秦皇岛入海口邻近海域夏季大型底栖动物群落结构变化及影响因子研究
2014-05-22张建乐牛纪祝石伟杰
李 莉,张建乐,牛纪祝,石伟杰,马 新
(国家海洋局 秦皇岛海洋环境监测中心站,河北 秦皇岛 066002)
0 引言
近年来,随着区域经济快速发展,大量污染物随入海河流排入海中,沿岸河口生态环境承受巨大压力。大型底栖生物作为海洋生态系统的重要组成部分,因其群落组成结构及动态变化,通常对环境状况和变化具有重要的指示作用,已在污染监测和环境质量评价中被加以利用。国内学者已对胶州湾、珠江口、渤海湾、长江口以及泉州湾等近岸海域大型底栖动物群落结构进行了调查研究[1-6]。本文调查区位于大蒲河口、人造河口和洋河口邻近海域,这3个河口是秦皇岛海域的主要排污口。为了解秦皇岛河口近岸海域大型底栖动物群落结构的变化,分别于2009年夏季和2012年夏季对秦皇岛河口近岸海域大型底栖生物群落进行调查分析,旨在探明秦皇岛河口近岸海域生态环境质量变化规律,从而为海洋资源进一步开发和利用提供理论依据。
1 调查研究方法
1.1 采样方法
2009年8月和2012年8月分别对秦皇岛河口近岸海域进行了大型底栖动物调查,共布设15个调查站(图1),每个站位用0.05m2的表层采泥器重复取样2次(以2次成功采样为准)。用孔径为0.5mm的网筛冲洗去泥,所获生物样品用体积分数5%的中性甲醛溶液固定,具体操作按《海洋监测规范》(GB17378.7-2007)第七部分:近海污染生态调查和生物监测处理[7]。

图1 采样站位Fig.1 Sampling stations
1.2 数据处理分析
群落多样性采用香农-威纳多样性指数(Shannon-Wiener index,H′)、Pielou均匀度指数(J′)和丰度指数(D)分析,计算公式为:
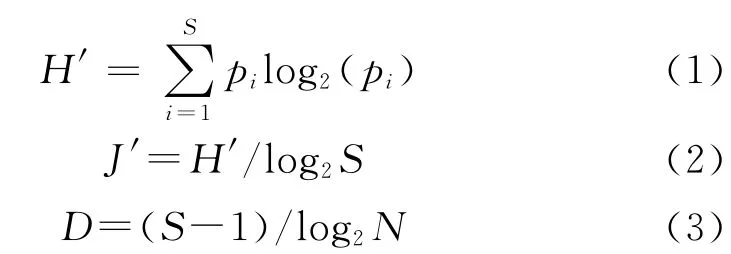
式中:pi为测站第i种的丰度与样品丰度的比值,S为测站中样品的总种类数,N为样品的总丰度。
优势种的确定采用物种的优势度(Y),公式为:

式中:ni为群落中第i种的丰度,fi为该种的出现频率,N为总丰度。当Y>0.02时,该种为优势种[8]。
应用PRIMER 5.0软件进行分析:大型底栖动物群落划分采用Bray-Curtis相似性系数聚类分析(CLUSTER)和多维排序尺度排序(MDS)分析相结合的方法;通过ANOSIM分析检验不同群落间结构差异的显著性;通过SIMPER分析群落中的优势种和造成群落差异的物种;通过BIOENV和BVSTEP分析得出能够影响大型底栖动物群落结构的最佳环境因子组合[9-10]。
应用SPSS 13.0软件对大型底栖动物种类数、丰度和多样性指数与主要环境因子之间的相关性进行双变量相关分析。
1.3 功能群的划分
底栖动物功能群是具有相同生态功能的底栖动物的组合。按照食性类型将近岸海域大型底栖动物划分为以下5类功能群[11-12]:
(1)浮游生物食者(planktophagous group,Pl):依靠各种过滤器官滤取水体中微小的浮游生物,如许多双壳类和甲壳类等。
(2)植食者(phytophagous group,Ph):主要以维管束植物和海藻为饵料,如某些腹足纲、双壳纲和蟹类等。
(3)肉食者(camivorous group,C):捕食小型动物和动物幼体,如某些环节动物和十足类等。
(4)杂食者(omnivorous group,O):依靠皮肤或鳃的表皮,直接吸收溶解在水中的有机物,也可取食植物腐叶和小型双壳类、甲壳类,如某些腹足纲、双壳纲和蟹类等。
(5)碎屑食者(detritivorous group,D):摄食地表的有机碎屑,吞食沉积物,在消化道内摄取其中的有机物质,如某些线虫和双壳类等。
2 结果
2.1 种类组成
2009年夏季共鉴定出大型底栖动物34种,其中多毛类17种,软体类6种,节肢类8种,鱼类1种,纽形类1种,星虫类1种。以优势度Y>0.02界定优势种,2009年夏季优势种为日本美人虾Callianassa japonica和钩虾Gammarus sp.。2012年夏季共鉴定出大型底栖动物27种,其中多毛类12种,软体类4种,节肢类8种,棘皮类2种,纽虫类1种。优势种为豆形短眼蟹Xenophthalmus pinnotheroides和彩虹明樱蛤Moerella iridescens。与2009年相比,2012年优势种种类变化较大,但物种优势度相差不大。从优势种优势度可以看出,2009年和2012年夏季优势物种的主导作用均较明显,主要原因是日本美人虾和豆形短眼蟹的丰度值皆极高,消弱了其他物种在数量上的优势。
2.2 大型底栖动物丰度和生物量
2009年8月各站位大型底栖动物丰度变化范围为30~460ind/m2,平均丰度为186ind/m2。丰度组成以节肢类为主,约占总丰度的68.21%;多毛类丰度次之,约占总丰度的19.29%;其他4类则很少。2012年8月各站位大型底栖动物丰度变化范围为20~520ind/m2,平均丰度为116ind/m2。丰度组成以节肢类为主,约占总丰度的59.77%;多毛类和软体类的丰度次之,均约占总丰度的17.82%;其他2类则很少。大型底栖动物丰度高值区主要集中出现在外侧海域,呈现由近岸向外海递增的趋势(图2)。
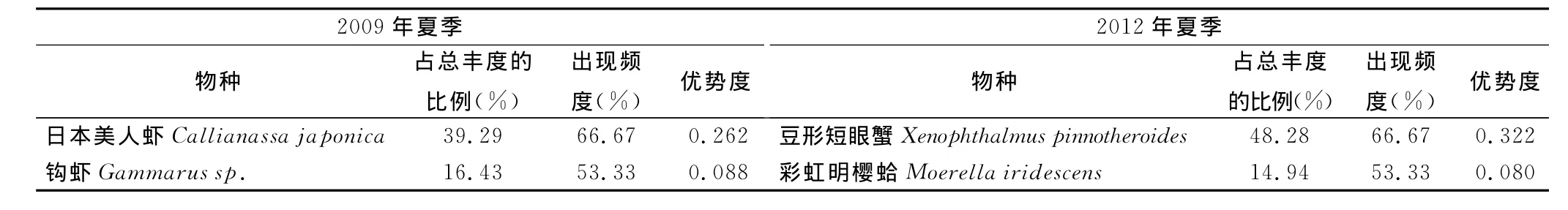
表1 调查海域大型底栖动物优势种及优势度Tab.1 Dominant species and predominance of macrobenthos in the survey area

图2 调查区夏季大型底栖动物丰度的平面分布Fig.2 Horizontal distribution of macrobenthic abundance in survey area in summer
2009年8月各站位大型底栖动物生物量变化范围为0.65~330.43g/m2,平均生物量为60.26g/m2。其中软体类生物量最高,约占总生物量的38.96%;节肢类和多毛类的生物量次之,分别约占总生物量的28.87%和22.04%;其他3类则较少。2012年8月各站位大型底栖动物生物量变化范围为0.44~87.94g/m2,平均值为11.33g/m2。其中棘皮类生物量最高,约占总生物量的49.96%;节肢类次之,约占总生物量的35.28%;其他3类则较少。
2.3 大型底栖动物群落多样性分析
2009年夏季调查海域大型底栖动物Shannon-Wiener多样性指数介于0.337~2.58,平均值为1.91。除人造河口部分站位均匀度指数较小外,其他站位的均匀度指数均介于0.6~1.0之间。Margalef丰度指数介于0.137~0.978,平均值为0.649。Shannon-Wiener多样性指数呈现由近岸向外海递增的趋势,低值区主要集中在人造河口的西南部。
2012年夏季调查海域大型底栖动物Shannon-Wiener多样性指数介于0.00~2.65,平均值为1.69。除人造河口和洋河口个别站位均匀度指数较小外,其他站位的均匀度指数均介于0.7~1.0之间。Margalef丰度指数介于0.00~0.903,平均值为0.518。Shannon-Wiener多样性指数主要集中在0.5~1.5之间,分布不均匀,高值区和低值区呈斑块状零星分布。与2009年相比,2012年夏季大型底栖动物Shannon-Wiener多样性指数和 Margalef丰度指数略有下降,Pielou均匀度指数略有升高。
2.4 大型底栖动物群落结构分析
从图3可以看出,2009年夏季调查海域大型底栖动物可划分为3个群落:群落Ⅰ仅由D3站组成,优势种为菲律宾蛤仔Venerupis Philippinaram和西方似蛰虫Amaeana occidentalis;群落Ⅱ由R2、D2和Y1站组成,优势种为豆形短眼蟹和独指虫Aricidea fragilis;群落Ⅲ由 R1,R3,R4,R5,D1,D4,D5,Y2,Y3,Y4和Y5站组成,优势种为日本美人虾和钩虾。通过对3个聚类组进行相似性分析(ANOSIM),表明不同群落类型之间大型底栖动物组成呈显著性差异(R=0.818,P =0.1%<0.05)。由SIMPER分析进一步了解群落间的差异性大小及形成差异的原因。结果显示:群落Ⅰ和群落Ⅱ之间差异最大,不相似性达到100%,两个群落的物种完全不同;其次群落Ⅰ和群落Ⅲ间不相似性系数为96.78,主要贡献者为菲律宾蛤仔、日本美人虾、钩虾和西方似蛰虫。

图3 2009年夏季大型底栖动物群落结构聚类图(a)和多维尺度分析图(b)Fig.3 Cluster dendroguam(a)and multi-dimentional scaling(MDS)analysis(b)of macrobenthos community in summer 2009
从图4可以看出,2012年夏季调查海域大型底栖动物可划分为4个群落:群落Ⅰ由D1和D2站组成,优势种为多齿全刺沙蚕Nectoneanthes multignatha;群落Ⅱ由R1,D3和Y3站组成,优势种为豆形短眼蟹和长吻沙蚕Glycera chirori;群落Ⅲ由Y4和Y5站组成,优势种为豆形短眼蟹和轮双眼钩虾Ampelisca cyclops;群落Ⅳ由R2,R3,R4,R5,Y1,Y2,D4和D5站组成,优势种为豆形短眼蟹和彩虹明樱蛤。通过对4个聚类组进行相似性分析(ANOSIM),表明不同群落类型之间的大型底栖动物组成呈显著性差异(R=0.806,P =0.1%<0.05)。SIMPER分析显示,群落Ⅰ和群落Ⅱ之间差异最大,不相似性达到100%,两个群落的物种完全不同;其次群落Ⅰ和群落Ⅳ间不相似系数为98.16,主要贡献者是豆形短眼蟹、彩虹明樱蛤、多齿全刺沙蚕及小头虫。由此可以看出,2009年夏季和2012年夏季大型底栖动物群落不相似性主要贡献者相差较大。
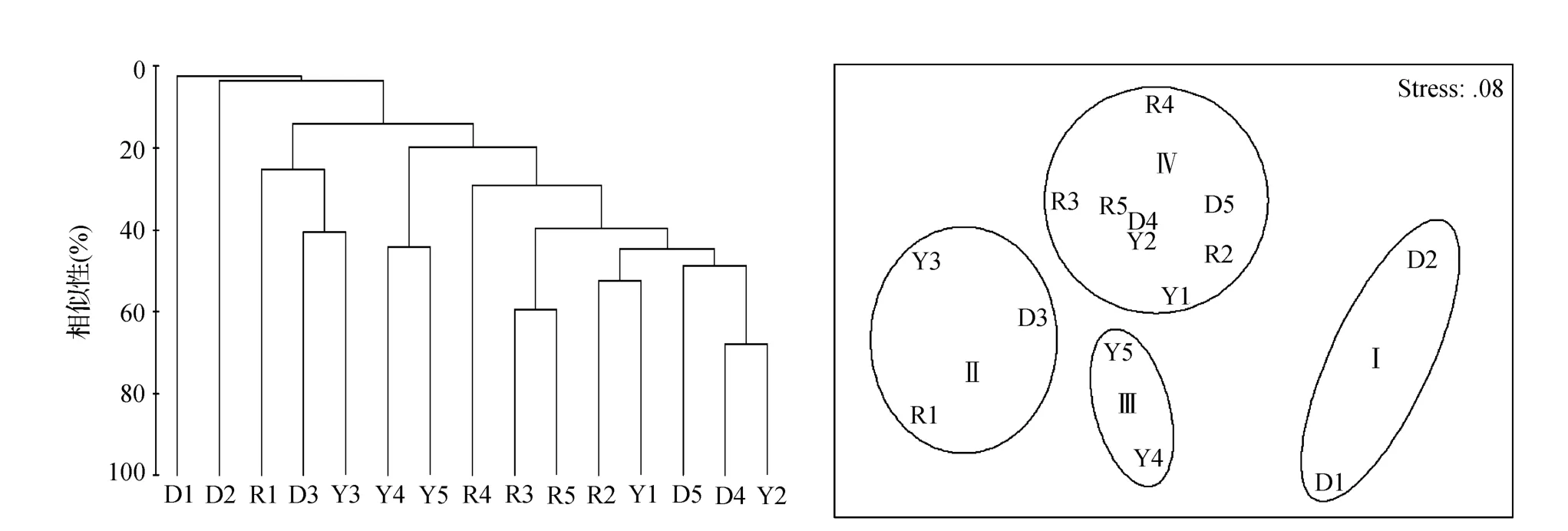
图4 2012年夏季大型底栖动物群落结构聚类图(a)和多维尺度分析图(b)Fig.4 Cluster dendroguam(a)and multi-dimentional scaling(MDS)analysis(b)of macrobenthos community in summer 2012
2.5 大型底栖动物功能摄食类群
2009年夏季调查海域大型底栖动物功能群种类组成以肉食者和碎屑食者为主,功能群丰度百分比高低依次为D>O>C>Pl。群落Ⅰ功能摄食类群的密度相对丰度最大为浮游生物食者,其密度相对丰度为60%;群落Ⅱ以杂食者占主要优势,其密度相对丰度为62.96%;群落Ⅲ以碎屑食者为主,其密度相对丰度为54.20%,肉食者和杂食者次之,密度相对丰度分别为21.01%和20.59%(图5a)。
2012年夏季调查海域大型底栖动物功能群种类组成以肉食者和碎屑食者为主,功能群丰度百分比高低依次为O>Pl>D>C。群落Ⅰ和群落Ⅱ功能摄食类群的密度相对丰度均以肉食者为主,其密度相对丰度分别为44.44%和50.0%;而群落Ⅲ和群落Ⅳ均以杂食者为主,其密度相对丰度分别为74.07%和58.06%(图5b)。

图5 调查海域大型底栖动物功能摄食类群密度相对丰度Fig.5 Relative density abundance of functional feeding groups for macrobenthos in the survey sea
2.6 大型底栖动物与环境因子相关性分析
为了解不同环境因子对大型底栖动物群落结构的影响状况,对调查海域大型底栖动物群落结构同水深、盐度、COD、无机氮、无机磷、沉积物硫化物、有机碳和石油类分别进行Spearman相关系数计算、BIOENV和BVSTEP分析。

表2 调查海域大型底栖动物与环境因子相关性分析Tab.2 Spearman correlation coefficient between macrobenthos and environmental factors in the survey area
从表2可以看出,2009年夏季大型底栖动物种类数与水深呈极显著正相关,与无机磷呈显著负相关;丰度与水深呈极显著正相关,与COD呈极显著负相关,与沉积物石油类呈显著负相关。2012年夏季大型底栖动物种类数、丰度和生物量均与水深呈极显著正相关;种类数与沉积物硫化物呈显著负相关。
应用PRIMER 5.0软件进行BIOENV和BVSTEP分析显示,影响2009年夏季大型底栖动物群落结构的主要环境因子是水深和无机磷含量组合(r=0.461);影响2012年夏季大型底栖动物群落结构的主要环境因子是水深、盐度和沉积物有机碳含量组合(r=0.260)。
3 讨论
与2009年8月相比,2012年8月大型底栖动物种类数有所减少,种类组成均以多毛类和节肢类为主。就适盐性而言,两次调查主要包括近岸广盐种和河口广盐种,与笔者对滦河口-北戴河海域大型底栖动物的研究结果一致[13]。两次调查秦皇岛近岸海域大型底栖动物物种数和丰度变化趋势一致,皆呈现由近岸向外海递增的趋势。
2009年夏季和2012年夏季调查海区大型底栖动物的总生物量呈现明显变化。以2009年为参考点,2012年夏季大型底栖动物的总生物量下降了81.21%。从4大类底栖动物对总生物量的贡献来看,两次调查的底栖生物群落结构发生了显著改变。2009年夏季调查中软体动物的生物量为352.19 g/m2,占总生物量的38.96%,生物量贡献率最大,其次是节肢动物和多毛类,在调查中未发现棘皮动物。2012年夏季调查中棘皮动物的生物量高达84.87 g/m2,为总生物量的最大贡献者,节肢动物生物量次之,其他3类生物量较少,均约占总生物量的7%。总体而言,河口近岸海域大型底栖动物的总生物量组成变化明显,各生态类群的优势地位出现了明显更替,个体较大、生长周期较长的棘皮动物逐渐取代个体较小、生长周期较短的多毛类,成为秦皇岛入海口邻近海域主要优势种群,这是种群繁殖策略上的一种改变,以适应秦皇岛入海口邻近海域自然环境。
通过BIOENV和BVSTEP分析发现,水深、无机磷、盐度和沉积物有机碳是影响调查海区大型底栖动物群落结构的主要环境因子。两次调查中影响大型底栖动物结构的主要环境因子存在明显差异,除水深外其他主要环境因子皆不同。通常盐度是影响河口区大型底栖生物分布的主要因素[14]。然而2009年夏季调查研究表明盐度对大型底栖动物群落结构影响不大,这可能由于2009年夏季盐度变化相对较小,大型底栖动物栖息在海底,盐度的轻微波动对其群落结构影响较小。无机磷和沉积物有机碳成为本调查海区的主要环境因子,可能与两次调查中大型底栖动物的食物利用方式不同有关。无机磷作为限制性营养盐,影响浮游植物生长,进而影响以浮游植物为食的大型底栖生物,而沉积物有机碳主要影响以有机碳为主要食物来源的大型底栖生物。然而,海洋环境的多变性和复杂性决定了影响大型底栖动物群落结构的因素也相当复杂。除本研究中的环境因子,影响大型底栖动物群落结构的环境因子还有很多,如底层 DO[15]、总氮[16]、总磷[17]、沉积物中重金属[18]和底质类型[19]等,因此要更好地了解影响大型底栖动物群落长期变化的关键因素,还需要长期的历史数据积累和进一步深入研究和分析。
4 结论
(1)2009年和2012年夏季调查海区大型底栖动物种类组成皆以多毛类和节肢动物为主,物种数和丰度皆呈现由近岸向外海递增的趋势。与2009年相比,2012年大型底栖动物种类数有所减少,优势种种类变化较大,但物种优势度变化不明显。
(2)2009年和2012年夏季调查海域大型底栖动物的总生物量组成变化明显,各生态类群的优势地位出现了明显更替。
(3)通过聚类分析,2009年夏季大型底栖动物可划分为3个群落,2012年夏季大型底栖动物可划分为4个群落。两次调查中不同群落类型之间大型底栖动物组成皆呈显著性差异,群落不相似性主要贡献者差异性较大。
(4)根据BIOENV和BVSTEP分析结果表明,2009年夏季影响大型底栖动物群落结构的主要环境因子是水深和无机磷含量;2012年夏季影响大型底栖动物群落结构的主要环境因子是水深、盐度和沉积物有机碳含量。
(References):
[1]BI Hong-sheng,SUN Song,SUN Dao-yuan.Changes of macrobenthic communities in Jiaozhou bay[J].Oceanologia et Limnologia Sinica,2001,32(2):132-137.
毕洪生,孙松,孙道元.胶州湾大型底栖生物群落的变化[J].海洋与湖沼,2001,32(2):132-137.
[2]PENG Song-yao,LAI Zi-ni,JIANG Wan-xiang,et al.Study community structure of macrozoobenthos and impact factors in Pearl River Estuary[J].Acta Hydrobiology Sinica,2010,34(6):1 179-1 189.
彭松耀,赖子尼,蒋万祥,等.珠江口大型底栖动物的群落结构及影响因子研究[J].水生生物学报,2010,34(6):1 179-1 189.
[3]ZHANG Jing-huai,GAO Yang,FANG Hong-da,et al.Ecological characteristics of macrobenthic communities in Pearl River Estuary[J].Acta Ecologica Sinica,2009,29(6):2 989-2 999.
张敬怀,高阳,方宏达,等.珠江口大型底栖生物群落生态特征[J].生态学报,2009,29(6):2 989-2 999.
[4]CAI Wen-qian,LIU Lu-san,QIAO Fei,et al.Study on the changes of macrobenthos communities and their causes in Bohai Bay[J].Environmental Science,2012,33(9):3 104-3 109.
蔡文倩,刘录三,乔飞,等.渤海湾大型底栖生物群落结构变化及原因探讨[J].环境科学,2012,33(9):3 104-3 109.
[5]SUN Ya-wei,CAO Lian,QIN Yu-tao,et al.Analysis of macrobenthos community structure in the adjacent sea area of Changjiang River Estuary[J].Marine Science Bulletin,2007,26(2):66-70.
孙亚伟,曹恋,秦玉涛,等.长江口邻近海域大型底栖生物群落结构分析[J].海洋通报,2007,26(2):66-70.
[6]LI Rong-guan,WANG Jian-jun,ZHENG Cheng-xing,et al.The ecology of macrobenthos community in Quanzhou Bay,Fujian Province[J].Acta Ecologica Sinica,2006,26(11):3 562-3 571.
李荣冠,王建军,郑成兴,等.泉州湾大型底栖生物群落生态[J].生态学报,2006,26(11):3 562-3 571.
[7]GB17378.7—2007The specification for marine monitoring—Part 7:Ecological survey for offshore pollution and biological monitoring[S].2007.
GB17378.7—2007海洋监测规范 第七部分:近海污染生态调查和生物监测处理[S].2007.
[8]CLARKE K R,WARWICK R M.Change in marine communities:An approach to statistic analysis and interpretation [M].Plymouth:Plymouth Marine Laboratory,1994.
[9]CLARK K R,GORLEY R N.PRIMER v5:Use Manual/Tutorial[M].Plymouth:PRIMER-E Ltd.2001.
[10]WU Rong-jun,LI Rui-xiang,ZHU Ming-yuan,et al.Multivariate analysis with primer on marine phytoplankton community structure in mesocosm system[J].Oceanologia et Limnologia Sinica,2006,37(4):316-321.
吴荣军,李瑞香,朱明远,等.应用PRIMER软件进行浮游植物群落结构的多元统计分析[J].海洋与湖沼,2006,37(4):316-321.
[11]BOAVENTURA D,FONSECA L C,TELES-FERREIRA C.Trophic structure of macrobenthic communities on the Portuguese coast:A review of lagoobal,estuarine and rocky littoral habitats[J].Acta Oecologica,1999,20(4):407-415.
[12]ZHU Xiao-jun,LU Jian-jian.Functional groups of zoobenthos in the intertidal zone of Jiuduansha,the Yangtze River Estuary[J].Zoological Rsearch,2003,24(5):355-361.
朱晓君,陆健健.长江口九段沙潮间带底栖动物的功能群[J].动物学研究,2003,24(5):355-361.
[13]LI Li,CHEN Wu-jun,ZHANG Zhen.Ecological characteristics of macrobenthic communities in Luanhe river estuary-Beidaihe[J].Journal of Anhui Agricultural Science,2012,40(26):13 049-13 054.
李莉,陈武军,张臻.滦河口-北戴河海域大型底栖生物群落特征研究[J].安徽农业科学,2012,40(26):13 049-13 054.
[14]HOLLAND A F,SHAUGHNESSY A T,HIEGEL M H.Longterm variation in mesohaline Chesapeake Bay macrobenthos:Spatial and temporal patterns[J].Estuaries,1987,10(3):227-245.
[15]SHOU Lu,ZENG Jiang-ning,LIAO Yi-bo,et al.Seasonal dis-tribution of macrozoobenthos and its relations to environmental factors in Oujiang River estuary sea area[J].Chinese Journal of Applied Ecology,2009,20(8):1 958-1 964.
寿鹿,曾江宁,廖一波,等.瓯江口海域大型底栖动物分布及其与环境的关系[J].应用生态学报,2009,20(8):1 958-1 964.
[16]WANG Zong-xing,WEI Qin-sheng,LIU Jun,et al.Canonical correspondence analysis on macrobenthos community in summer and its environment in the offshore water of Rushan Bay[J].Chin J Appl Environ Biol,2012,18(4):599-604.
王宗兴,韦钦胜,刘军,等.乳山湾外海夏季大型底栖动物分布与环境因子的典范对应分析[J].应用与环境生物学报,2012,18(4):599-604.
[17]LIAO Yi-bo,SHOU Lu,ZENG Jiang-ning,et al.Spatiotemporal distribution of macrobenthic communities and its relationships with environmental factors in Sanmen Bay[J].Chinese Journal of Applied Ecology,2011,22(9):2 424-2 430.
廖一波,寿鹿,曾江宁,等.三门湾大型底栖动物时空分布及其与环境因子的关系[J].应用生态学报,2011,22(9):2 424-2 430.
[18]LI Xin-zheng,LI Bao-quan,WANG Hong-fa,et al.Community structure of macrobenthos in coastal water off Rushan,southern Shandong Peninsula,and the relationships with environmental factors[J].Acta Oceanol Sin,2009,28(5):81-93.
[19]GANESH T,RAMAN A V.Macrobenthic community structure of the northeast Indian shelf,Bay of Bengal[J].Marine Ecology Progress Series,2007,341:59-73.
