高盐度海滩红树林造林试验
2014-07-12陈玉军廖宝文邓智泓魏军发
陈玉军,廖宝文,李 玫,邓智泓,魏军发,管 伟
(1 中国林业科学研究院 热带林业研究所,广东 广州510520;2 电白县林业局,广东 电白525400)
红树林是沿海防护林体系的重要组成部分,素有“海上森林”之称,能够防风消浪[1-2],保护海堤,减少和防止沿海地区台风暴潮等自然灾害的发生,保护和发展红树林资源是华南沿海地区生态建设所面临的迫切任务.我国近十年来在华南沿海地区逐步开展了红树林资源恢复工作,在普通的宜林地滩涂上已基本实现或正在进行红树林的恢复和发展工作.由于种种人为或生物学原因,现存的尚未实现造林的潮间带滩涂基本上为高盐度海滩等困难立地.
目前,国内外开展了广泛的红树林资源恢复与发展的研究工作.泰国、菲律宾、墨西哥、美国佛罗里达等国家和地区开展的研究工作涉及人工林与天然林的差异性分析、红树林成功恢复的标准、影响恢复的因素、恢复的必要性与管理对策等方面[3-9].国内目前有多个机构组织开展了红树林恢复和发展研究工作,对造林地选择和树种选择进行了深入研究[10-25].在盐度对红树植物幼苗生长及生理生态影响方面,国内也开展了较多的室内试验研究[26-30].综观国内外红树林恢复研究现状,基本解决了普通宜林地条件下红树林的恢复理论和技术,而对高盐度海滩极端立地条件下红树林野外种植的试验研究,目前鲜有报道.本研究通过在广东电白县水东湾高盐度困难立地开展红树林种植试验,初步确定高盐度条件下红树植物的生长适应性,为我国华南沿海红树林资源的恢复和发展提供理论依据.
1 材料与方法
1.1 试验地概况
本研究地点设立在广东茂名市电白县水东湾的沿岸及二洲岛潮间带区域(21°22'~22°42'N,110°19'~111°41'E).地处北回归线以南,属热带亚热带季风温和气候.年平均气温23 ℃,月平均最高气温28.5 ℃(7月份),月平均最低气温15.68 ℃(1月份).降雨季节长,年降雨量1 500~1 800 mm.由于水东湾海域河流淡水的注入较少,海水盐度高,平均在30‰以上.土壤类型多样(表1),主要为海岸沙质、泥沙质、泥质及硬泥质,其中沙质土壤的肥力较差,硬泥质土壤则影响植物的根系发育.较高的海水盐度和较差的土质条件,均不利于红树植物的生长.所属电白县有红树林天然林165 hm2,人工林600 hm2,红树林种类有白骨壤Avicennia marina、红海榄Rhizophora stylosa、秋茄Kandelia obovata、桐花树Aegiceras corniculatum、木榄Bruguiera gymnorrhiza、拉关木Laguncularia racemosa 和无瓣海桑Sonneratia apetala 等.

表1 不同类型土壤的养分含量Tab.1 Nutrition contents of different soil types
1.2 研究方法
于2010年6月,分别在水东湾渔塘基围和岛屿的潮间带区域试验种植红树植物.试验树种为红海榄、秋茄、拉关木和无瓣海桑,其中红海榄、秋茄为慢生乡土树种,拉关木和无瓣海桑为速生的引进树种.红海榄以实生苗和胚轴苗2 种形式种植(本文中红海榄除特殊注明外,以实生苗种植),其他树种均以实生苗的形式种植.慢生红树植物的试验种植规格分别为0.25 m×0.25 m、0.25 m×0.50 m、0.50 m×0.50 m、0.50 m×1.00 m、1.00 m×1.00 m,速生红树植物的试验种植规格分别为1.00 m×1.00 m、1.50 m×1.50 m、2.00 m×2.00 m.试验种植所选择的土壤质地条件分别为沙质和泥质,立地的滩面高度以平均海平面为参照,海拔高度范围为-0.4~0.7 m.
分别于2011 和2012年6月,即在试验林1年生和2年生时,调查试验地各种红树植物的地径、树高和冠幅生长量,以及各树种的保存率.分沙质地和泥质地2 个试验区域分别调查,根据不同的滩面高度调查取样,以试验地的实测水位再参照当日的潮汐表数据来确定相对平均海平面的高度,数据分析中采用的滩面高度指标为0.2 m 范围内的中间值,如滩面高度0 指滩面高度在-0.1 m~0.1 m 之间,其余以此类推.每个树种采用对不同种植规格和地点随机抽样调查,生长量的测量按每种设计和类型分3个重复,每个重复取样数为20~30 株,保存率的确定按试验设计的全部植株的存活情况统计.
野外观测数据均采用Microsoft Excel 2003 软件处理.采用SPSS 16.0 统计分析软件进行方差分析和差异显著性检验(LSD 法),显著性水平设定为α=0.05.
2 结果与分析
2.1 滩面高度对红树植物生长的影响
2.1.1 沙质土壤条件下滩面高度对红树植物生长的影响 沙质土壤条件下滩面高度对红海榄、秋茄、拉关木和无瓣海桑4 种红树植物2年生时的生长量及保存率影响较大.由表2 可知,滩面高度在0(即平均海平面)附近时,4 种红树植物的生长量和保存率基本达到最大值(仅秋茄的保存率除外),随着滩面高度的增加或降低,生长量和保存率呈下降的趋势,其中,滩面高度在0.7 m 时,生长量和保存率基本降到最小值(仅秋茄的保存率有增加).红海榄在滩面高度为0 时树高和地径最大,分别为1.44 m 和1.1 cm,保存率在90%以上,而在滩面高度为0.7 m 的情况下树高、地径生长量和保存率仅为滩面高度为0 时的40.9%、34.7%和76.5%.秋茄在滩面高度为0.7 m 时的树高和地径生长量仅为滩面高度为0 时的61.1%和65.2%,但保存率增加约1 倍.拉关木在滩面高度为0.7 m 时的树高、地径和冠幅生长量仅为滩面高度为0 时的48.9%、51.2%和46.5%,保存率变化不大.无瓣海桑在滩面高度为0.7 m 时的树高和地径生长量仅为滩面高度为0 时的39.8%和43.5%,保存率更低,仅为滩面高度为0 时的26.8%.

表2 沙质土壤条件下滩面高度对红树植物生长的影响1)Tab.2 Effects of mudflat elevation on the growth of mangrove species on site of sandy soil
2.1.2 泥质土壤条件下滩面高度对红树植物生长的影响 在泥质土壤条件下,滩面高度对红海榄、秋茄、拉关木和无瓣海桑4 种红树植物2年生时的生长量及保存率的影响与在沙质生境中相似,滩面高度在0 附近时生长量和保存率基本上最大.红海榄在滩面高度为0 时树高、地径和保存率分别为1.07 m、1.40 cm 和98.4%,当滩面高度高于或低于此高度时,红海榄的地径生长量和保存率均减低,但当滩面高度高于0 时的地径减低幅度要比低于0 时大得多,在滩面高度为0.7 m 时的树高、地径生长量及保存率仅为滩面高度为0 时的34.6%、40.0%和80.2%.秋茄在滩面高度为0.7 m 时的树高、地径生长量和保存率仅为滩面高度为0 时的75.7%、72.7%和78.4%.从滩面高度为0 上升到滩面高度为0.7 m,拉关木和无瓣海桑的树高、地径和冠幅生长量均降低50%左右,拉关木的保存率由50.8%降至17.4%,无瓣海桑的保存率由39.0%降至15.7%(表3).由此可见,红海榄和秋茄在各种滩面水平上都具有较强的生长适应性,保存率均较高.在较高的滩面水平上,拉关木和无瓣海桑的生长适应性较差,保存率很低.

表3 泥质土壤条件下滩面高度对红树植物生长的影响1)Tab.3 Effects of mudflat elevation on the growth of mangrove species on site of muddy soil
2.2 土壤条件对红树植物生长的影响
试验林种植2年后,在滩面高度不同的情况下,土壤质地对红海榄生长的影响程度具有一定的差异性.在中等滩面高度情况下(滩面高度在0 附近),红海榄在沙质地和泥质地的生长量和保存率很接近.在较高的滩面水平上(滩面高度为0.2 m),红海榄在沙地条件的生长量和保存率要明显高于泥质条件下的情况.但当滩面水平更高时(滩面高度为0.4 m),红海榄地径的生长量在沙质条件下略低于泥质条件,树高生长量在沙质条件下略高于泥质条件(表4).
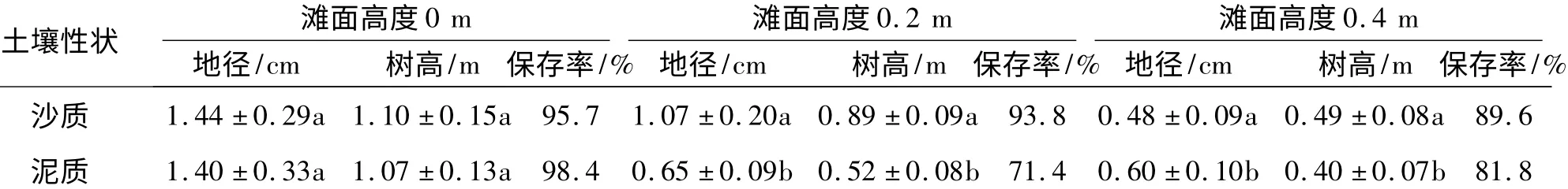
表4 不同土壤条件下红海榄的生长表现1)Tab.4 Growth performance of Rhizophora stylosa in different soil types
2.3 不同树种的生长适应性
2.3.1 沙质地条件下不同树种的生长适应性 在普通沙质地条件下(滩面高度为0),试验林种植2年后,红海榄和秋茄的保存率很低,其中红海榄的保存率低于30%,长势也较差,但以胚轴苗形式种植的红海榄保存率很高(95.7%),极少死亡,长势也很好,拉关木和无瓣海桑的保存率很高,超过80%,长势良好(表5);红海榄和秋茄的生长量较小,地径0.7 cm 左右,树高0.5 m 左右,无瓣海桑的生长量较大,拉关木的生长量最大,地径约6.0 cm,树高约2.0 m.在沙质高滩地极端条件下(滩面高度为0.7 m),试验林种植2年后,所试验的4 个树种中,除了无瓣海桑保存率极低外,红海榄、秋茄和拉关木的保存率均较高,在70%以上,其中拉关木的保存率接近90%;红海榄、秋茄和无瓣海桑的生长量较小,其中红海榄和秋茄地径大于0.4 cm,树高低于0.5 m,拉关木的生长量最大,地径为3.0 cm,树高为1.09 m,但4 个树种的生长量均比一般立地条件下低得多(表5).
可见,在沙质地条件下,与其他树种相比,拉关木的保存率和生长量均最大,表明比其他3 个树种更适合在沙质地上生长.

表5 沙质地条件下各树种的生长表现1)Tab.5 Growth performance of different mangrove species on sandy soil
2.3.2 泥质地条件下不同树种的生长适应性 在泥质地条件下种植2年后,红海榄(2 种种植形式)和秋茄的保存率都很高,在70%以上,长势也较好;拉关木和无瓣海桑的保存率很低,在40%以下,幼树长势也一般.红海榄和秋茄生长量较小,地径仅为0.6 cm 左右,树高仅为0.4 m 左右;拉关木和无瓣海桑的生长量较大,其中拉关木地径超过2.0 cm,树高接近1.0 m(表6).
可见,在泥质地条件下,红海榄和秋茄的生长量较小,但保存率很高,表明其适合在泥质地上种植.拉关木和无瓣海桑的保存率均较低,表明在泥质地上不易成活和保存.

表6 泥质地条件下各树种的生长表现Tab.6 Growth performance of different mangrove species on muddy soil
2.4 海藻对红树林植物生长的影响
由于试验地处于开阔海湾,河流淡水的补充较少,因此海水的盐度较高,另外,试验地海水富营养化严重,在冬春季节会滋生大量海藻,并覆盖红树林幼苗分布区域,退潮后会埋压幼苗、幼树,直接影响和危害幼苗的成活及生长.在迎风区域,海藻多为异地漂移所致,海藻量大且具有流动性,对幼苗、幼树影响较严重,背风区海藻一般为原地生长形成,移动性小,数量相对于迎风面少.由表7 可见:红海榄生长较缓慢,幼树低矮,在迎风面其2年的地径和树高生长量均不足背风面生长量的80%,在迎风面的保存率仅为背风面的40.6%;拉关木生长较快,树体较粗壮,在迎风面个体生长量与背风面很接近,但在迎风面的保存率仅为背风面的43.7%.

表7 不同海藻分布区域红树植物的生长表现Tab.7 Growth performance of mangrove species in different marine algae areas
2.5 种植措施对红树植物生长的影响
2.5.1 种植规格对红树植物生长的影响 沙质地和泥质地条件下不同种植规格的红海榄幼树2年生时的长势均较好,保存率很高,基本全部存活.沙质条件下,不同种植规格的幼树2年生时地径和树高的生长量差异不明显;在泥质条件下,随着种植密度的增加,2年生时的地径生长量趋向于减低,树高的生长量趋向于增加(表8).

表8 不同种植规格的红海榄幼树的生长情况1)Tab.8 Growth performance of Rhizophora stylosa with different planting specifications
2.5.2 苗木种植形式对红树林幼树生长的影响在普通沙地条件下(滩面高度为0),以胚轴苗形式种植的幼树生长量和保存率要高于以实生苗形式种植的情况.由表5 可知,以胚轴苗形式种植的红海榄2年生幼树的地径和树高生长量分别达到以实生苗形式种植的2.1 和2.2 倍,保存率达到以实生苗形式种植的3.7 倍.调查发现,胚轴在插植1 周后即可从基部萌发出幼根.这表明沙地条件有利于胚轴的生根和发芽,能有效为幼苗供应水分和养分.
在泥质条件下,以胚轴苗和实生苗种植的幼树生长差异性要远小于沙质的立地条件.由表6 可知,在普通泥质条件下(滩面高度为0.2 m),以胚轴苗形式种植的红海榄幼树2年生的地径生长量与以实生苗形式种植的无显著差异,树高生长量比以实生苗种植的高36.3%,但保存率略低于以实生苗种植的幼树的情况.在泥质高滩地条件下(滩面高度为0.4 m),以胚轴苗形式种植的红海榄幼树2年生的地径生长量略低于以实生苗形式种植的情况,树高生长量与以实生苗形式种植的无显著差异,保存率也与以实生苗形式种植的情况较接近.
可见,在沙地条件下,以胚轴苗与实生苗形式种植的幼树生长和成活表现出极显著的差异性,以胚轴苗形式种植的幼树生长量和保存率远高于以实生苗形式种植的情况.而在泥质地条件下,以胚轴苗和实生苗形式种植的幼树生长量和保存率的差异性不太明显.
2.5.3 胚轴插植深度对红树林幼树的影响 由表9可知,以红海榄胚轴长度的1/3、1/2 和2/3 这3 种不同插植深度作对比试验,在种植1年后,胚轴插植深度对幼树成活及生长的影响较小.由于胚轴插植初期,在尚未生根的情况下,若造林地出现较大的风浪,胚轴易被海水卷起而流失,因此,在种植胚轴苗时,胚轴应尽量深植于泥滩中,种植的深度以不超过胚轴长度的2/3 为宜.

表9 胚轴插植深度对1年生红海榄幼树的影响Tab.9 Effects of planting depth of hypocotyl on the growth of Rhizophora stylosa saplings
3 讨论与结论
3.1 滩面高度对红树植物幼树生长的影响
在高盐度海滩,滩面高度对红树植物的生长有显著的影响.无论是在沙质土壤上,还是在泥质土壤上,试验树种均是在中等的滩面高度即平均海平面附近时,具有最大的生长量和保存率,当滩面高度低于或高于此高度水平时,生长量和保存率均表现出下降的趋势.但与对保存率的影响相比,滩面高度对生长量的影响更为显著.以红海榄为例,当滩面高度大于0.4 m 时,在沙质和泥质土壤上其地径和树高生长量低于其最大生长量的50%,而保存率均高于其最大保存率的75%.红树林生长在海岸潮间带,当滩面过高时,海水的淹浸时间过短,不能充分满足红树植物对水分的要求,所以生长量会有所下降.另一方面,当滩面水平过低时,红树植物的海水淹浸时间过长,造成其在厌氧环境中时间过长,呼吸作用受到抑制,因此生长量随之受到影响.此外,保存率受滩面高度的影响较小,表明滩面高度并非红树植物成活的决定性因素.
3.2 土壤质地对红树植物幼树生长的影响
在高盐度海滩,土壤质地对红树植物生长的影响与滩面高度有关.在一定的滩面水平上,沙质和泥质条件下红海榄的生长量很接近,而在某些情况下,生长量的差异性又很明显.如在滩面高度为0 附近时,红海榄在沙质和泥质条件下生长量基本相同,地径接近1.4 cm,树高接近1.1 m;在滩面高度为0.2 m 时,红海榄在沙地条件的生长量明显高于泥质条件下的情况;但当滩面高度为0.4 m 时,红海榄的地径生长量在沙质条件下又低于泥质条件.但无论在何种滩面高度上,在沙质地和泥质地上红海榄的保存率都很接近,均在80%以上.由此可见,在生长定居初期(种植后2年内),滩面高度和土壤质地这2 个条件中,滩面高度对红树植物的生长影响更明显,而土壤质地对红树植物的生长影响未表现出确定性.滩面高度的差异性直接决定水分的供应,可见,水分条件是影响红树植物生长的关键因素.土壤质地不同,其养分含量是有差异性的,而由前述研究结果可知,养分含量并非红树植物定居生长的决定因素.
3.3 不同树种的生长差异性
在高盐度生境中,沙质高地上无瓣海桑保存率很低,仅为21.7%,拉关木、红海榄和秋茄的保存率均高于70%.从生长量上看,无瓣海桑和拉关木同为速生树种,但无瓣海桑的地径和树高生长量仅为拉关木的30%和50%左右,接近慢生树种红海榄和秋茄的生长量,可见,在沙质高地上,无瓣海桑的适应性最差.在普通沙地条件下,红海榄(以胚轴苗种植)、拉关木和无瓣海桑的保存率均高于80%,但红海榄(以实生苗种植)和秋茄的保存率均低于45%,生长量也很低,表明在普通沙地条件下慢生树种以实生苗形式种植的幼树生长适应性很差.在泥质高滩地上,拉关木和无瓣海桑的保存率均低于40%,在普通泥质滩地上的保存率均低于20%,表明这2 个树种在泥质滩地上的生长适应性都很差.
3.4 种植材料对红树植物幼树生长的影响
在高盐度海滩沙地条件下,红海榄胚轴苗的生长量和保存率远高于实生苗的种植情况,在泥质条件下,以胚轴苗和实生苗形式种植的红海榄幼树生长量和成活率的差异性不明显.这表明与以实生苗种植的幼树相比,以胚轴苗形式种植的幼树更适合于在沙地条件下生长.胚轴苗采集简单易行,而且不需要培育可直接插植.所以,在沙地条件下种植胚轴苗既能提高成活率和生长量,又能减少造林的成本.
3.5 林地海藻对红树植物幼树生长的影响
红树植物的生长除受到土壤质地和淹浸条件等影响外,在盐度过高的海滩上还易受到冬春季节近岸水体中海藻的影响.由于海藻的滋生速度快,又极难清除,特别是在退潮后,滞留的大量海藻会埋压幼苗幼树.通过观察发现,海藻对红树植物幼树的影响与风浪大小有关,当风浪较大时,海藻在水体中的摇曳会对红树植物幼树造成严重危害.而在风浪小的地方,海藻的流动性小,对幼树的影响小.由于生长较快的树种在短期内能形成较大的树体,对海藻埋压的抵抗力较强,因此在红树植物种植初期,海藻对生长较慢的低矮幼树影响较大,对生长较快的种类影响较小.拉关木生长比红海榄快,所以受海藻的影响较小.
3.6 与一般盐度条件下的生长比较
在与本试验区接近的深圳湾,淤泥深厚肥沃,海水盐度一般情况下低于15‰,有研究表明2.5年生的无瓣海桑的平均树高为5.3 m,平均地径为7.9 cm[31];3年生的秋茄树高1.20 m,地径3.01 cm[32].本研究中高盐度海滩普通泥质地条件下,2年生无瓣海桑树高1.14 m,地径1.63 cm,通过比较不同生境条件下的年均生长量可以发现,无瓣海桑在高盐度海滩的树高和地径生长速度仅为一般盐度条件下的26.9%和25.8%.与无瓣海桑的情况相似,在本研究中高盐度海滩普通泥质地条件下2年生秋茄树高和地径分别为0.38 m 和0.59 cm,树高和地径生长速度仅为一般盐度条件下的47.5%和29.4%.这充分表明过高的海水盐度对红树植物的生长具有一定的抑制作用,红树植物在高盐度条件下的生长速度远低于一般盐度条件下的生长情况.
[1]陈玉军,廖宝文,黄勃,等.秋茄(Kandelia obovata)和无瓣海桑(Sonneratia apetala)红树人工林消波效应量化研究[J].海洋与湖沼,2011,42(6):764-770.
[2]陈玉军,廖宝文,李玫,等.无瓣海桑和秋茄人工林的减风效应[J].应用生态学报,2012,23(4):959-964.
[3]MACINTOSH D J,ASHTON E C,HAVANON S.Mangrove rehabilitation and intertidal biodiversity:A study in the ranong mangrove ecosystem,Thailand[J].Estuar,Coast Shelf Sci,2002,55(3):331-345.
[4]WALTERS B B.Local mangrove planting in the philippines:Are fisherfolk and fishpond owners effective restorationists?[J].Rest Ecol,2000,8(3):237-246.
[5]ELLISON A M.Mangrove restoration:Do we know enough?[J].Rest Ecol,2000,8(3):219 –229.
[6]FIELD C D.Mangrove rehabilitation:Choice and necessity[J].Hydrobiologia,1999,413(1):47-52.
[7]TOLEDO G,ROJAS A,BASHAN Y.Monitoring of black mangrove restoration with nursery-reared seedlings on an arid coastal lagoon[J].Hydrobiologia,2001,444(1/2/3):101-109.
[8]KATON B M,POMEROY R S,GARCES L R,et al.Rehabilitating the mangrove resources of Cogtong Bay,Philippines:A comanagement perspective[J].Coast Manag,2000,28(1):29-37.
[9]IMBERT D,ROUSTEAU A,SCHERRER P.Ecology of mangrove growth and recovery in the Lesser Antilles:State of knowledge and basis for restoration projects[J].Rest Ecol,2000,8(3):230-236.
[10]LU Changyi,LIN Peng.The techniques of afforestation the mangrove and its role in coastline protection[C]∥WANG Ying.Island environment and coast development.Nanjing:Nanjing University Press,1993:321-328.
[11]莫竹承,梁士楚,范航清.广西红树林造林技术的初步研究[C]∥范航清,梁士楚.中国红树林研究与管理.北京:科学出版社,1995:164-172.
[12]刘治平.深圳福田红树林生态造林方法技术研究[J].生态科学,1995(2):100-104.
[13]莫竹承,范航清.红树林造林方法的比较[J].广西林业科学,2001,30(2):73-76.
[14]廖宝文,郑德璋,郑松发,等.红树植物秋茄造林技术的研究[J].林业科学研究,1996,9(6):586-592.
[15]廖宝文,郑德璋,郑松发,等.红树植物桐花树育苗造林技术的研究[J].林业科学研究,1998,11(5):474-480.
[16]廖宝文,李玫,陈玉军,等.中国红树林恢复与重建技术[M].北京:科学出版社,2010.
[17]郑松发,郑德璋,廖宝文,等.直接引进的乔木种群对原灌木群落及其种群的扰动效应[J].林业科学研究,1996,9(3):246-254.
[18]郑松发,郑德璋,廖宝文,等.不疏伐的灌木环境对乔木种群的适宜度及群落改造决策[J].林业科学研究,1998,11(3):289-294.
[19]郑德璋,廖宝文,郑松发,等.红树林主要树种造林与经营技术研究[M].北京:科学出版社,1999.
[20]陈玉军,陈文沛,郑松发,等.广东番禺红树林造林研究分析[J].生态科学,2001,20(1.2):25-31.
[21]陈玉军,廖宝文,彭耀强,等.红树植物无瓣海桑北移引种研究[J].广东林业科技,2002,19(2):9-12.
[22]陈玉军,郑松发,廖宝文,等.珠海市淇澳岛红树林引种扩种问题的探讨[J].广东林业科技,2002,18(2):31-36.
[23]吴中亨,蔡俊欣,叶亲柏.无瓣海桑引种及推广效果分析[J].广东林业科技,2000,16(2):6-10.
[24]李建清,徐何方,叶丽珍,等.秋茄红树林北移引种造林技术[J].浙江林业科技,2001,21(6):51-53.
[25]王伯荪,廖宝文,王勇军,等.深圳湾红树林生态系统及其持续发展[M].北京:科学出版社,2002.
[26]陈长平,王文卿,林鹏.盐度对无瓣海桑幼苗的生长和某些生理生态特性的影响[J].植物学通报,2000,17(5):457-461.
[27]王文卿,林鹏.盐度对红树植物木榄生长的影响[J].厦门大学学报:自然科学版,1999,38(2):273-279.
[28]莫竹承,范航清,何斌源.海水盐度对两种红树植物胚轴萌发的影响[J].植物生态学报,2001,25(2):235-239.
[29]诸姮,胡宏友,卢昌义.盐度对药用红树植物老鼠簕种子萌发和幼苗生长的影响[J].厦门大学学报:自然科学版,2008,47(1):131-135.
[30]郑文教,林鹏.盐度对红树植物海莲幼苗的生长和某些生理生态特性的影响[J].应用生态学报,1992,3(1):9-14.
[31]李云,郑德璋,陈焕雄,等.红树植物无瓣海桑引种的初步研究[J].林业科学研究,1998,11(1):39-44.
[32]廖宝文,郑德璋,郑松发,等.红树植物秋茄造林技术的研究[J].林业科学研究,1996,9(6):586-592.
