凯乌德’、北黄花菜ב贝拉’、北黄花菜ב蓝光’、萱草×黄花菜的后代表现比较一致,均为夜间闭合,其余12个组合后代的花朵闭合时间变异较大,在一天各时间段均有闭合,无明显规律,说明调控花朵闭合的基因比较复杂。
3 结论与讨论
萱草花朵开放与闭合受不同的基因调控。在所研究的20个杂交组合中,萱草花朵开放时间与闭合时间在各组合杂交后代中表现出完全不同的分离规律,表明萱草的花朵开放时间和闭合时间受不同基因控制。
萱草花朵开放时间受一个主基因控制。基于重点杂交组合和辅助杂交组合后代的花朵开放时间的观察结果,萱草的花朵开放时间具有一对相对性状,即白天开花与夜间开花,判断夜间开花(N)对于白天开花(n)是显性性状,北黄花菜、黄花菜是纯合体(NN),所以,北黄花菜和黄花菜不论与夜间开花还是白天开花的二倍体萱草杂交,后代都是夜间开花。而黄花菜的品种‘四月花’是杂合体(Nn),因此,‘四月花’后代与北黄花菜和黄花菜有不同表现,与夜间开花亲本杂交,后代全部夜间开花;而与白天开花的萱草杂交,后代出现了夜间开放类型与白天开放类型株数接近1∶1或3∶1的分离比,该分离比符合孟德尔单基因分离定律。这与Hasegawa et al.[9]认为花朵白天开放对于夜间开放是显性的研究结论相反,这也正是当初发现结论与本课题组研究杂交结果不同而进行大量杂交的原因。Hasegawa et al.[9]用于推断显隐性关系的群体只有一个,是将萱草和黄花菜进行人工杂交得到的47株F1代个体(黄花菜做母本得到32株,萱草做母本得到15株),其中大多数(44株)表现为白天开花。该试验组合单一,只有2个亲本,而且杂交亲本之一的萱草多数为3倍体,课题组进行杂交验证的多倍体组合黄花菜ב贝拉’也表现的与2倍体组合不同,表明杂交后代的倍性会影响杂交后代的基因显隐关系的判别。课题组验证中萱草×黄花菜组合后代虽然少,但是花朵开放时间与Hasegawa et al.[9]的结果相反,是夜间开花的。
萱草花朵的开放时间主要受核基因控制。萱草花朵的开放时间在4个正反交组合后代中表现一致,表明花朵开放时间性状受细胞质基因影响极小,主要受核基因控制。F1代植株中仍然存在个别植株的开花时间与亲本表现的不一致,这表明可能存在其他次要的基因与主基因互作,共同调控花朵的开放时间,这与Matsumoto et al.[8]的研究结果一致。
萱草花朵闭合时间比较复杂。杂交组合北黄花菜和黄花菜做亲本的5个组合后代花朵闭合时间较集中,均为夜间闭合,而其余10个杂交组合后代以及北黄花菜自交的后代花朵白天和夜间均有闭合,且后代分离比例偏离3∶1或1∶1;黄花菜品种‘四月花’的杂交后代花朵闭合时间变异极大,几乎在一天的各个时段都有分布,缺乏明显的峰值。这一结果证明,萱草花朵的闭合时间受多基因调控。这与Nitta et al.[10]的研究结果也是不同的,他根据萱草和黄花菜杂交后代推断萱草花朵闭合受一个主基因调控。而课题组对20个昼夜开花类型萱草间杂交后代的研究结果证明,萱草花朵闭合时间较为复杂。
萱草的单花花期在白天开花与夜间开花的杂交后代中得到了延长。黄花菜ב红色海盗’和北黄花菜ב儿童节’杂交后代的单花花期从亲本的12 h左右,延长到24 h左右,表明利用萱草不同花朵开放时间的亲本杂交,可以延长萱草单朵花期,增加萱草的观赏时间。在试验设计的20个杂交组合中,亲本以及F1代植株,不论白天开放还是夜间开放,单朵花持续开放的时间均为12 h或12 h的倍数,如24 h。其中黄花菜和北黄花菜后代全为24 h,不同组合的‘四月花’后代分别有55.56%和52.63%为24 h,其余为12 h,表明调控开花的生化途径的启动与关闭是以12 h为时间间隔进行的,相关基因是在植物自身的生物节律下起调控作用,该结论与Yanovsky et al.[11]的研究结果一致。
白天开花萱草的虫媒主要为蝶类、蜂类,而傍晚开花萱草的虫媒主要为蛾类,不同种类萱草的主要传粉生物及其相关生物学特性可能是萱草开闭花机制形成的原因之一,另外,原产北方的大苞萱草、北萱草、小萱草、北黄花菜、小黄花菜等花期(5─6月份)较之偏南分布的种类花期(6─8月份)显著偏早,因此,花期时段的日照长度可能对单花期也有影响,目前对于花朵的开放与闭合机制尚不清楚[12],也没建立相关的基因控制模式。萱草由于其花朵只开1 d并且天然存在夜间开花和白天开花类群,是花朵开放和闭合研究很好的模式材料,如能够在萱草中进行更全面的杂交,并对夜间开花类型和白天开花类型的F2代进行分析,将能够更好地揭示植物花朵开放闭合时间的遗传规律和基因调控机理,为揭示植物花朵开放和闭合的遗传规律奠定基础,也为更进一步地研究延长花卉的观赏期提供理论基础。
[1] van Doorn W G. van meeteren U. Flower opening and closure: a review[J]. Journal of Experimental Botany,2003,389(54):1801-1812.
[2] Moore-Ede M C, Sulzman F M, Fuller C A. The clocks that time us: physiology of the circadian timing system[M]. Cambridge: Harvard University Press,1982.
[3] 徐小冬,谢启光.植物生物钟研究的历史回顾与最新进展[J].自然杂志,2013,35(2):118-126.
[4] 熊治廷,陈心启,洪德元.国产萱草属夜间开花类群的分类研究[J].植物分类学报,1996,34(6):586-591.
[5] 陈心启.中国植物志:第14卷[M].北京:科学出版社,1999:52-54.
[6] Stout A B. Types of anthesis inHemerocallisand their heredity in F1hybrids[J]. Bull Torrey Bot Club,1946,73(2):134-154.
[7] 何琦.不同倍性萱草(Hemerocallisspp. &cvs.)杂交育种研究[D].北京:北京林业大学,2012.
[8] Matsumoto T, Yasumoto A A, Nitta K. Difference in flowering time as an isolating barrier[J]. Journal of Theoretical Biology,2013,317:161-167.
[9] Hasegawa M, Yahara T, Yasumoto A, et al. Bimodal distribution of flowering time in a natural hybrid population of daylily (Hemerocallisfulva) and nightlily (Hemerocalliscitrina)[J]. Journal of Plant Research,2006,119(1):63-68.
[10] Nitta K, Yasumoto A A, Yahara T. Variation of flower opening and closing times in F1and F2hybrids of daylily (Hemerocallisfulva; Hemerocallidaceae) and nightlily (H.citrina)[J]. American Journal of Botany,2010,97(2):261-267.
[11] Yanovsky M J, Kay S A. Living by the calendar: How plants know when to flower[J]. Nature Reviews Molecular Cell Biology,2003,4(4):265-275.
[12] van Schie C N, Haring M A, schuurink C R. Regulation of terpenoid and benzenoid production in flowers[J]. Current Opinion in Plant Biology,2006,9(2):203-208.
Opening and Closing Time of F1Hybrids of Daylily (Hemerocallisfulva) and Nightlily (Hemerocalliscitrina)/
Jia Heyan, Gao Yike, He Qi, Zhu Lin(Beijing Forestry University, Beijing 100083, P. R. China); Zhang Qixiang(National Engineering Research Center for Floriculture, Beijing Forestry University)//
贾贺燕,男,1988年6月生,北京林业大学园林学院,硕士研究生。
高亦珂,北京林业大学园林学院,教授。E-mail:gaoyk@ bjfu.edu.cn。
2013年9月8日。
S682.1+9; Q32
1) 北京市园林绿化局花卉生产优势品种育种研发与示范项目(YLHH201400114)。
责任编辑:任 俐。
