不同林龄南方红豆杉人工林细根结构及C和N含量比较与相关性分析
2014-05-11刘艳艳1b林勇明1b陈建忠吴承祯1b
刘艳艳,林 晗,1b,巢 林,林勇明,1b,陈建忠,吴承祯,1b,2,①
(1.福建农林大学:a.林学院,b.福建省高校森林生态系统过程与经营重点实验室,福建福州350002; 2.武夷学院生态与环境学院,福建武夷山354300;3.建阳市林业局,福建建阳354200)
不同林龄南方红豆杉人工林细根结构及C和N含量比较与相关性分析
刘艳艳1a,林 晗1a,1b,巢 林1a,林勇明1a,1b,陈建忠3,吴承祯1a,1b,2,①
(1.福建农林大学:a.林学院,b.福建省高校森林生态系统过程与经营重点实验室,福建福州350002; 2.武夷学院生态与环境学院,福建武夷山354300;3.建阳市林业局,福建建阳354200)
对不同林龄南方红豆杉〔Taxus chinensis var.mairei(Lemee et Lévl.)Cheng et L.K.Fu〕人工林不同等级细根的结构参数及C和N含量进行了比较,并分析了细根结构参数与C和N含量的相关性。结果显示:不同林龄南方红豆杉Ⅰ~Ⅴ级细根的根直径、比根长(SRL)和比表面积(SSA)均有差异,6、7和8 a植株细根的平均直径分别为1.073、1.815和1.734 mm,平均SRL分别为14.09、12.88和14.12 m·g-1,平均SSA分别为54.93、45.85和50.72 cm2·g-1;随根序等级提高,同龄植株细根的SRL和SSA均依次降低、根直径则逐渐增大,且总体差异显著(P<0.05);总体上,除Ⅰ级细根外,7 a植株各级细根的SRL和SSA均最小;6 a植株各级细根的直径均最小。各林龄植株细根的根尖密度和分叉数无显著差异(P>0.05),但均以6 a植株最大。细根干质量以6 a植株最低、7 a植株最高,且差异显著。随根序等级提高,细根C含量逐渐增加、N含量则逐渐降低,同龄植株Ⅰ~Ⅴ级细根的C和N含量总体上差异显著;各级细根中,7 a植株的C含量均最高、8 a植株的N含量均最低;6、7和8 a植株细根的平均C含量分别为454.41、501.90和441.55 mg·g-1,平均N含量分别为12.63、11.99和5.88mg·g-1。各林龄植株细根的C含量与细根直径和干质量均正相关,与SRL、SSA和根尖密度均负相关;N含量与细根直径均负相关,与SRL、SSA和根尖密度均正相关;此外,N含量与干质量以及C和N含量与分叉数的相关性均无规律性。研究结果表明:树龄对南方红豆杉细根直径影响较大,而不同序级细根的结构差异更明显。
南方红豆杉人工林;细根;结构;C和N含量;林龄;相关性
细根为植物的功能性根,其分支系统复杂[1],对林木的生理功能有重要作用[2];细根不但是森林生态系统碳和养分循环的重要组成部分[3-4],也是供应水分和养分的重要器官[5]。细根的数量、分布、形态和结构与其功能密切相关,这些特征除受其自身遗传特性的影响外,对土壤资源状况及环境变化也较为敏感[6]。此外,细根不是一个均质的周转很快的系统,而是由形态、寿命及生理上差异巨大的不同部分组成的高度异质的混合体[7],对细根形态异质性的研究是认识细根动态变化、了解细根功能及准确估计细根寿命的前提[8-9]。故对同一树种不同林龄细根的结构、C和N含量及其相互关系的研究对进一步揭示细根结构对植株C和N分布的影响特征有重要作用。
目前大部分学者将直径小于2 mm作为细根的划分标准,但Pregitzer等[10]认为这类根的形态和功能差异很大,而按照根系发育顺序并利用序级划分细根,对其结构与功能关系的预测十分有利[11]。此外,与地上部分的相关研究相比,细根的研究缺乏深度和广度,特别是对细根结构与功能方面的研究较少[12],研究内容集中于其序级和直径等结构参数对C、N含量的影响,且获得的研究结果相似,但对细根的其他结构参数与其C、N含量间的关系尚不明了。
目前,有关南方红豆杉〔Taxus chinensis var. mairei(Lemee et Lévl.)Cheng et L.K.Fu〕生态特征的研究较多,主要涉及种群生命表[13]、种内种间竞争[14]和点格局[15]等方面,而对其根系特征及根系在养分循环中的作用等方面的相关研究有所欠缺。鉴于此,作者以不同林龄(6、7和8 a)南方红豆杉人工林为研究对象,测定不同等级细根的直径、比根长、比表面积、根尖密度和分叉数及干质量等结构和形态参数,并测定细根的C和N含量,明确其细根时空分布特征及其对C、N吸收与释放的影响效应,为南方红豆杉人工林合理经营提供参考依据。
1 研究地概况和研究方法
1.1 研究地概况
研究地位于福建省建阳市溪东国有林场内,地处武夷山南麓;地理坐标为北纬27°20′、东经118°19′,平均海拔352 m,面积3 383 km2。属中亚热带季风性气候;昼夜温差大,年均温18℃;雨季集中,年均降雨量1 697 mm;年均日照时数1 802 h,无霜期282 d。植被属中亚热带常绿阔叶林,森林覆盖率达72.5%。土壤为红壤和黄壤,土层厚度90~110 cm,有机质含量3%~5%,pH 5.0~pH 6.5,含沙量低于10%。
于2005年、2006年和2007年将南方红豆杉套种于1989年种植的杉木〔Cunninghamia lanceolata (Lamb.)Hook.〕人工林中,形成杉木-南方红豆杉混交林,南方红豆杉初始种植密度均为10 000 hm-2。选取伴生树种、林分结构、立地条件和森林经营管理措施基本一致但南方红豆杉株龄分别为6、7和8 a的林分设置样地,各林分的基本概况见表1。每一林龄的林分各设置3块面积为20 m×20 m的样地,每块样地各选取3株大小相近的样株。
1.2 研究方法
1.2.1 细根采集和处理方法 于2013年7月上旬参照Guo等[9]的完整土块法取样。在每株南方红豆杉样株基部0.5~1.0 m范围内分别挖取4个10 cm× 10 cm×20 cm的土块,将土块内完整根系全部取出,清除根表面土壤后装入自封袋,置于2℃~3℃冰箱内冷藏。根据Pregitzer等[10]的分级方法对采集的完整根系进行分级,最远端的根尖定为Ⅰ级根,2个Ⅰ级根交汇处为Ⅱ级根,依次类推至Ⅴ级根。分别将分级后的各级细根装入盛有去离子水的玻璃皿内,置于冰箱中冷藏,供结构及形态指标测定。
1.2.2 指标测定方法 用Epson Expression 10000 XL型数字化扫描仪(Epson America,Inc.)分别对各级细根进行扫描,3次重复;然后用Win RHIZO(Pro 2009c)根系图像分析软件分析细根图像,直接获得各级细根的直径、根长、表面积、根尖数及分叉数。扫描完成后先分别称量各级细根的鲜质量,然后将所有样品分别置于65℃烘箱中处理48 h,称量各级细根的干质量。根据以下公式计算细根的比根长(SRL, specific root length)、比表面积(SSA,specific surface area)、根尖密度和分叉数:SRL=总根长/干质量; SSA=根表面积/干质量;根尖密度=各林龄细根根尖数/根长;分叉数=各林龄细根分叉数/根长。
将上述烘干后的各级细根分别放入玛瑙研钵中研磨成粉并过2 mm筛,装入自封袋中备用。分别精确称量约1 mg(精度0.01)细根粉末,用锡纸小杯包裹;用Euro Vector EA 3000型元素分析仪(Euro Vector S.P.A,Italy)分析各级细根的C和N含量,重复3次。用Callidus Software Interface(Version 5.1)软件直接读取数据。
1.3 数据计算和统计分析
用EXCEL 2007数据处理软件对C和N含量测定数据进行单位换算。利用SPSS 13.0统计分析软件、采用单因素和双因素方差分析法进行差异显著性分析;采用相关性分析和回归分析法分析不同林龄细根的结构和形态指标与C、N含量的相关性。
2 结果和分析
2.1 细根形态和结构指标的比较
不同林龄南方红豆杉不同等级细根的直径、比表面积(SSA)和比根长(SRL)以及细根的根尖密度、分叉数和干质量的测定结果见表2。
由表2可见:不同林龄南方红豆杉Ⅰ~Ⅴ级细根的SRL和SSA均有一定差异。Ⅰ级细根的SRL以7 a植株最大、8 a植株最小,且二者间差异显著(P<0.05);而Ⅱ~Ⅴ级细根的SRL均以8 a植株最大、7 a植株最小;3个林龄Ⅱ级细根的SRL差异显著,7 a植株Ⅲ~Ⅴ级细根的SRL显著小于6和8 a植株,但后二者间差异不显著。Ⅰ级细根的SSA以7 a植株最大、8 a植株最小;Ⅱ级细根的SSA以8 a植株最大、7 a植株最小,但三者间均无显著差异;Ⅲ级细根的SSA以6 a植株最大、7 a植株最小,Ⅳ和Ⅴ级细根的SSA以8 a植株最大、7 a植株最小,但6和8 a植株间Ⅲ~Ⅴ级细根的SSA差异不显著。
6、7和8 a南方红豆杉细根的平均SRL分别为14.09、12.88和14.12 m·g-1,平均SSA分别为54.93、45.85和50.72 cm2·g-1;同一林龄植株各级细根的SRL和SSA的变化规律相似,均随根序等级升高依次降低,且同一林龄各级细根的SRL和SSA总体上差异显著。总体上看,除Ⅰ级细根外,7 a南方红豆杉的不同等级细根的SRL和SSA均小于6和8 a南方红豆杉。表明不同等级细根的SRL和SSA差异比不同林龄间更为明显。
不同林龄南方红豆杉的细根直径差异明显,细根平均直径以7 a植株最大(1.815 mm)、8 a植株次之(1.734 mm)、6 a植株最小(1.073 mm)。其中,6 a植株的各级细根直径均最小且总体上与另2个林龄植株差异显著;Ⅰ和Ⅱ级细根直径均随林龄增大逐渐增加且均以8 a植株最大,Ⅲ~Ⅴ级细根直径则以7 a植株最大,但7和8 a植株Ⅰ~Ⅴ级细根直径均无显著差异。同一林龄Ⅰ~Ⅴ级细根直径均随根序等级提高逐渐增大,各等级细根直径总体上差异显著。
根尖密度和分叉值越大,表明根系吸收养分或水分的能力越强。不同林龄南方红豆杉细根的根尖密度和分叉数无显著差异,但均以6 a植株最大,表明其根系的吸收能力大于7和8 a植株。不同林龄植株的细根干质量有一定差异,以6 a植株最低,7 a植株最高,且后者是前者的2倍以上,差异显著;8 a植株细根干质量则与6和7 a植株无显著差异。
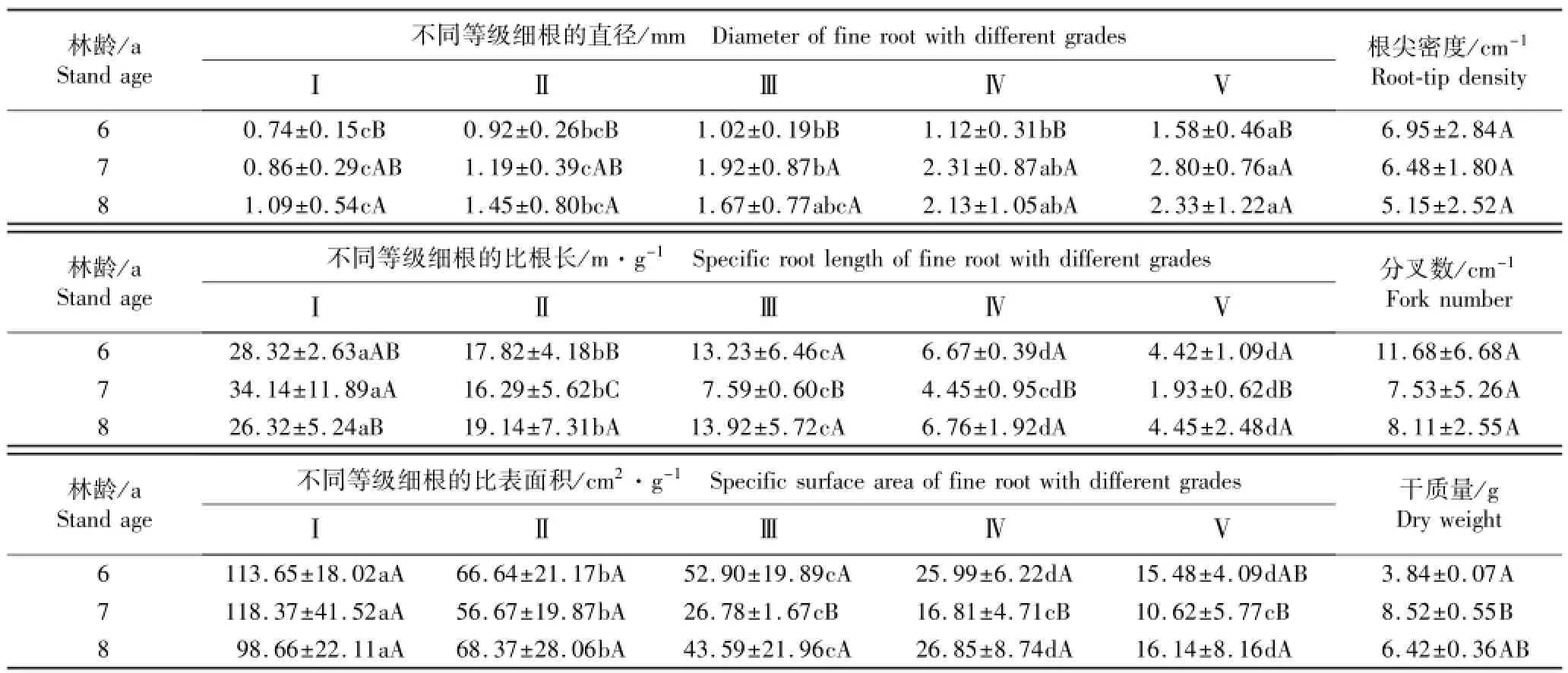
表2 不同林龄南方红豆杉人工林细根结构和形态指标的比较(¯X±SD)1)Table 2 Com parison on fine root structure and morphological indexes of Taxus chinensis var.mairei(Lem ee et Lévl.)Cheng et L.K.Fu plantation w ith different stand ages(¯X±SD)1)
2.2 细根中C和N含量的比较
不同林龄南方红豆杉各级细根中C和N含量的测定结果见表3。结果表明:随根序等级增大,不同林龄南方红豆杉细根中C含量均呈逐渐上升的趋势,而N含量则呈逐渐降低的趋势;林龄、根序以及林龄和根序的交互作用对细根中C和N含量的影响均达到极显著(P<0.01)或显著水平(P<0.05)。
6、7和8 a南方红豆杉细根的平均C含量分别为454.41、501.90和441.55 mg·g-1。各级细根的C含量均以7 a植株最高,且与6和8 a植株差异显著;Ⅰ和Ⅱ级细根的C含量以6 a植株最低,Ⅲ~Ⅴ级细根的C含量以8 a植株最低;但6和8 a植株Ⅰ~Ⅲ级细根C含量无显著差异,二者的Ⅳ和Ⅴ级细根C含量则差异显著。此外,同一林龄Ⅰ~Ⅴ级细根的C含量总体上差异显著。
各级细根的N含量均以8 a植株最低,且与6和7 a植株差异显著;Ⅰ和Ⅱ级细根的N含量以7 a植株最高、6 a植株次之,二者间无显著差异;Ⅲ~Ⅴ级细根的N含量以6 a植株最高、7 a植株次之,二者间差异显著;6、7和8 a植株细根的平均N含量分别为12.63、11.99和5.88 mg·g-1。此外,同林龄Ⅰ~Ⅴ级细根的N含量总体上差异显著。
2.3 细根形态和结构指标与C和N含量的相关性
不同林龄南方红豆杉细根的形态和结构参数与其C和N含量的相关系数见表4。
不同林龄植株细根的C含量与其直径和干质量均呈正相关,与SRL、SSA和根尖密度均呈负相关。其中,6 a植株细根的C含量与SRL显著相关(P<0.05),与SSA极显著相关(P<0.01);7 a植株细根的C含量与根直径、SRL、SSA和干质量均显著相关;8 a植株细根的C含量与SRL、根尖密度和干质量均显著相关,与SSA极显著相关;各龄级植株细根的C含量与其余指标的相关性均未达到显著水平(P>0.05)。
3个林龄南方红豆杉细根的N含量与细根直径均呈负相关,与SRL、SSA和根尖密度均呈正相关,6 a植株细根的N含量与干质量呈正相关,7和8 a植株细根的N含量与干质量则呈负相关。其中,6 a植株细根的N含量与根尖密度显著相关,与根直径极显著相关;7 a植株细根的N含量与SRL和SSA显著相关,与根直径极显著相关;8 a植株细根的N含量与SRL、SSA和干质量显著相关,与根尖密度极显著相关;各龄级植株细根N含量与其余指标的相关性均没有达到显著水平。
此外,不同林龄南方红豆杉细根的C和N含量与分叉数呈正相关或负相关,但相关性均不显著。表明细根的分叉数对其营养成分积累无明显影响。
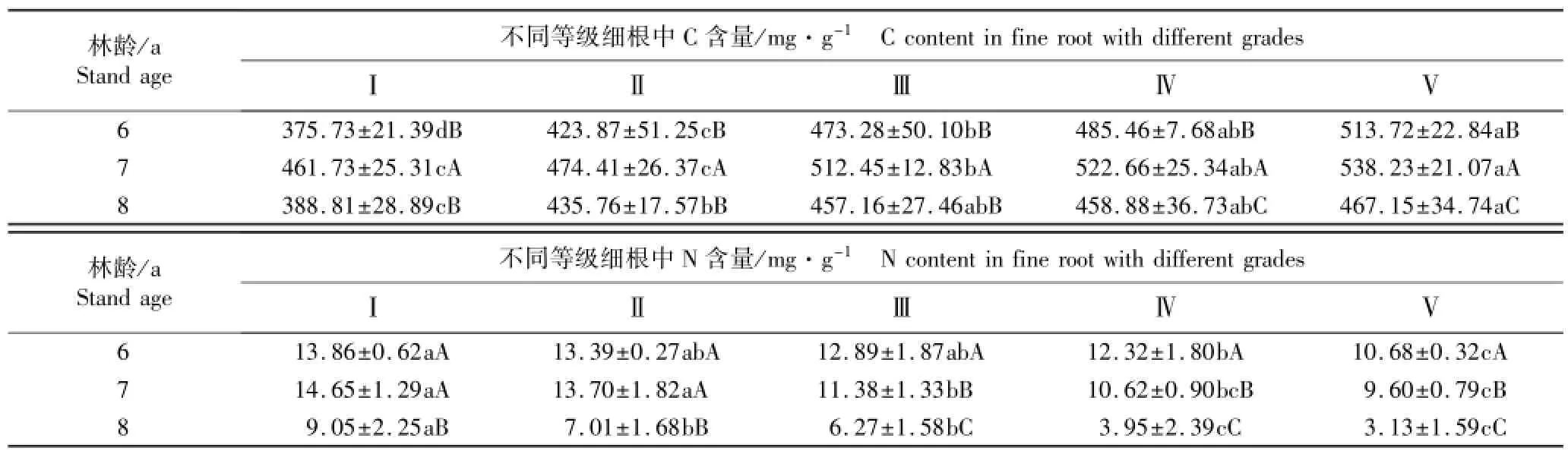
表3 不同林龄南方红豆杉人工林细根中C和N含量的比较(¯X±SD)1)Table 3 Comparison on C and N contents in fine root of Taxus chinensis var.mairei(Lemee et Lévl.)Cheng et L.K.Fu plantation w ith different stand ages(¯X±SD)1)
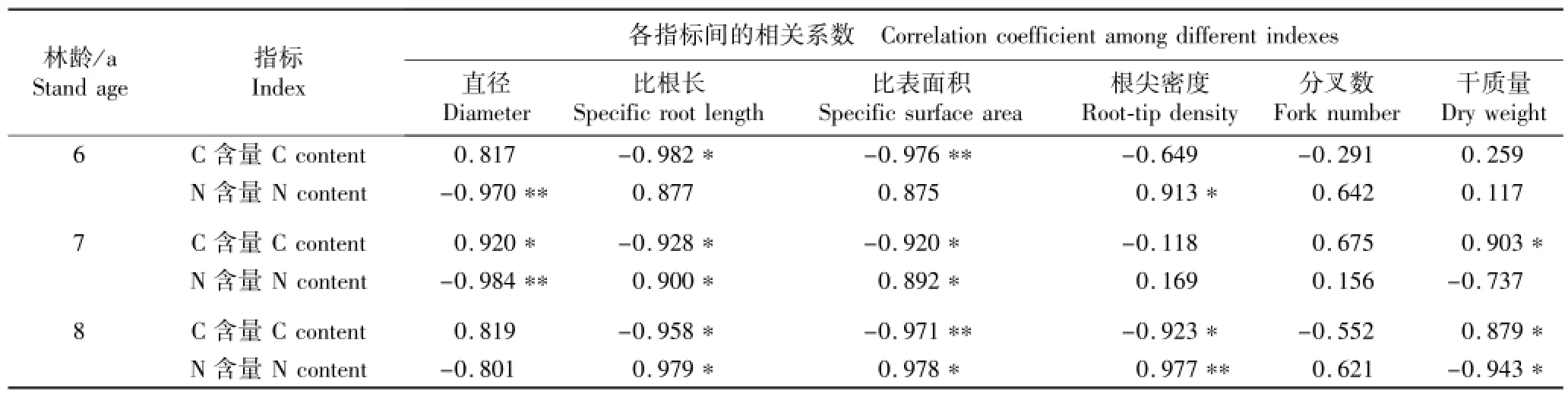
表4 不同林龄南方红豆杉人工林细根结构和形态指标与C和N含量的相关系数1)Table 4 Correlation coefficient of structure and morphological indexesw ith C and N contents of fine root of Taxus chinensis var.mairei(Lemee et Lév l.)Cheng et L.K.Fu p lantation with different stand ages1)
3 讨论和结论
根系表面积是研究水分吸收或养分吸收的重要参数之一[16],而细根比表面积则表示相同单位的面积投入到细根生物量用于吸收养分的效率,其值越大则表明投入相同成本养分吸收效率就越高[17];比根长则是评估植物吸收水分和养分的重要指标[18]。他人的研究结果[19-22]表明:随根序等级增加,细根直径增加,而比根长和比表面积则降低。本研究中,3个林龄南方红豆杉细根均表现出“Ⅰ级根比根长和比表面积最高、直径最小,Ⅴ级根比根长和比表面积最低、直径最大”的现象,尤其7和8 a植株细根的直径可达2.33 mm以上,与前人得出的规律基本一致。但是, Pregitzer等[10]认为:低级根直径都小于0.3 mm、根长都小于2 mm;而本研究中,3个林龄植株的低等级细根的最小直径为0.74 mm、根长均大于2 mm,均高于前述的标准。这一现象与南方红豆杉自身的生理特征密切相关,植株可能通过根系连接长度的增加提高其对营养物质的获取能力[23]。此外,在不同的气候区域(温带、亚热带或热带)间细根直径均有较大差异,而不同树种的细根在结构(根序)和功能上也存在很大差异,如果仅依据“直径小于2mm或小于1mm”的标准来定义细根则具有一定的局限性[20]。
研究结果表明:南方红豆杉Ⅰ和Ⅱ级细根的直径随林龄增加显著增加;与6 a植株相比,7 a植株的Ⅴ级细根的直径明显增大,而其Ⅰ级细根的比根长和比表面积明显提高。而李瑞霞等[24]的研究结果表明:与40 a侧柏〔Platycladus orientalis(Linn.)Franco〕人工林相比,48 a侧柏的Ⅴ级细根直径显著降低、Ⅴ级细根根长和Ⅰ级细根的比根长显著增加;55 a侧柏Ⅳ级细根的直径和根长以及Ⅰ级细根的比根长显著增加。这种差异性和相似性除与树种自身遗传差异有关外,还与林龄长短、林龄间隔以及林分密度有关,且林龄间隔能直接影响不同树种细根的周转状况。此外,树龄对细根比表面积和比根长的影响不显著,而对细根直径有极显著影响;而各级细根间的比根长、比表面积和直径差异均达到极显著水平,表明同一树种不同序级间细根的结构差异更明显。
南方红豆杉细根的干质量以7 a植株最大、8 a植株次之、6 a植株最小,且6与7 a植株细根的干质量差异达显著水平。这与刘波等[25]对亚热带常绿阔叶林和Peichl等[26]对美国白松(Pinus strobus Linn.)的相关研究结果相似。而林希昊等[27]对不同林龄橡胶〔Hevea brasiliensis(Willd.ex A.Juss.)Müll.Arg.〕林细根生物量的研究以及马炜等[28]对不同林龄长白山落叶松(Larix olgensis A.Henry)细根生物量的研究均得出“根系生物量随林龄增大而增加”的结果,可能与样地的受干扰程度、林分密度以及树龄间隔等差异有关。林木细根生物量还受产地的气候、土壤类型、地形、季节、林隙和树冠等因素的影响[29-32];随细根的不断生长、死亡和分解,其生物量的年际变化也较大。而本研究选择的3个林龄分别仅间隔1 a,供试样株也均为样地内长势较好的植株,且种植于同一样地内,气候差异的影响较小,因此,由于个体间的生长差异造成细根生长量出现上述结果。但林龄间隔对不同树种细根结构差异的影响则有待深入研究。
南方红豆杉细根结构及其与C、N含量均有不同程度的相关性:细根的比根长、比表面积和根尖密度与C含量呈负相关,而与N含量呈正相关,仅差异的显著性不同。细根结构与C和N含量间的关系与林木根系所处的土壤环境密切相关,其中,土壤的物理性质(如温度、容重、含水率等)[33-34]、化学性质及微生物环境均会影响根系的生长与分布。随着土壤养分和水分等条件的改变,C的分配格局以及细根的生理功能也会发生变化[35]。刘苑秋等[36]对退化红壤重建森林初期细根特征及其作用机制的研究结果表明:林木细根分布与土壤有机质、全N和有效K水平及其pH值等指标间存在一定的线性关系。当土壤资源有效性增加时,C向地下部分的分配增加,以促进细根生长和生物量的积累,增强细根吸收水分和养分的能力[37-38]。因此,探讨不同林龄南方红豆杉人工林细根结构与C和N含量的关系以及与土壤物理和化学性质及微生物环境之间的联系,对南方红豆杉人工林的合理经营和管理具有一定的应用意义。
[1] PREGITZER K S.Fine rootof trees——a new perspective[J].New Phytologist,2002,154(2):267-270.
[2] MAKITA N,HIRANO Y,MIZOGUCHI T,et al.Very fine roots respond to soil depth:biomass allocation,morphology,and physiology in a broad-leaved temperate forest[J].Ecological Research,2011,26(1):95-104.
[3] PREGITZER K S,KING JS,BURTON A J,et al.Responses of tree fine roots to temperature[J].New Phytologist,2000,147(1): 105-115.
[4] USMAN S,SINGH S P,RAWAT Y S,et al.Fine root decomposition and nitrogen mineralisation patterns in Quercus leucotrichophora and Pinus roxburghii forests in central Himalaya [J].Forest Ecology and Management,2000,131:191-199.
[5] CALDWELL M M.Competition between root systems in natural communities[M]∥GERGORY P J,LAKE JV,ROSE D A.Root Development and Function.Cambridge:Cambridge University Press,1987.
[6] HENDRICK R L,PREGITZER K S.The dynamics of fine root length,biomass,and nitrogen content in two northern hardwood ecosystems[J].Canadian Journal of Forest Research,1993,23 (12):2507-2520.
[7] 王政权,郭大立.根系生态学[J].植物生态学报,2008,32 (6):1213-1216.
[8] TRUMBORE SE,GAUDINSKI JB.The secret lives of roots[J]. Science,2003,302:1344-1345.
[9] GUO D,LIH,MITCHELL R J,et al.Fine root heterogeneity by branch order:exp loring the discrepancy in root turnover estimates between minirhizotron and carbon isotopic methods[J].NewPhytologist,2008,177(2):443-456.
[10] PREGITZER K S,DEFOREST JL,BURTON A J,etal.Fine root architecture of nine North American trees[J].Ecological Monographs,2002,72(2):293-309.
[11] FITTER A H,STICKLAND T R.Architectural analysis of plant root systemsⅢ.Studies on plants under field conditions[J].New Phytologist,1992,121(2):243-248.
[12] 刘 佳,项文化,徐 晓,等.湖南会同5个亚热带树种的细根构型及功能特征分析[J].植物生态学报,2010,34(8): 938-945.
[13] 洪 伟,王新功,吴承祯,等.濒危植物南方红豆杉种群生命表及谱分析[J].应用生态学报,2004,15(6):1109-1112.
[14] 刘 彤,李云灵,周志强,等.天然东北红豆杉(Taxus cuspidata)种内和种间竞争[J].生态学报,2007,27(3): 0924-0929.
[15] 王 磊,孙启武,郝朝运,等.皖南山区南方红豆杉种群不同龄级立木的点格局分析[J].应用生态学报,2010,21(2): 272-278.
[16] 刘建军.秦岭油松、锐齿栎根系生态研究[M].陕西:西北大学出版社,2002:36.
[17] BAUHUS J,KHANNA P K,MENDEN N.Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii[J].Canadian Journal of Forest Research, 2000,30(12):1886-1894.
[18] BERNTSON G M.Modelling root architecture:are there tradeoffs between efficiency and potential of resource acquisition?[J].New Phytologist,1994,127(3):483-493.
[19] 熊德成,黄锦学,杨智杰,等.亚热带6种树种细根序级结构和形态特征[J].生态学报,2012,32(6):1888-1897.
[20] 常文静,郭大立.中国温带、亚热带和热带森林45个常见树种细根直径变异[J].植物生态学报,2008,32(6):1248-1257.
[21] 师 伟,王政权,刘金梁,等.帽儿山天然次生林20个阔叶树种细根形态[J].植物生态学报,2008,32(6):1217-1226.
[22] 许 旸,谷加存,董雪云,等.海南岛4个热带阔叶树种前5级细根的形态、解剖结构和组织碳氮含量[J].植物生态学报, 2011,35(9):955-964.
[23] 杨小林,张希明,李义玲,等.塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略[J].植物生态学报,2008,32 (6):1268-1276.
[24] 李瑞霞,凌 宁,郝俊鹏,等.林龄对侧柏人工林碳储量以及细根形态和生物量的影响[J].南京林业大学学报:自然科学版,2013,37(2):21-27.
[25] 刘 波,余艳峰,张贇齐,等.亚热带常绿阔叶林不同林龄细根生物量及其养分[J].南京林业大学学报:自然科学版, 2008,32(5):81-84.
[26] PEICHL M,ARAIN M A.Above-and below-ground ecosystem biomass and carbon pools in an age-sequence of temperate pine plantation forests[J].Agricultural and Forest Meteorology,2006, 140:51-63.
[27] 林希昊,王真辉,陈秋波,等.不同树龄橡胶(Hevea brasiliensis)林细根生物量的垂直分布和年内动态[J].生态学报,2008,28(9):4128-4135.
[28] 马 炜,孙玉军,郭孝玉,等.不同林龄长白落叶松人工林碳储量[J].生态学报,2010,30(17):4659-4667.
[29] VOGT K A,VOGT D J,PALMIOTTO P A,et al.Review of root dynamics in forest ecosystems grouped by climate,climatic forest type and species[J].Plant and Soil,1995,187(2):159-219.
[30] IDOL TW,POPEPE,PONDER F,Jr.Fine rootdynamicsacross a chronosequence of upland temperate deciduous forests[J].Forest Ecology and Management,2000,127:153-167.
[31] 朱 慧,洪 伟,吴承祯,等.天然更新的檫木林根系生物量的研究[J].植物资源与环境学报,2003,12(3):31-35.
[32] 杨秀云,韩有志.关帝山华北落叶松人工林细根生物量空间分布及季节变化[J].植物资源与环境学报,2008,17(4):37-40.
[33] 程云环,韩有志,王庆成,等.落叶松人工林细根动态与土壤资源有效性关系研究[J].植物生态学报,2005,29(3):403-410.
[34] 韦兰英,上官周平.黄土高原不同演替阶段草地植被细根垂直分布特征与土壤环境的关系[J].生态学报,2006,26(11): 3740-3748.
[35] BURTON A J,PREGITZER K S,HENDRICK R L.Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forest[J].Oecologia,2000,125(3):389-399.
[36] 刘苑秋,罗良兴,刘亮英,等.退化红壤重建森林初期细根特征及其作用机理研究[J].应用生态学报,2005,16(9): 1735-1739.
[37] KING J S,ALBAUGH T J,ALLEN H L,et al.Below-ground carbon input to soil is controlled by nutrient availability and fine root dynamics in loblolly pine[J].New Phytologist,2002,154 (2):389-398.
[38] GAUDINSKIJB,TRUMBORE SE,DAVIDSON E A,et al.The age of fine-root carbon in three forests of the eastern United States measured by radiocarbon[J].Oecologia,2001,129(3):420-429.
(责任编辑:惠 红,佟金凤)
Com parison on structure and C and N contents of fine root and their correlation analysis of Taxus chinensis var.mairei p lantation w ith different stand ages
LIU Yanyan1a,LIN Han1a,1b,CHAO Lin1a,LIN Yongming1a,1b,CHEN Jianzhong3,WU Chengzhen1a,1b,2,①(1.Fujian Agriculture and Forestry University:a.College of Forestry,b.Key Laboratory of Forest Ecosystem Process and Management of Fujian University,Fuzhou 350002,China;2.College of Ecology and Environment, Wuyi University,Wuyishan 354300,China;3.Forestry Bureau of Jianyang City,Jianyang 354200, China),J.Plant Resour.&Environ.2014,23(3):8-14
Structure parameters and C and N contents of different grade fine roots of Taxus chinensis var. mairei(Lemee et Lévl.)Cheng et L.K.Fu plantation with different stand ages were compared,and correlation between fine root structure parameters and its C and N contents was analyzed.Results show that there are differences in root diameter,specific root length(SRL)and specific surface area(SSA)ofⅠ-Ⅴgrades of fine roots of T.chinensis var.mairei with different stand ages,average root diameters of 6,7 and 8 a plants are 1.073,1.815 and 1.734 mm,average SRL are 14.09,12.88 and 14.12 m·g-1,and average SSA are 54.93,45.85 and 50.72 cm2·g-1,respectively;with enhancing of rootgrade,gradually,SRL and SSA of fine root of same age plants decreasewhile root diameter increaseswith generally significant difference(P<0.05);in general,exceptⅠgrade fine root,SRL and SSA of different grade fine roots of7 a plantall are lower than those of6 and 8 a plants;root diameter of different grade fine roots of6 a plant are the smallest.There are no significant difference(P>0.05)in root-tip density and fork number among different age plants,but those of 6 a plant are the highest.Dry weight of fine root of6 a plant is the lowestwhile that of 7 a plant is the highestwith significant difference.With enhancing of root grade,C content in fine root increases gradually while N content decreases gradually. There are generally significant difference in C and N contents amongⅠ-Ⅴgrade fine roots of the same age plant.In different grades of fine roots,C content in 7 a plant is the highestwhile N content in 8 a plant is the lowest;average C contentof6,7 and 8 a plants is454.41,501.90 and 441.55mg·g-1and average N content is12.63,11.99 and 5.88 mg·g-1,respectively.In fine root,C content has positive correlation with root diameter and dry weight and negative correlation with SRL,SSA and root-tip density;N content has negative correlation with root diameter and positive correlation with SRL,SSA and root-tip density;furthermore,there is no regular correlation in N content with dry weight,in C and N contents with fork number.It is suggested that tree age has a stronger influence on fine root diameter of T.chinensis var.mairei,while it ismore obvious in structure variation of fine rootwith different grades.
Taxus chinensis var.mairei plantation;fine root;structure;C and N contents;stand age; correlation
Q945.12;S791.49.01
A
1674-7895(2014)03-0008-07
10.3969/j.issn.1674-7895.2014.03.02
2014-02-25
国家自然科学基金资助项目(30671664);国家教育部博士学科专项基金项目(200803890010);福建省科学技术厅重点项目(2011N0002)
刘艳艳(1986—),女,山西吕梁人,硕士研究生,主要研究方向为森林生态学。
①通信作者E-mail:fjwcz@126.com
