Human bone marrow mesenchymal stem cell transplantation attenuates axonal injury in stroke rats
2014-06-01YiXuShiweiDuXinguangYuXiaoHanJincaiHouHaoGuo
Yi Xu, Shiwei Du, Xinguang Yu, Xiao Han, Jincai Hou, Hao Guo
1 Department of Neurosurgery, General Hospital of Chinese PLA, Beijing, China
2 Department of Neurosurgery, General Hospital of Armed Police Forces, Beijing, China
3 Xiyuan Hospital, China Academy of Chinese Medical Sciences, Institute of Basic Medical Sciences, Beijing, China
4 Beijing Key Laboratory of Pharmacology of Chinese Materia Medica, Institute of Basic Medcal Sciences of Xuyuan Hospital, Beijing, China
Human bone marrow mesenchymal stem cell transplantation attenuates axonal injury in stroke rats
Yi Xu1, Shiwei Du2, Xinguang Yu1, Xiao Han3,4, Jincai Hou3,4, Hao Guo3,4
1 Department of Neurosurgery, General Hospital of Chinese PLA, Beijing, China
2 Department of Neurosurgery, General Hospital of Armed Police Forces, Beijing, China
3 Xiyuan Hospital, China Academy of Chinese Medical Sciences, Institute of Basic Medical Sciences, Beijing, China
4 Beijing Key Laboratory of Pharmacology of Chinese Materia Medica, Institute of Basic Medcal Sciences of Xuyuan Hospital, Beijing, China
Previous studies have shown that transplantation of human bone marrow mesenchymal stem cells promotes neural functional recovery after stroke, but the neurorestorative mechanisms remain largely unknown. We hypothesized that functional recovery of myelinated axons may be one of underlying mechanisms. In this study, an ischemia/reperfusion rat model was established using the middle cerebral artery occlusion method. Rats were used to test the hypothesis that intravenous transplantation of human bone marrow mesenchymal stem cells through the femoral vein could exert neuroprotective effects against cerebral ischemia via a mechanism associated with the ability to attenuate axonal injury. The results of behavioral tests, infarction volume analysis and immunohistochemistry showed that cerebral ischemia caused severe damage to the myelin sheath and axons. After rats were intravenously transplanted with human bone marrow mesenchymal stem cells, the levels of axon and myelin sheath-related proteins, including microtubule-associated protein 2, myelin basic protein, and growth-associated protein 43, were elevated, infarct volume was decreased and neural function was improved in cerebral ischemic rats. These fi ndings suggest that intravenously transplanted human bone marrow mesenchymal stem cells promote neural function. Possible mechanisms underlying these bene fi cial effects include resistance to demyelination after cerebral ischemia, prevention of axonal degeneration, and promotion of axonal regeneration.
nerve regeneration; human bone marrow mesenchymal stem cells; ischemic stroke; neural function; neuroprotection; microtubule-associated protein 2; myelin basic protein; growth associated protein 43; neuraxon; myelin sheath; demyelination; axon regeneration; neural regeneration
Xu Y, Du SW, Yu XG, Han X, Hou JC, Guo H. Human bone marrow mesenchymal stem cell transplantation attenuates axonal injury in stroke rats. Neural Regen Res. 2014;9(23):2053-2058.
Introduction
Cerebral ischemia is an important clinical neurological disorder, and there is considerable interest in protecting neurological function following cerebral ischemia. To date, the best method for treating such patients has been thrombolytic therapy in the acute period; however, few patients can receive this therapy because of the limited time window.
Mesenchymal stem cells have the potential to differentiate into various cell types, including osteoblasts, chondrocytes, adipocytes, and hepatocytes, as well as other tissue lineages including neurons, neural stem cells, microglial cells, and astrocytes (Deng et al., 2006). Mesenchymal stem cells are regarded as one of the most promising types of stem cells used for the treatment of experimental cerebral ischemic stroke to promote neurological function (Tang et al., 2007; Li et al., 2010; Bao et al., 2011). However, the neurorestorative mechanisms by which mesenchymal stem cells improve functional recovery remain largely unknown.
Myelinated axons are essential for the normal functioning of the brain. Increasing evidence using a variety of markers targeting different structures within axons has demonstrated the acute vulnerability of axons to cerebral ischemia in vivo (Dewar et al., 1997; Yam et al., 1998). Myelinated axons are susceptible to anoxia, and functional and structural impairment occurs within the axon itself after focal cerebral ischemia (Philippa et al., 2000).
In the present study, we assessed neurological status in ischemic model rats generated by transient middle cerebral artery occlusion (MCAO) following intravenous injection of human bone marrow mesenchymal stem cells (hBMSCs). Furthermore, we analyzed the alteration of axons and myelin sheath-associated proteins, such as microtubule-associated protein 2 (MAP-2), myelin basic protein (MBP), and growth-associated protein 43 (GAP-43), in these animals. This was done in an effort to show whether the mechanism by which transplantation of hBMSCs protects against cerebral ischemia is associated with the ability to attenuate axonal injury, such as demyelination.
Materials and Methods
Animals
Twenty adult male Sprague-Dawley rats, weighing 250—300 g, were purchased from Beijing Weitong Lihua Experimental Animal Technology Co., Ltd. (Beijing, China; certi fi cateNo. SCXK (Jing) 2012-0001). All studies were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Rats were maintained on a 12-hour light/dark cycle with free access to food and water.
hBMSC preparation and culturein vitro
Bone marrow aspirates were obtained by puncture of the posterior iliac crest of healthy human donors (Fang et al., 2003) after informed consent was obtained. Mononuclear cells in bone marrow specimens were separated on Percoll-Paque Hydroxypropylmethylcellulose (Dow, Beijing, China). Flk1+, CD34—, CD31—hBMSCs (the cell line was obtained under phase one clinical approval; approval No. 2004L04792, Food and Drug Administration, USA) were puri fi ed using immunomagnetic beads. Mononuclear cells were plated at a density of 1 × 106cells/25 cm2cell culture fl ask in 20 mL of culture medium containing 58% DMEM/ F12 (Gibco, Grand Island, NY, USA) + 40% MCDB-201 (Sigma, St. Louis, MO, USA), 2% fetal bovine serum (Hyclone, Grand Island, NY, USA), 10 ng/mL epidermal growth factor, 10 ng/mL platelet-derived growth factor, 2 mmol/L L-glutamine (Hyclone, USA) and 100 μg/mL penicillin, and cultured at 37°C in a humidified atmosphere containing 5% CO2. After 1 day, nonadherent cells were washed and removed. The medium was replaced with the same fresh medium every third day. Plastic-adherent hBMSCs were digested with 0.125% trypsinase and 0.01% EDTA upon reaching 80% confluence. hBMSCs at passage 3 were collected and used for transplantation. Nucleated marrow cells were counted using a fl ow cytometer to ensure adequate cell number for transplantation. hBMSCs were analyzed by fl ow cytometry (eFluor, eBioscience, San Diego, CA, USA) for phycoerythrin anti-human phycoerythrin-Flk-1, CD31 and CD34.
Establishing MCAO and reperfusion models
Sprague-Dawley rats (n = 20) were initially anesthetized with 3.5% chloral hydrate. We induced transient MCAO using an intraluminal vascular occlusion method (Koizumi et al., 1986; Nagasawa et al., 1989). In brief, rats were anesthetized with 3.5% chloral hydrate. Rectal temperature was maintained at 37°C throughout the surgical procedure using a heating lamp. The right common carotid artery, external carotid artery, and internal carotid artery were exposed. A length of 40 mm of mono fi lament nylon suture (0.24 mm diameter) was advanced from the external carotid artery into the lumen of the internal carotid artery until it blocked the origin of the middle cerebral artery. At 2 hours after MCAO, the nylon suture was withdrawn until the suture tip cleared the lumen of the external carotid artery to realize reperfusion.
Grouping
At 24 hours after MCAO, rats were evaluated for modified neurological severity score (mNSS), which is based on a series of motor (Borlongan et al., 1995), sensory (Markgraf et al., 1992), balance and re fl ex tests (Germano et al., 1994). Scores of 1—6 indicate mild injury, scores of 7—12 indicate moderate injury, and scores of 13—18 indicate severe injury. MCAO rats with an mNSS of 10—14 were randomly assigned to two groups: an MCAO + saline group (n = 6) and an MCAO + hBMSCs group (n = 6).
hBMSC transplantation
At 24 hours after MCAO and neurological assessment, rats were anesthetized as described above and received transplantation of hBMSCs or saline. The femoral vein was punctured under a microscope to ensure the needle was properly inserted. Approximately 1 × 106hMSCs in 10 μL of saline (Mahmood et al., 2003) were injected into the animals in the MCAO + hBMSCs group at a rate of 1 μL/min; an equal volume of saline was injected into animals in the MCAO + saline group at the same rate. The wounds were stitched up immediately after transplantation.
Behavioral testing
After MCAO surgery, all rats underwent two behavioral tests, an adhesive-removal somatosensory test and the mNSS test, before reperfusion, and at 1 day, 3 days, 1 week, 2 weeks, 3 weeks, and 4 weeks after reperfusion, by an investigator who was blinded to the experimental groups.
The adhesive-removal somatosensory test was applied to measure forelimb somatosensory asymmetries (Schallert et al., 1984). Small adhesive-backed paper dots (63.62 mm2) were used as bilateral tactile stimuli and applied to the radial aspect of the wrist of each forelimb on three trials per day in the home cage. The number of seconds at which the rat contacted and removed the stimuli was recorded. Individual trials were separated by at least 10 minutes. The animals were trained in the adhesive-removal dot test for 3 days prior to MCAO surgery. Once the rats were able to remove the dots within 10 seconds, they were subjected to MCAO.
The mNSS test is a composite of motor, sensory, balance, and re fl ex tests, used to grade various aspects of neurological function (Borlongan et al., 1995). Neurological function is graded on a scale of 0—18 (normal score = 0, maximal de fi cit score = 18). A single point is awarded for a speci fi c abnormal behavior or for the lack of a tested re fl ex. The higher the score is, the more severe the injury is.
Slice preparation
All rats were allowed to survive for 4 weeks after MCAO and reperfusion, and were used for morphologic analysis. Rats were deeply anesthetized with 10% chloral hydrate. The vascular system was transcardially perfused with heparinized PBS, followed by 4% paraformaldehyde. The brains were immersed in 4% paraformaldehyde in PBS for 24 hours. Then, the brain tissues were dissected into six 2-mm coronal blocks and embedded in paraf fi n. A series of adjacent 3-μm-thick slices were cut from each block.
Measurement of infarct volume
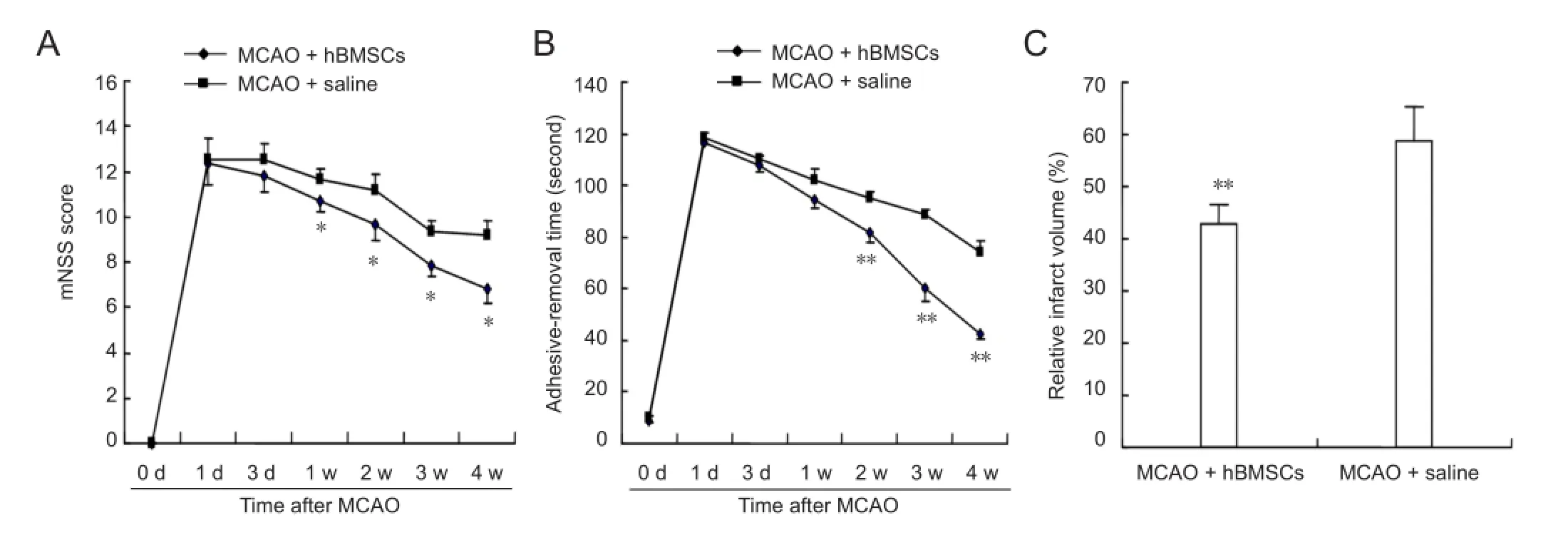
Figure 1 Results of behavioral functional tests and infarct volume in a rat model of middle cerebral artery occlusion (MCAO) after treatment with human bone marrow mesenchymal stem cells (hBMSCs) or saline.
The brain sections were stained with hematoxylin and eosin and photographed under a microscope (XDS-1B, Chongqing Optical Instrument Factory, Chongqing, China). Relative infarct volume was analyzed using the Image Pro Plus 6.0 analysis system (Media Cybernetics, Silver Spring, MD, USA). The indirect lesion area, in which the infarct area in the ipsilateral hemisphere was subtracted from the area of the contralateral hemisphere, was calculated. Relative infarct volume is presented as percent volume of the indirect lesion compared with the contralateral hemisphere (Swanson et al., 1990).
Immunohistochemical staining
Sections were processed for double-immunofluorescence staining for MAP-2/GAP-43 or MAP-2/MBP. Briefly, after blocking in 5% normal goat serum for 30 minutes at 37°C, slides were incubated at 4°C overnight in mouse anti-MAP-2 polyclonal antibody (1:10,000; Boehringer, Ingelheim am Rhein, Rhineland-Palatinate, Germany), and then treated with Cy3-conjugated sheep anti-mouse IgG (1:500; Chemicon, Temecula, CA, USA). This was followed by treatment with rabbit anti-GAP-43 polyclonal antibody (1:300; Abcam, Cambridge, MA, USA) or rabbit anti-MBP (1:500; polyclonal, Chemicon), followed by FITC-conjugated sheep anti rabbit IgG (1:50; Chemicon) staining.
To visualize the nuclei, the sections were counterstained with 6-diamidinao-2-phenyl-indole (DAPI; Dojindo, Kumamoto, Japan).
Negative control sections from each rat received identical preparation for immunohistochemical staining, except that the primary antibodies were omitted.
Immuno fl uorescence-labeled slides were examined under a confocal laser-scanning microscope (Nikon, Eclipse-8oi, Tokyo, Japan); green (FITC) and red (Cy3) fluorochromes on the slides were excited by the laser beam at 488 nm and 647 nm, and emissions were acquired sequentially with a photomultiplier tube through 522 nm and 670 nm emission fi lters.
Statistical analysis
Data are presented as the mean ± SD and were analyzed using SPSS 17.0 software (SPSS, Chicago, IL, USA). Oneway analysis of variance was used for statistical comparisons among groups. A P value of less than 0.05 was considered statistically signi fi cant.
Results
Growth of hBMSCs in vitro
The results of human BMSC characterization and karyotype analysis showed that the hBMSCs used in this study were positive for fetal liver kinase-1 (Flk-1) and negative for CD31 and CD34. hBMSCs at passage 5 were used for karyotype analysis, which was conducted to check chromosome stability. No trisomy, tetraploidy, or chromosome rearrangement was observed.
Neurological functional testing after hBMSC transplantation
The mNSS test and adhesive-removal dot test were applied to compare behavioral functions of rats between the hBMSC-treated and control groups. There was no difference in neurological functions before cell transplantation. At 1—4 weeks after MCAO, significant functional recovery was observed in the adhesive-removal dot test in the hBMSC-treated group (P < 0.05 or P < 0.01;Figure 1B), while neurological severity scores in mNSS test were obviously lower in the hBMSC-treated group compared with the control group (P <0.01;Figure 1A).
Infarct volume after hBMSC transplantation
By hematoxylin and eosin staining, the infarct was mainly seen in the cortex and striatum at 4 weeks after MCAO. The relative infarct volume in hBMSC-treated rats was significantly less than that in the saline-treated group (P < 0.01;Figure 1C).
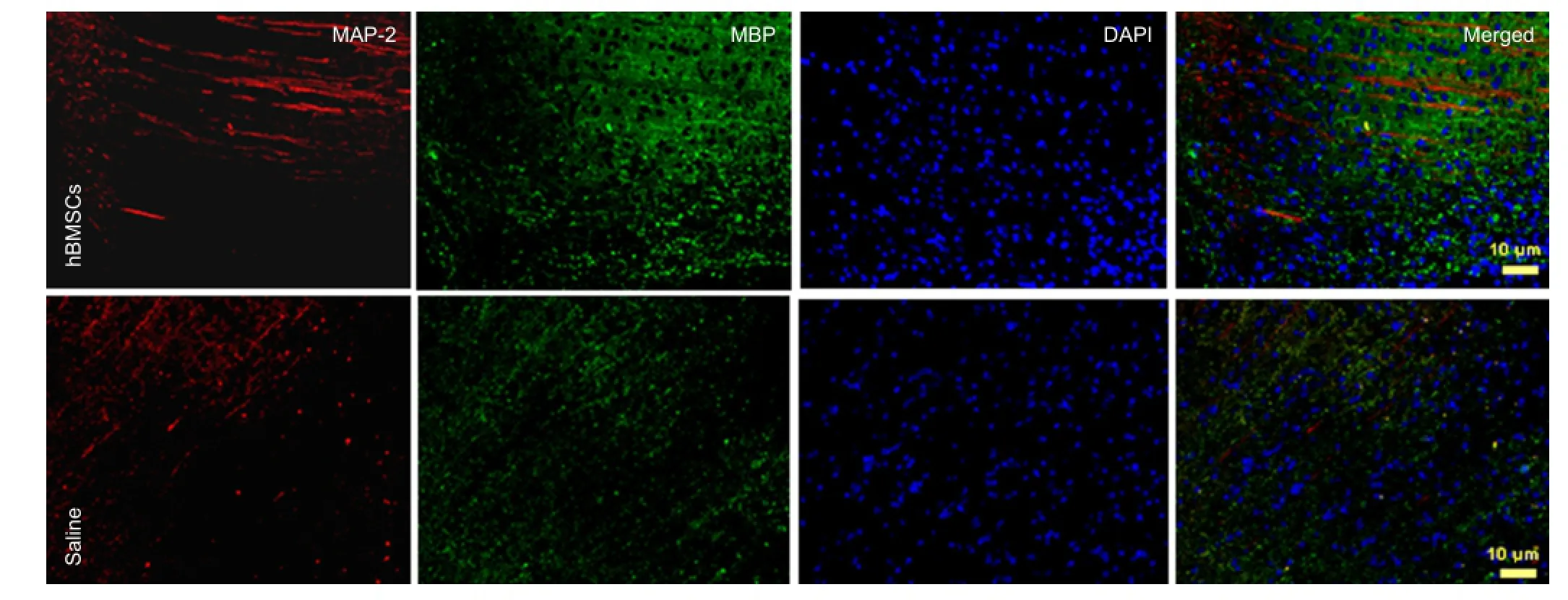
Figure 2 Double immuno fl uorescence labeling for MAP-2/GAP-43 in brain sections of hBMSC- or saline-treated rats at 4 weeks after MCAO (× 20).
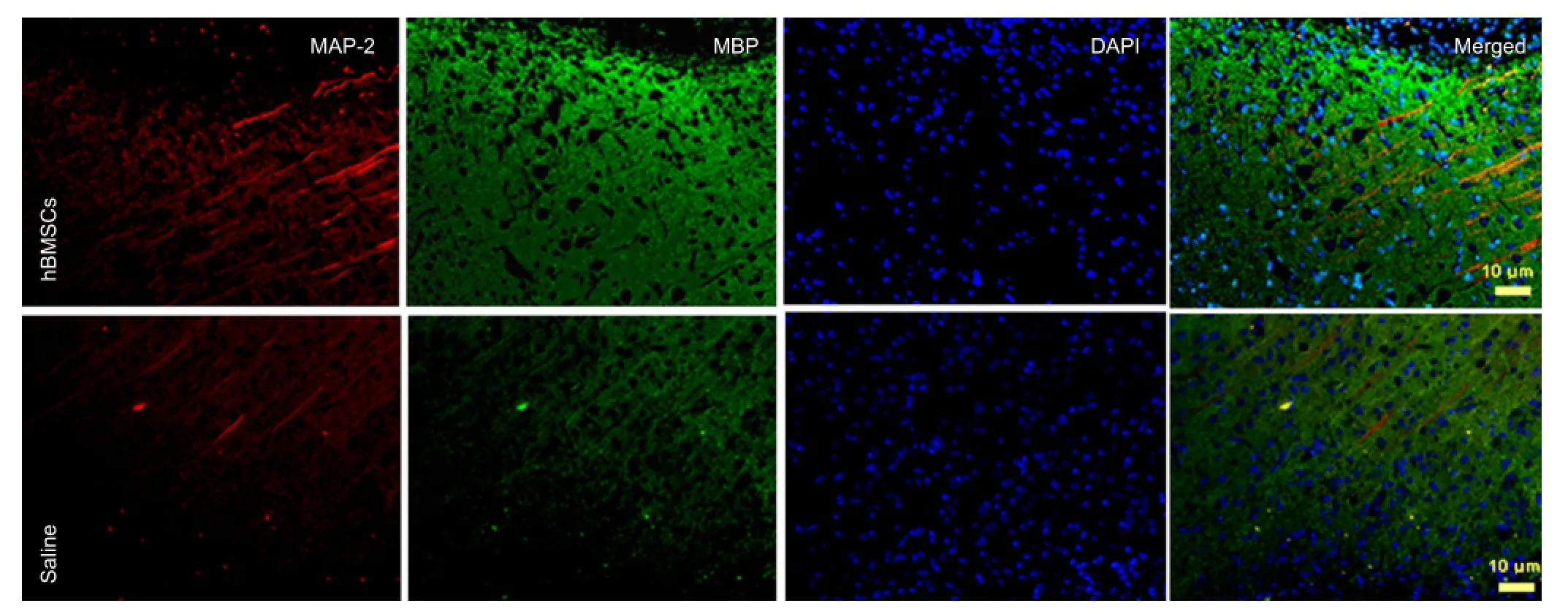
Figure 3 Double immuno fl uorescence labeling for MAP-2/MBP in brain sections of hBMSC- or saline-treated rats at 4 weeks of MCAO (× 20).
Double immunohistochemistry staining for MAP-2/ GAP-43, MAP-2/MBP in the infarct area of cerebral ischemic rats after hBMSC transplantation
MAP-2 fl uorescence showed the density and courses of axons around the infarct-affected cortex. GAP-43 was distributed in the neuropil, throughout axons. At 4 weeks after 2 hours of MCAO, MAP-2 and GAP-43 levels were decreased more signi fi cantly in the ischemic core of MCAO rats. MAP-2 was strongly expressed in the infarct area and was colocalized with the higher levels of GAP-43 in hBMSC-treated rats than in the saline-treated group. This indicated that hBMSCs could prevent axonal degeneration after cerebral ischemia as well as promote axonal regeneration (Figure 2).
In the hBMSC-treated group, the fluorescence for MBParound the infarction area displayed fasciculation at equal pace with axon course. The fasciculation distribution of MAP-2 and MBP were successive and integrated relatively than saline-treated group, which indicated that hBMSC transplantation could possibly prevent demyelination after cerebral ischemia (Figure 3).
Discussion
In the present study, we demonstrated that transplantation of hBMSCs into MCAO rats improved neurological function and reduced infarct volume. This was accompanied by elevated levels of the axon- and myelin sheath-related proteins MAP-2, MBP and GAP-43. Possible mechanisms underlying these beneficial effects include resistance to demyelination after cerebral ischemia, prevention of axonal degeneration, and promotion of axon regeneration.
Cerebral ischemia is a major cause of death and disability worldwide. Thrombolytic treatment is effective when administered within 4.5 hours after stroke (Shinozuka et al., 2013). However, only a few patients are able to receive treatment within this limited therapeutic window (Shinozuka et al., 2013). Furthermore, thrombolytic treatment may increase the risk of intracranial hemorrhage (Hacke et al., 2008).
There is increasing evidence that transplantation of BMSCs could represent an alternative therapy for ischemic stroke. The transplanted BMSCs signi fi cantly promote functional recovery after ischemic stroke in animal models (Ding et al., 2013) and in the clinic (Lee et al., 2010; Honmou et al., 2011). However, it is unclear what brings about the purported bene fi t from BMSC transplantation.
Previous studies have reported that BMSCs could differentiate into brain parenchymal cells and replace the injured neurons in the infarct area (Woodbury et al., 2000; Chen et al., 2001), which was regarded as the main goal of cell therapy (Woodbury et al., 2000; Chen et al., 2001). However, very few transplanted cells were found in the brain and, of these, only a small percentage of cells expressed neuronal cell markers (Chen et al., 2002; Shen et al., 2007). Moreover, after BMSC transplantation, these cells are very unlikely to have truly integrated into the parenchymal tissue and to have formed the complex connections that promote functional recovery (Chen et al., 2002).
Other studies have shown that BMSC transplantation can reduce cell apoptosis (Huang et al., 2013), induce angiogenesis (Guo et al., 2012), promote proliferation of endogenous cells (Bao et al., 2011), and induce the production of trophic and angiogenic factors (Chen et al., 2002; Zhang et al., 2005). We studied the anti-demyelination effects of hBMSC transplantation, and examined the effects of hBMSC transplantation on promoting the regeneration of axonsand myelin sheaths.
MAP-2, a marker of dendrosomatic neuronal injury, has been perceived primarily as a static, structural protein, being necessary along with other intracellular components, e.g., actin and neuro fi laments, to maintain the neuroarchitecture. Several studies have reported decreases in MAP-2 levels in the gerbil brain after ischemia (Matesic et al., 1994; Dawson et al., 1996). Loss of MAP-2 may participate in the initial phase of neuronal dysfunction, and dendritic breakdown may be a fi rst sign of neurodegeneration as early as 1 hour after cerebral ischemia (Dawson et al., 1996). Moreover, MAP-2 has roles in the growth, differentiation, and plasticity of neurons, with key roles in neuronal responses to growth factors, neurotransmitters, synaptic activity, and neurotoxins (Johnson et al., 1992). The results of these studies indicate that modi fi cation and rearrangement of MAP-2 is an early obligatory step in many processes that modify neuronal function.
GAP-43 (a 43-kDa growth-associated protein) is a nervous tissue-speci fi c protein, synthesized at high levels during axonal outgrowth during neuronal development and regeneration (Jacobson et al., 1986; Goslin et al., 1988; Stoemer et al., 1995). Axonal sprouting, a component of anatomic plasticity, can be identi fi ed by the elevated expression of GAP-43 (Skene et al., 1989; Buffo et al., 1997). An acute increase in the level of expression of GAP-43 in the cortex is observed after cortical ischemia (Stroemer et al., 1993; Stoemer et al., 1995) and in the substantia nigra after striatal ischemic injury in adult rats (Goto et al., 1994). These reports suggest that GAP-43 is a sensitive marker for the assessment of axonal damage and the regenerative response in the ischemic area.
MBP is a major myelin-associated protein located with the membranes of neurons. Changes in MBP levels are used as a diagnostic indicator of demyelination (Yam et al., 2000).
Our data indicate that treatment with hBMSCs decreased infarct volume and improved functional recovery relative to the saline-treated group 4 weeks after MCAO. In an effort to showthe role of axon- and myelin sheath-associated proteins in cerebral ischemia, we assessed the levels and colocalization of MAP-2/MBP and MAP-2/GAP-43 using double-staining immunohistochemistry. The results of the present study demonstrated that these proteins are preferentially expressed in the cerebral infarct zone in hBMSC-treated rats 4 weeks after MCAO, relative to the levels in saline-treated animals, suggesting a compensatory role for these proteins in this region. MAP-2 is sensitive to ischemia. The modi fi cation and rearrangement of MAP-2 contributed to the improvement of neuronal function (Li et al., 1998). The increase in the levels of MAP-2 in the transplantation group was associated with recovery of neuronal function, combined with the expression of MBP, an indicator of demyelination in cerebral ischemia, suggesting that neuronal functional recovery is related to this anti-demyelinating effect of hBMSCs. In the present study, the level of GAP-43, a sensitive marker for the assessment of axonal damage and regeneration, was also higher in the ischemic area in the hBMSC-treated group compared with the saline-treated group, suggesting a possible role for hBMSCs in axonal protection and regeneration.
Author contributions:Xu Y, Du SW and Yu XG designed the study and provided critical revision of the manuscript for intellectual content. Xu Y performed research, analyzed data and wrote the paper. Han X, Hou JC, Guo H gave technical and material support. All authors approved the final version of the manuscript.
Con fl icts of interest:None declared.
Bao X, Wei J, Feng M, Lu S, Li G, Dou W, Ma W, Ma S, An Y, Qin C, Zhao RC, Wang R (2011) Transplantation of human bone marrow-derived mesenchymal stem cells promotes behavioral recovery and endogenous neurogenesis after cerebral ischemia in rats. Brain Res 1367:103-113.
Borlongan CV, Randall TS, Cahill DW, Sanberg PR (1995) Asymmetrical motor behavior in rats with unilateral striatal excitotoxic lesions as revealed by the elevated body swing test. Brain Res 676:231-234.
Buffo A, Holtmaat AJ, Savio T, Verbeek JS, Oberdick J, OestreicherAB, Gispen WH, Verhaagen J, Rossi F, Strata P (1997) Targeted overexpression of the neurite growth-associated protein B-50/GAP-43 in cerebellar Purkinje cells induces sprouting after axotomy but not axon regeneration into growth-permissive transplants. J Neurosci 17:8778-8791.
Chen J, Li Y, Wang L, Lu M, Zhang X, Chopp M (2001) Therapeutic bene fi t of intracerebral transplantation of bone marrow stromal cells after cerebral ischemia in rats. J Neurol Sci 189:49-57.
Chen X, Li Y, Wang L, Katakowski M, Zhang L, Chen J, Xu Y, Gautam-SC, Chopp M (2002) Ischemic rat brain extracts induce human marrow stromal cell growth factor production. Neuropathology 22:275-279.
Dawson DA, Hallenbeck JM (1996) Acute focal ischemia-induced alterations in MAP-2 immunostaining: description of temporal changes and utilization as a marker for volumetric assessment of acute brain injury. J Cereb Blood Flow Metab 16:170-174.
Deng J, Petersen BE, Steindler DA, Jorgensen ML, Laywell ED (2006) Mesenchymal stem cells spontaneously express neural proteins in culture and are neurogenic after transplantation. Stem Cells 24:1054-1064.
Dewar D, Dawson DA (1997) Changes of cytoskeletal protein immunostaining in myelinated fi ber tracts after focal cerebral ischemia in the rat. Acta Neuropathol 93:71-77.
Ding X, Li Y, Liu Z, Zhang J, Cui Y, Chen X, Chopp M (2013) The sonic hedgehog pathway mediates brain plasticity and subsequent functional recovery after bone marrow stromal cell treatment of stroke in mice. J Cereb Blood Flow Metab 3:1015-1024.
Fang B, Shi M, Liao L, Yang S, Liu Y, Zhao RC (2003) Multiorgan engraftment and multilineage differentiation by human fetal bone marrow Flk1+/CD31-/CD34-Progenitors. Hematother Stem Cell Res 12:603-613.
Germano AF, Dixon CE, d’Avella D, Hayes RL, Tomasello F (1994) Behavioral de fi cits following experimental subarachnoid hemorrhage in the rat. J Neurotrauma 11:345-353.
Goslin K, Schreyer DJ, Skene JHP, Banker G (1988) Development of neuronal polarity: GAP43 distinguishes axonal from dendritic growth cones. Nature 336:672-674.
Goto S, Yamada K, Inoue N, Nagahiro S, Ushio Y (1994) Increase expression of growth-associated protein GAP-43/B-50 following cerebral hemitransection or striatal ischemic injury in the substantia nigra of adult rats. Brain Res 647:333-339.
Guo F, Lv S, Lou Y, Tu W, Liao W, Wang Y, Deng Z (2012) Bone marrow stromal cells enhance the angiogenesis in ischaemic cortex after stroke: involvement of notch signaling. Cell Biol Int 36:997-1004.
Hacke W, Kaste M, Bluhmki E, Brozman M, Davalos A, Guidetti D, Larrue V, Lees KR, Medeghri Z, Machnig T, Schneiger D, Kummer R, Wahlgren N, Toni D, the ECASS Investigators (2008) Thrombolysis with alteplase 3-4.5 hours after acute ischemic stroke. N Engl J Med 359:1317-1329.
Honmou O, Houkin K, Matsunaga T, Niitsu Y, Ishiai S, Onodera R, Waxman SG, Kocsis JD (2011) Intravenous administration of auto serum-expanded autologous mesenchymal stem cells in stroke. Brain 134:1790-1807.
Huang W, Mo X, Qin C, Zheng J, Liang Z, Zhang C (2013) Transplatation of differentiated bone marrow stromal cells promotes motor functional recovery in rats with stroke. Neurol Res 35:320-328.
Jacobson RD, Virag I, Skene JH (1986) A protein associated with axon growth, GAP43 is widely distributed and developmentally regulated in rat CNS. J Neurosci 6:1843-1855.
Johnson GV, Jope RS (1992) The role of microtubule-associated prtein 2 (MAP-2) in neuronal growth, plasticity, and degeneration. J Neurosci Res 33:505-512.
Koizumi J, Yoshida Y, Nakazawa T (1986) Experimental studies of ischemic brain edema: a new experimental model of cerebral embolism in rats in which recirculation can be introduced in the ischemic area. Jpn J Stroke 8:1-8.
Lee JS, Hong JM, Moon GJ, Lee PH, Ahn YH, Bang OY (2010) A longterm follow-up study of intravenous autologous mesenchymal stem cell transplantation in patients with ischemic stroke. Stem Cells 28:1099-1106.
Li J, Zhu H, Liu Y, Li Q, Lu S, Feng M, Xu Y, Huang L, Ma C, An Y, Zhao RC, Wang R, Qin C (2010) Human mesenchymal stem cell transplantation protects against cerebral ischemic injury and upregulates interleukin-10 expression in Macacafascucularis. Brain Res 1334:65-72.
Li Y, Jiang N, Powers C, Chopp M (1998) Neuronal damage and plasticity identi fi ed by microtubule-associated protein 2, growth-associated protein 43, and cyclin D1 immunoreactivity after focal cerebral ischemia in rats. Stroke 29:1972-1981.
Matesic DF, Lin RCS (1994) Microtubule-associated protein 2 as an early indicator of ischemia-induced neurodegeneration in the gerbil forebrain. J Neurochem 63:1012-1020.
Markgraf CG, Green EJ, Hurwitz BE (1992) Sensorimotor and cognitive consequences of middle cerebral artery occlusion in rats. Brain Res 575:238-246.
Nagasawa H, Kogure K (1989) Correlation between cerebral blood fl ow and histologic changes in a new rat model of middle cerebral artery occlusion. Stroke 8:1037-1040.
Mahmood A, Lu D, Lu M, Chopp M (2003) Treatment of traumatic brain injury in adult rats with intravenous administration of human bone marrow stromal cells. Neurosurgery 53:697-702.
Schallert T, Whishaw IQ (1984) Bilateral cutaneous stimulation of the somatosensory system in hemidecorticate rats. Behav Neurosci 98:518-540.
Shen LH, Li Y, C hen K, Cui Y, Zhang C, Kapke A, Lu M, Savant-Bhonsale S, Chopp M (2007) One-year follow-up after bone marrow stromal cell treatment in middle-aged female rats with stroke. Stroke 38:2150-2156.
Shinozuka K, Dailey T, Tajiri N, Ishikawa H, Kim DW, Pabon M (2013) Stem cells for neurovascular repair in stroke. J Stem Cell Res Ther 4:129-132.
Skene J (1989) Axonal growth-associated proteins. Annu Rev Neurosci 12:127-156.
Stroemer RP, Kent TA, Hulsebosch CE (1993) Acute increase in expression of growth associated protein GAP43 following cortical ischemia in rat. Neurosci Lett 162:51-54.
Stoemer RP, Kent TA, Hulsebosch CE (1995) Neocortical Neural sprouting, synaptogenesis, and behavioral recovery after neocortical infarction in rat. Stroke 26:2135-2144.
Swanson RA, Morton MT, Tsao-Wu G, Savalos RA, Davidson C, Sharp FR (1990) A semiautomated method for measuring brain infarct volume. J Cereb Blood Flow Metab 10:290-293.
Tang Y, Yashuhara T, Hara K, Matsukawa N, Maki M, Xu L, HessDC, Borlongan CV (2007) Transplantation of bone marrow-derived stem cells: a promising therapy for stroke. Cell Transplant 6:159-169.
Woodbury D, Schwarz EJ, Prockop DJ, Black IB (2000) Adult rat and human bone marrow stromal cells differentiate into neurons. J Neurosci Res 61:364-370.
Yam PS, dewar D, McCulloch J (1998) Axonal injury in the rat after a focal ischemic lesion. J Neurotrauma 15:441-450.
Yam PS, Dunn LT, Graham DI, Dewar D, McCulloch J (2000) NMDA receptor blockade fails to alter axonal injury in focal cerebral ischemia. J Cereb Blood Flow Metab 20:772-779.
Zhang J, Li Y, Chen J, Cui Y, Lu M, Elia SB, Mitchell JB, Hammill L, Vanguri P, Chopp M (2005) Human bone marrow stromal cell treatment improves neurological functional recovery in EAE mice. Exp Neurol 195:16-26.
Copyedited by McGowan D, Norman C, Wang J, Yang Y, Li CH, Song LP, Zhao M
10.4103/1673-5374.147930
Xinguang Yu, M.D., Department of Neurosurgery, General Hospital of Chinese PLA, Beijing 100853, China, strongxu2002@sina.com.
http://www.nrronline.org/
Accepted: 2014-11-26
杂志排行

中国神经再生研究(英文版)的其它文章
- Angioplasty and stenting for severe vertebral artery ori fi ce stenosis: effects on cerebellar function remodeling veri fi ed by blood oxygen level-dependent functional magnetic resonance imaging
- A more consistent intraluminal rhesus monkey model of ischemic stroke
- Pathogenesis of glaucoma: how to prevent ganglion cell from axonal destruction?
- Puerarin protects brain tissue against cerebral ischemia/reperfusion injury by inhibiting the in fl ammatory response
- Pretreatment with scutellaria baicalensis stem-leaf total fl avonoid protects against cerebral ischemia/ reperfusion injury in hippocampal neurons
- Overexpression of C-terminal fragment of glutamate receptor 6 prevents neuronal injury in kainate-induced seizure via disassembly of GluR6-PSD95-MLK3 signaling module