Research Progress on Nitrogen Use and Plant Growth
2014-07-02CuiXinYanQingweiSunJialinXiaoShuangXieFuchunandChenYajun
Cui Xin, Yan Qing-wei, Sun Jia-lin, Xiao Shuang, Xie Fu-chun, and Chen Ya-jun*
1 College of Animal Sciences, Northeast Agricultural University, Harbin 150030, China
2 College of Horticulture, Northeast Agricultural University, Harbin 150030, China
Introduction
Nitrogen (N)is one of the macronutrients required by plant growth, and the uptake of available N in the soil is an important indicator of plant growth. Soil N is also a determining abiotic factor for the plant growing in nutrient-def i cient soils (Chen et al., 2002).Large amounts of N fertilizer are applied to achieve maximum yield in agriculture production. N fertilizers have been exclusively used in China since 1970s,especially in some agriculture-intense areas (Zhu et al., 2010). However, N use efficiency (NUE)in China is 10%-20% lower than that of some developed countries (Ju and Zhang, 2003). On the other hand, the over-fertilization causes environmental problems, such as nitrate contamination and eutrophication of water bodies. The annual application of N is 25 000 000 tons, which is three times as the average application rate globally. Thus, promoting N use efficiency(NUE)becomes urgent for controlling their possible pollution to the ecosystem (Wu and Zhao, 2010).We reviewed the advance in current research related to the relationship between N and plant growth, and hopefully it could provide a reference source for the future studies about this topic.
N and Its Importance
N and its forms
Nitrogen is one of the essential macronutrients for plant growth and development (Wu and Zhao, 2010).N amount in plants ranks as the fourth largest one,which is after C, O and H.
Nitrogen in biosphere exists in three forms primarily: gaseous N in the air, organic and inorganic N pools in soils. Although N2accounts for 78% in the air,they can be absorbed by a small population of plants(leguminous plants and some other plants with root nodules)only through a process called bio N fi xation(Zhang, 2007). The organic N pool in soils consists of the residues and compost of animal bodies, waste,plants, and microorganisms. Besides, some N-rich amino acids, amide, and urea, most of the organic N is hydrophobic and cannot be easily dissolved into soil solution to supply the plant growth (Wang, 1999).Inorganic N pool that accounts about 1%-2% of the total N in the soil contains nitrate and ammonium mainly. Some other transitional forms like nitrite,nitrous and nitride N are also contained but at very low level. The inorganic N has high solubility, smaller molecule size, high mobility, and polarity. These characters make them a high availability for plants in soils, and therefore become the main N source for the plant growth (Zhang and Xu, 2011).
Nitrogen fertilizers in agricultural production are categorized by N groups contained in the molecules.Some commonly once used were listed in Table 1(Shen, 2001).

Table 1 N fertilizers categorized by N groups
Importance of N
Nitrogen is one of the major components of proteins,nucleic acids, lipid and some hormones in plants.N contributes 40%-50% of crop yield (Lu and Hu,2006), meanwhile, it affects the plant life cycle by assembling or regulating the cytoplasm, nuclear and membrane system using these molecules. Nitrogen is involved in the synthesis of some enzymes, coenzymes, and prosthetic groups (NAD+, NADP+, and FAD+). Therefore, N level has an immediate effect on cell division and growth in plants (Tian et al., 2011).
Besides, the photosynthesis is affected by N level in plants. Some researches pointed out that photosynthetic rate was increased with N level within a certain range in plants. When a critical level reached,any further increments in N would cause a decreasing photosynthetic rate (Evans, 1983). Some other researchs reported that N in plants played an important part in the anti oxidase activities (Li et al., 1997).N level is related to plant growth and development by affecting the water physiology in plants (Wu and Zhao, 2010). N level is proportional to that of ABA in plants, thus involving in the regulation of plant stomata activity. Optimal N level in plants could depress ABA level, sustain the stomata aperture, and increase the photosynthetic rate and yield (Garcia and Lamattia,2002).
Besides, nitrogen is important in improving WUE(water use efficiency)and soil water use. Possible mechanisms to enhance WUE by improving N nutrition in crop plants are presented in Fig. 1 (Waraich et al., 2011).
N Metabolism Related to Plant Growth and Development
The morphology, especially the root structure, has been of hot concern in uptake and utilization of N by plants under water deficiency. Root system plays an important role in the uptake of water and nutrients, synthesis of plant hormones, and the storage of photosynthetic assimilates. Root length,depth, thickness, density, weight, root to shoot ratio,penetration, osmotic adjustment are all good indicators used for related researches (Luo and Zhang, 2001).Cheng et al. (2007)found that the total root length,density, surface area, and biomass were significantly higher in rice cultivars which have higher NUE than those with low NUE and therefore could be used as reliable indicators in high NUE rice cultivar selection.Ye et al. (2010)compared the rice cultivars with different NUE and found that the cultivars with high NUE had higher total root length, surface area, and amount of root apexes than those with low NUE at the full heading stage. Gu et al. (2011)investigated the root distribution of some rice cultivars under varied water and N conditions and found that the root growth and distribution could be optimized by adjusting N application and irrigation regime. Wang et al. (2011)repored that root vigor tended to be decreased by water stress to a higher extent in plants with low NUE compared with those with high NUE on root characteristics, N, and water use efficiency in maize. The morphology and spatial distribution of maize cultivars with high NUE are more optimized for having deeper growth, which was beneficial for uptaking water and nutrients from soil and delaying the aging process. The root dry weight of maize treated with N fertilizer is higher than the control regardless of water conditions. The increase of shoot biomass is even more evident in N treated plants. A lower root to shoot ratio causes a greater water loss through the leaf respiration and impairment in water balance in crops (Zhang and Shan, 1995). Increasing root density and length are considered as a low energy demanding strategy to obtain water and N under water stress by plants (Hofer, 1986). Postma and Lynch (2011)indicated that the aerenchyma in root enhanced the growth potential of maize in low N condition. Nitrogen absorption by plants is related to the water conductivity of root-shoot system. Under drought condition, N uptake was regulated by lowering the root-shoot water conductivity to sustain high water use efficiency(Radin and Eidenbock, 1982). The respiration rate of root system is enhanced by N supply. Roots of plants with suff i cient N supply have higher respiration rates compared with those having N def i ciency under water stress (Xu, 2010). Unlike water use efficiency of wheat is improved by N addition, the improvement in NUE can only be obtained with low N supply (Li et al., 2003).
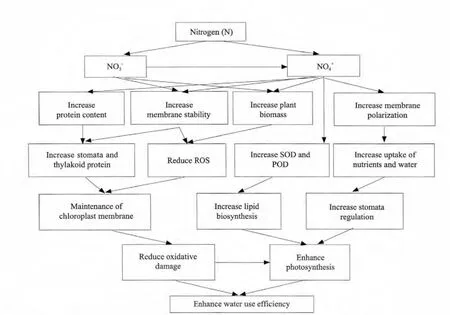
Fig. 1 Possible mechanisms through which nitrogen improves water eff i ciency in crop plants
The relationship the aerial parts of plants water and N was studied in different crops as well. Zhou et al.(2006)found that the tillering rate was accelerated by applying N under water stress. Different N forms and application rates affected the morphology of aerial parts, for example, plant type, leaf shape, leaf orientation, biomass, and yield (Cai et al., 2004).
The current researches related to N fertilizers and turfgrass mainly focused on N leachate, absorption, application rate and turf quality (Subhrajit et al., 2007). Poa spp. have varied N absorption ability under regular water condition; nitrogen adsorption ability based on root weight is proportional to NUE reversely; nitrogen adsorption ability of Poa spp.mowed routinely is positively correlated to leaf length,surface area, and root length (Jiang and Michael,2000). The impacts of water and N stress happened simultaneously on turf grass growth has barely been reported. Sun et al. (2010)pointed that the plants treated by high N rate were taller than the control in a research with Zoysia japonica under the same drought condition.
Physiological Metabolism of Plant Nitrogen
Many studies were conducted on plant nitrogen absorption, transformation, and physiology mechanisms improvement since 1980s (Bushoven and Hull, 2001).There were also many researches on physiological study of nitrogen metabolism with different water and nutrient contents. Physiological indicators, such as N content of enzyme activity, photosynthesis parameters,respiration parameters and osmoregulation substances,were used to test and evaluate the ability of plants to absorb and transform nitrogen. Nowadays, researchers focus on the function of nitrogen metabolism on enzyme regulation mechanism in plants (Dong, 2010).Nitrogen absorbed from plant roots was reduced to ammonia by nitrate reeducate (NR)and nitrite reeducate (NiR)in cytoplasm, continued to be reduced to glutamate and glutamine by glutamate synthase(GOGAT)and glutamine synthase (GS), then to aspartate (Asp)and asparagine (Asn), and finally to different amino acids and proteins for plant growth. The enzymes above were key enzymes for plant nitrogen metabolism, and their functions on physiological regulation function of N metabolism were investigated a lot (Yang et al., 2011). NR which is the fi rst enzyme in the nitrogen uptake assimilation process was concerned a lot. Researches showed that water stress reduced plant NR activity, affected N uptake and reduced N content. The content and activity of NR were different in different plants, and also at different growth periods and sites, and they were varied with environmental factors. Reasonable N application could improve NR activity, promote plant growth, improve photosynthesis rate and plant water use eff i ciency (Waraich et al., 2011).
Ribulose bisphosphate carboxylase (Rubisco),which was another important enzyme in N metabolism maintenance and distribution, was also investigated.This enzyme as an important nitrogenous compound in plants (occupying more than 50% of soluble protein in leaves)played an important role in adjusting the balance between carbon and nitrogen, especially in transforming between carbon and nitrogen in dry conditions. The decline in the activity of Rubisco would reduce the nitrogen content of leaves (Cheng and Fuchigami, 2000). Malic acid type of plant metabolism (NADP-ME)had higher nitrogen use efficiency than that of aspartate-type plants (NADME), which was indicated by lower content of N and soluble protein in leaves and was related with the high transformation rate of Rubisco (Górny, 2001).
Molecular Mechanism of Plant Nitrogen Metabolism
-concentration was low, so investigation on NRT2 became the focus of the study on plant nitrogen absorption and application (Wu and Zhao, 2010).Gene that was related with ammonium-nitrogen was a family member of AMT, and played an important role in plants absorbing nitrogen from soil. Glutamine synthetase gene and glutamate synthase gene expression were inf l uenced by different environmental factors, and amino acid synthesis required coupling of those two and involvement of various amino acid aminotransferases (Wang, 2011). Glutamate synthase gene mainly adjusted proline synthesis in phloem as a main metabolite for N source during drought stress. The excess expression of this gene in plants can improve the content of soluble protein, total amino acid, and total N in leaves, which are benef i cial for plant stress tolerance (Cai et al., 2009). QTL gene mapping showed that GS on N accumulation,transformation and nitrogen use eff i ciency (NUE)was co-adjusted by many representative gene components.GS gene played an important role in N allocation and transportation in many rhizomatous grasses(Swarbreck et al., 2011). Two glutamine synthetase genes and three aquaporin (TIPs)genes were isolated from ryegrass (Lolium perenne L.), where the glutamine synthetase gene was to control N absorption and transportation, and aquaporin (TIPs)gene was to influence N transportation through tonoplast into vacuole to store N (Nord-Larsen et al., 2009).
Therefore, there are 10 ammonium nitrogen absorption genes (AMT), two nitrate absorption genes, three GS genes, and three GOGAT genes related with N metabolism. But we should still focus on N absorption and transportation, while less on individual gene.
Prospects of Plant Nitrogen Metabolism Research
In recent years, many studies were conducted on plant N uptake, nitrogen use efficiency, and its transformation process and a series of new understanding were achieved. However, more studies are still required to further clarify the regulation mechanism of the nitrogen cycle in plants.
With the development of physiology, ecology,molecular biology, plant nutriology and their interactions, the understanding of nitrogen cycle mechanism will be further enhanced, and provide theoretical and practical basis for improving plant nutrient cycling, reuse and nitrogen use eff i ciency.
The future research may include selecting high nitrogen use efficiency varieties by studying the difference of nitrogen use efficiency among varieties and considering N requirement associated with morphology, growth property and cell physiological metabolism; further investigating key gene regulating mechanism related with nitrogen metabolism;studying nitrogen signal transduction pathways;improving nitrogen use eff i ciency and cultivating new varieties by transgenosis.
Bushoven J T, Hull R J. 2001. Nitrogen use efficiency is linked to nitrate reductase activity and biomass partitioning between roots and shoots of perennial ryegrass and creeping bentgrass. International Turf grass Society Research Journal, 9: 245-252.
Cai H M, Zhou Y, Xiao J H, et al. 2009. Over expressed glutamine synthetase gene modifies nitrogen metabolism and abiotic stress responses in rice. Plant Cell Rep, 28: 527-537.
Cai W D, Liu Y P, Li H Q, et al. 2004. Effects of application of nitrogenous fertilizer on the short-dated cotton of top removal planting type. Chinese Journal of Eco-Agriculture, 12(3): 92-94.
Chen C L, Liao L P, Wang S L, et al. 2002. Effect of phenolics on 15N nutrient absorption and distribution of Cunninghamia lanceolata.Journal of Plant Ecology, 26(5): 525-532.
Cheng J F, Dai T B, Jing Q, et al. 2007. Root morphological and physiological characteristics in relation to nitrogen absorption eff i ciency in different rice genotypes. Acta Pedologica Sinica, 44(2):266-272.
Cheng L L, Fuchigami L H. 2000. Rubisco activation state decreases with increasing nitrogen content in apple leaves. Experimental Botany, 51(351): 1687-1694.
Dong F R. 2010. Analysis of activities and characterization of the gene expression of key nitrogen metabolism enzymes in rice genotype with different N use eff i ciency. Yangzhou University, Yangzhou.
Evans R J. 1983. Nitrogen and photosynthesis in the fl ag leaf of wheat(Triticum aestivum Linn.). Plant Physiology, 72(2): 297-302.
Garcia M C, Lamattia L. 2002. Nitric oxide and abscisic acid cross talk in guard cells. Plant Physiology, 128: 790-792.
Górny A G. 2001. Variation in utilization efficiency and tolerance to reduced water and nitrogen supply among wild and cultivated barleys. Kluwer Academic Publishers, 117: 59-66.
Gu D X, Tang L, Xu Q J, et al. 2011. Root growth and distribution in rice cultivars as affected by nitrogen and water supply. Journal of Plant Ecology, 35(5): 558-566.
Hofer R M. 1986. Root hair formation and elongation of primary maize roots. Physiologic Plant arum, 68(4): 653-656.
Jiang Z C, Michael S W. 2000. Nitrate uptake of seeding and mature Kentucky bluegrass plants. Crop Science, 44(2): 567-574.
Ju X T, Zhang F S. 2003. Thinking about nitrogen recovery rate.Ecology and Environment, 12(2): 192-197.
Li F S, Kang S Z, Zhang J H, et al. 2003. Effects of atmospheric CO2enrichment, water and applied nitrogen on water-and nitrogen-use eff i ciencies of wheat. Plant and Soil, 254: 279-289.
Li X L, Gao D S, Gu M R, et al. 1997. Effects of ammonium and nitrate nitrogen on the activities of superoxide dismutase and peroxidase of apple trees. Plant Physiology Communications, 33(4): 254-256.
Lu J L, Hu A T. 2006. Plant nutrition. Higher Education Press, Beijing.pp. 23-25.
Luo L J, Zhang Q F. 2001. The status and strategy on drought resistance of rice (Oryza sativa L.). Chinese J Rice Sci, 15(3): 209-214.
Nord-Larsen P H, Kichey T, Jahn T P, et al. 2009. Cloning, characterization and expression analysis of tonoplast intrinsic proteins and glutamine synthetase in ryegrass (Lolium perenne L.). Plant Cell Rep, 28: 1549-1562.
Orsel M, Krapp A, Daniel-Vedele F. 2002. Analysis of the NRT2 nitrate transporter family in Arabidopsis structure and gene expression.Plant Physiology, 129: 886-896.
Postma J A, Lynch J P. 2011. Root cortical aerenchyma enhances the growth of maize on soils with suboptimal availability of nitrogen,phosphorus, and potassium. Plant Physiology, 156: 1190-1201.
Radin J W, Eidenbock M P. 1982. Water relations of cotton plant under nitrogen eff i ciency III. Plant Physiol, 67: 115-119.
Saha S K, Trenholm L E. 2007. Effect of fertilizer source on nitrate leaching and St. augustine grass turf grass quality. Hort Science,42(6): 1478-1481.
Shen Q R. 2001. The general theory of soil and fertilizer. Higher Education Press, Beijing. pp. 241-252.
Sun M, An Y, Wang Q, et al. 2010. Effect of water stress and nitrogen application on morphological and physiological character of Zoysia japonica cv. Shanghai. Pratacultural Science, 27(9): 57- 63.
Swarbreck S M, Defoin Platel M, Hindle M, et al. 2011. New perspectives on glutamine synthetase in grasses. Journal of Experimental Botany, 62(4): 1511-1522.
Tian J, Guo S R, Sun J, et al. 2011. Effects of exogenous spermidine on nitrogen metabolism of cucumber seedlings under high temperature stress. Chinese Journal of Ecology, 30(10): 2197-2202.
Wang H, Lnukai Y, Yamauchi A. 2006. Root development and nutrient uptake. Critical Reviews in Plant Sciences, 25(3): 279-301.
Wang J F. 2011. Effects of irrigation and nitrogen coupling on root characteristics, nitrogen and water utilization in different summer corn genotypes. Shandong Agricultural University, Tai'an.
Wang P. 2011. Genotypic differences in nitrogen metabolism and the analysis of physiological mechanism. Huazhong Agricultural University, Tai'an.
Wang Z. 1999. Plant physiology. China Agricultural Press, Beijing. pp.80-110.
Waraich E A, Ahmad R, Ashraf M Y, et al. 2011. Improving agricultural water use efficiency by nutrient management in crop plants. Acta Agricultural Scandinavica, Section B-Soil & Plant Science, 61(4): 291-304.
Wu W, Zhao J. 2010. Advances on plants' nitrogen assimilation and utilization. Chinese Agricultural Science Bulletin, 26(13): 75-78.
Xu J D. 2010. Effects of soil water and nitrogen phosphorus nutrition on the growth of plant roots. Soil and Water Conservation Science and Technology in Shan'xi, 3: 12-15.
Yang J, Shi Y W, Li Z W, et al. 2011. Research progress of carbon and nitrogen metabolism of rice. Crop Research, 25(4): 383-387.
Ye L T, Fan J B, Xu Y H, et al. 2010. Characteristics of growth in rice genotypes with different nitrogen use eff i ciencys. Journal of Nanjing Agricultural University, 33(3): 77-81.
Zhang H Z. 2007. The identification of Dof transcription factors response to low nitrogen stress and the development of low-nitrogen tolerant transgenic rice. Huazhong Agricultural University, Wuhan.
Zhang H Z, Xu H Y. 2011. Research progress on the enzymes during plant nitrogen assimilation. Northern Horticulture, 20: 180-183.
Zhang S Q, Shan L. 1995. Effects of nitrogen nutrition on the drought adaptation and water use of spring wheat. Research of Soil and Water Conservation, 2(1): 31-35, 55.
Zhou Y, Guo S W, Song N, et al. 2006. Effects of nitrogen form and water stress interaction on photosynthesis, utilization of water and nitrogen of rice plants at the tillering stage. Chinese Journal of Rice Science, 20(3): 313-318.
Zhu Z L, Zhang S L, Yin B, et al. 2010. Historical comparison on the response curves of rice yield-nitrogen application rate in Tai Lake region. Plant Nutrition and Fertilizer Science, 16(1): 1-5.
杂志排行

Journal of Northeast Agricultural University(English Edition)的其它文章
- Farmer Labor Service Export from Heilongjiang Province to Russia
- Cultivation of Chinese Farmers' Integrity
- Regulation of Foliar Application DCPTA on Growth and Development of Maize Seedling Leaves in Heilongjiang Province
- Comparison of Physiological Properties Between Dwarf and Vinetype Cucumbers (Cucumis sativus Linn.)
- Effects of Substitute Media on Development of Potted Cyclamen percicum Mill.
- Expression of HSP72 in Mouse Preimplantation Embryos with Heat Shock