围封6年对温带典型草原坡地物种多样性及其与地上生物量的关系的影响
2014-05-08徐粒高琼王亚林
徐粒 ,高琼 *,王亚林
1. 北京师范大学地表过程与资源生态国家重点实验室,北京 100875;2. 北京师范大学减灾与应急管理研究院,北京 100875
生物多样性作为生态系统结构的重要度量,对生态系统功能的实现至为重要(Tilman和Downing, 1994; 张全国和张大勇, 2003)。由于研究尺度、研究对象、研究区域的差异,导致不同研究对生物多样性与生态系统功能之间关系的研究结果各异,包括生物多样性导致生态系统功能优化(Tilman和Downing, 1994)、组分有机体功能特点决定生态系统功能(MacGillivray等, 1995)及一些中间观点,但是生态学者一致认为生物多样性对于生态系统的功能及稳定性有着重要影响(张步翀等, 2006)。
物种多样性是反映群落结构的重要指标(郑伟等, 2012),是生物多样性的中心(孙涛等,2007)。生态系统生产力是生态系统功能的重要表现形式(王长庭等, 2005),而植物群落的生产力则是生态系统生产力的基础(江小雷等, 2010)。因而,探讨物种多样性和生产力及其之间关系(张全国和张大勇, 2003),对于阐明生物多样性与生态系统功能的关系及其内在机制有着重要意义(尚占环等, 2005),这也是生态学家研究和讨论的热点问题之一。由于研究尺度、环境条件等因素的差异(尚占环等, 2005),不同研究中物种多样性和生产力之间的关系表现不同,主要表现为单峰型(朱源等, 2007)、线性相关型(覃光莲等, 2002)、无关型(张全国和张大勇, 2002)等。Waide等对约200项相关研究结果统计发现(Waide等, 1999),物种丰富度和生物量的关系中,30%呈单峰关系,26%呈正线性关系,12%呈负线性关系,32%关系不明显。
草原是自然界主要的生态系统之一(韩国栋等,2007),约占全球陆地面积的20%,对于生态环境和人类社会有着重要的影响(白永飞等, 2000)。随着气候变化和人类活动的加剧(左万庆等, 2009),全球范围内草原退化现象越来越严重(王宏等, 2008),草原荒漠化(李博, 1997)、沙漠化、盐渍化、灌木化(Grover和 Musick, 1990)等严重威胁着草原生态系统的持续发展。放牧、刈割和围封等是人类对于草原生态系统的主要干扰方式(Weber等, 2000)。
不同的放牧强度对于草原群落结构和物种多样性有着不同的影响(Scimone等, 2007; Metera等,2010; 郑伟等, 2012)。在轻度放牧下,草场受到较低的外界刺激,群落由少量优势种统治,导致物种多样性降低(Metera等, 2010)。Connell提出“中度干扰假说”(Connell, 1978),认为适度干扰能增加群落和景观的多样性。适度放牧增加多样性的主要机制(江小蕾等, 2003):一是,家畜选择性采食抑制了适口性牧草的生长, 为耐牧性和不适口杂草类植物的生长提供了机会;二是,放牧增加了草地生境异质性,提高了资源的利用率。而过度放牧将会改变土壤理化性质,增大生境异质性,降低物种多样性,改变群落组成结构,导致草场退化(Czeglédi和Radácsi, 2005; 韩国栋等, 2007)。
Peco等人发现(Peco等, 2006),研究区在排除放牧后,约 60%的物种消失。Casasus等人发现(Casasús等, 2007),适当放牧能够抑制灌木的扩张和繁殖,但是它不会导致草地恢复成原有的理想状态,需要和刈割、火烧等措施相结合。由于适度放牧对于生物群落的保护和恢复功能,它已被作为一种重要的手段来恢复欧洲的一些退化草原,其主要任务就是增加物种丰富度(Dolek和 Geyer, 2002;Poschlod和WallisDeVries, 2002)。蒙旭辉等人(蒙旭辉等, 2009)、郑伟等人(郑伟等, 2012)和单贵莲等人(单贵莲等, 2012)的研究均表明,随着放牧强度的增加,物种多样性和群落生产力均表现为先增加后减少的趋势,即符合“中度干扰假说”。
在退化草原恢复重建过程中,围封禁牧以其投资少(单贵莲等, 2010)、见效快等特征(左万庆等,2009),被广泛应用于草原退化的防治。国内外研究发现,尽管在一定时期内,围封可显著提高草地物种多样性和生产力(左万庆等, 2009),但围封时间过长将不利于其维持较高的多样性和生产力(闫玉春和唐海萍, 2007)。
高军靖等人(高军靖等, 2013)研究不同干扰类型对呼伦贝尔草甸草原群落结构及物种多样性影响的比较,发现围封干扰有利于植物物种多样性的维持,而适度的刈割干扰则有利于植物群落的均一化。张伟娜等人(张伟娜等, 2013)对藏北高寒草甸不同管理措施样地调查发现,禁牧5年有利于维持较高的物种多样性和可利用生物量,而5年以上将不利于草场的恢复和利用。朱新萍等人(朱新萍等,2012)发现,中天山草场在围封25年后围栏外的群落丰富度指数和优势度指数均高于围栏内,而均匀度指数围栏内均高于围栏外。
当前,生态学家就放牧围封对草原群落结构和生产力的影响已开展了大量的研究(乔春连等, 2012;朱丽等, 2012)。然而,由于地域、气候等因素的不同(郭轶瑞等, 2007),国内外研究结果存在较大差异。本文以内蒙古太仆寺旗地区典型克氏针茅-羊草草原为研究对象,探究放牧围封对植物群落结构、物种丰富度与生产力(地上生物量)的影响,希望为研究区草原生态恢复重建和草原资源的合理利用提供理论依据。
1 研究区概况
研究区位于内蒙古自治区锡林郭勒盟太仆寺旗境内(北纬 41°35′~42°10′,东经 114°51′~115°49′),海拔1300~1800 m。该地区属中温带半干旱大陆性气候,冬季寒冷干燥,夏季温暖湿润,全年多西北风,年平均风速3.41 m,年平均气温2.1 ℃。最热为每年7月,平均气温21 ℃,最高气温达34 ℃,最冷为1月,平均气温-18 ℃左右,最低气温-36 ℃,全年无霜期100 d左右。最近35年平均降雨量384.5 mm,主要集中在 6、7、8三个月。土壤多为栗钙土,较贫瘠。研究区属内蒙古典型克氏针茅-羊草草原,主要植被类型为克氏针茅 Stipakrylovii、羊草Leymus chinensis、糙隐子草Cleistogenessquarrosa、冷蒿 Artemisia frigida、星毛委陵菜Potentillaacaulis、银灰旋花 Convolvulus ammannii等(单贵莲等, 2010)。典型草原上零星分布有以小叶锦鸡儿Caragana microphylla为主的灌丛斑块。
2 研究方法
2.1 样地设置
于2007年10月在内蒙古自治区锡林郭勒盟太仆寺旗东北黑山庙地区设置坡面围封禁牧样地。沿坡面从上至下400 m,横向50 m范围设置围 栏 (42°06.934′~42°07.100′N, 115°27.743′~115°27.978′E),海拔 1360.5~1395.6 m,坡向为西北-东南向(图 1)。
研究区在2003年前放牧利用至重度退化阶段,2003年后全年轻度放牧利用,每年9月中下旬打草1次。围栏内样地全年禁牧,在和围栏右边缘外相隔5 m的平行坡面位置,选取同样大小对照样地。
2.2 调查内容
2013年7月中旬,将围封样地和对照样地均沿坡上到坡下等分为20个小区,每个小区为20 m×50 m。每个小区内随机选取5个1 m×1 m的草本样方,调查内容包括草本的种类、总盖度、分盖度(针刺法)、高度等,共计200个。
图1 坡地样地设置(平均坡度0.09°)Fig.1 Plot setting of slope (average gradient 0.09°)
2.3 植物生活型功能群划分
根据典型草原植物群落物种组成的特点及植物生活型组成,将生活型功能群划分为:多年生禾草、多年生杂草、半灌木和一年生植物4个类型。
2.4 数据分析
优势度是表征一个种在群落中的地位与作用,本文采用的计算方法如下(张继义与赵哈林等,2004):

式(1)中,DV指物种的优势度(重要值)。
张勇通过调查该围封样地外草本植被的盖度、高度与生物量(张勇, 2011),建立回归关系得出以草本的盖度和高度为自变量,地上生物量为因变量的估测模型:
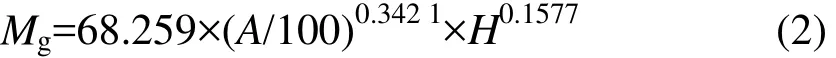
式(2)中,Mg为草本地上生物量(g),A为盖度(%),H为草本高度(cm)。
采用R和Excel2007等软件进行统计分析。
3 结果与分析
3.1 群落物种组成和结构的变化
放牧和围封下群落物种的组成、盖度、高度、生物量及重要值见表1。从表1可以看出,围栏内围封样地中的物种数为 31种,优势种为茵陈蒿、冷蒿、田旋花等,其中多年生禾草、多年生杂草、半灌木、一年生植物的重要值分别为 0.26、0.37、0.26、0.11。围栏外放牧样地中物种数为33种,优势种为冷蒿、糙隐子草、茵陈蒿等,其中多年生禾草、多年生杂草、半灌木、一年生植物的重要值分别为0.22、0.32、0.31、0.15。同时,围栏内各物种平均盖度和高度均显著大于围栏外(P<0.050),而围栏内各物种平均生物量大于围栏外样地,但差异不显著(P=0.059)。
3.2 样地物种丰富度、盖度、生物量沿坡度方向的变化
各功能组物种丰富度(图2)与盖度(图3)围栏内外的空间变化趋势基本一致。用线性回归分析物种丰富度和盖度,结果发现,除半灌木物种丰富度随坡位降低而降低外(其中半灌木物种丰富度未达到统计显著),其他功能组物种丰富度都有随坡位降低而增加的趋势,其中围栏内外多年生杂草和围栏外一年生植物物种丰富度随坡位降低增加显著(P<0.1)。围栏内外多年生禾草盖度随坡位下降而显著下降,而围栏内外一年生植物盖度和围栏内多年生杂草盖度均随坡位下降显著增加,表明功能组盖度与物种丰富度的空间变化不尽一致。
将各功能组物种丰富度和生物量叠加,群落物种丰富度和地上生物量(图 4)均随坡位下降而显著增加,初步显示了物种多样性与生态系统功能的空间变化的一致性。
3.3 围栏内外功能群物种丰富度和盖度的对比
运用方差分析中的配对T检验方法,对比围栏内外功能群物种丰富度、盖度和生物量(表2),结果发现,围栏内草本物种总丰富度显著低于围栏外(P<0.1),而围栏内总生物量却显著高于围栏外(P<0.1)。
围栏内多年生禾草和一年生植物物种丰富度均显著小于围栏外(P<0.1),而围栏内外多年生杂草和半灌木物种丰富度均差异不显著(P=0.454,P=0.298)。
围栏内外多年生禾草、多年生杂草盖度均差异不显著(P=0.432,P=0.317),围栏内半灌木盖度显著大于围栏外(P<0.1),围栏内一年生植物盖度显著小于围栏外(P<0.1)。
围栏内外多年生禾草、多年生杂草生物量均差异不显著(P=0.237,P=0.232)。围栏内半灌木生物量显著大于围栏外(P<0.1),围栏内一年生植物生物量显著小于围栏外(P<0.1)。
3.4 物种丰富度和生物量的关系
运用线性回归分析围栏内外草本半灌木总生物量和物种丰富度(表3),结果发现,围栏内,随着物种丰富度的增加生物量增加,但是关系不显著(P=0.295),而围栏外,随着物种丰富度的增加生物量显著增加(P<0.1)。

表1 围封和放牧下草地群落物种的盖度、高度、频数及重要值Table 1 Coverage,height,frequency and important value of grassland community under enclosure and grazing
4 讨论
4.1 群落物种组成和结构的变化
在干旱、半干旱地区,水分等环境因子是限制群落组成和结构的主要因子,环境因子的改良对群落的作用要大于放牧保护对群落的作用(Howard等,2012)。2013年该地区年降水量269.8 mm,在2013年1月,该地区遭遇30年一遇暴雪,导致在春季植被生长初期土壤水分充足。本研究中,围栏内外的群落物种组成和结构并没有明显差异,围栏内相对围栏外,多年生禾草、多年生杂草、半灌木、一年生植物的重要值分别高出0.04、0.05、– 0.05、–0.04,而围栏内外各物种平均生物量差异不显著。在放牧和刈割样地中,在家畜的采食和践踏作用下,植被的高度和盖度受到限制。周华坤等人(周华坤等, 2004)发现,植被群落的高度、总盖度与放牧强度成负相关。本研究中也发现围栏内各物种的平均高度和盖度均显著大于围栏外。
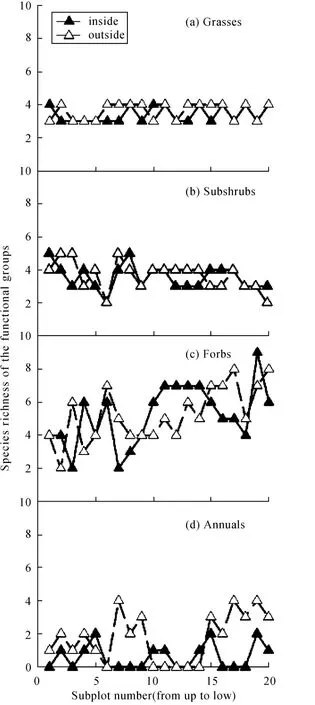
图2 围栏内外植物功能组物种丰富度的空间变化Fig.2 Spatial variation of species richness of the functional groups under enclosure and grazing
图3 围栏内外植物功能组盖度的空间变化Fig.3 Spatial variation of cover of the functional groups under enclosure and grazing
4.2 样地物种丰富度、盖度、生物量沿坡度方向的变化
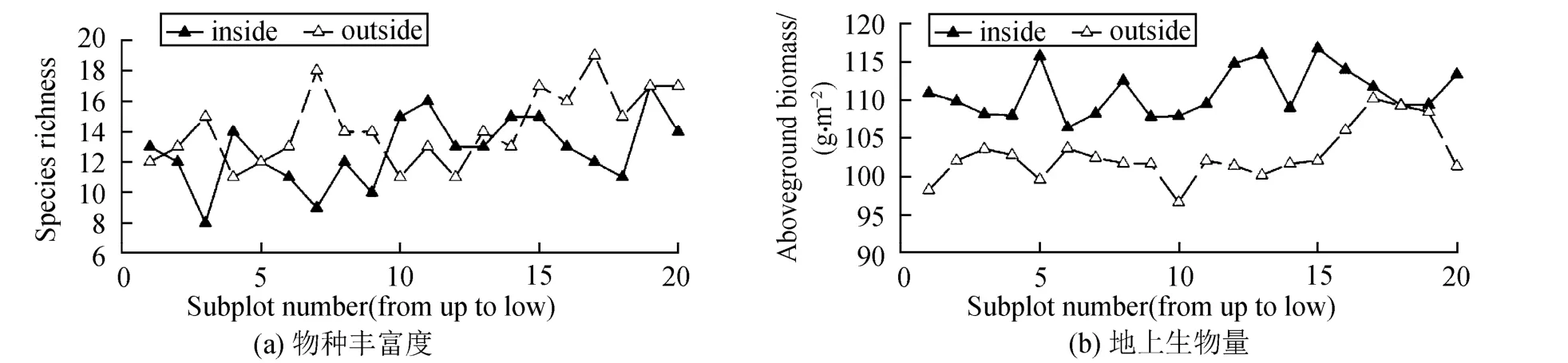
图4 围栏内外物种丰富度与地上生物量的空间变化Fig.4 Spatial variation of species richness and aboveground biomass of the functional groups under enclosure and grazing
在同一坡位上,围栏内外生境相似,导致各功能群物种丰富度与盖度围栏内外的空间变化趋势基本一致。坡面坡度随着坡位的降低而减少,地势变缓,土壤水分等土壤资源增加,植物生境发生改变,导致各功能群物种丰富度和盖度随着坡位的降低而变化,但功能组盖度与物种丰富度的空间变化不尽一致。同时将各功能组物种丰富度和生物量叠加,结果发现,群落物种丰富度和地上生物量均随坡位下降而显著增加,初步显示了物种多样性与生态系统功能的空间变化的一致性。

表2 围栏内外功能群物种丰富度、盖度和生物量的对比(内–外)Table 2 The difference of species richness, cover and aboveground biomass of the functional groups under enclosure and grazing(inside-outside)
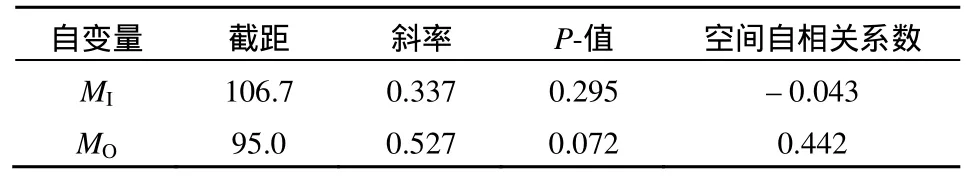
表3 草本半灌木总生物量和物种丰富度的回归方程系数Table 3 The coefficient of regression equation between herbal subshrub total biomass and species richness
4.3 围栏内外功能群物种丰富度和盖度的对比
围栏禁牧样地的物种丰富度要低于自由放牧样地(闫玉春等, 2009)。围封禁牧后,原有优势种补偿性生长,种间竞争加剧,其他低矮植物逐渐减少,导致物种丰富度降低(江小蕾等, 2003)。在放牧样地中,家畜的选择性采食和践踏作用,抑制了适口性牧草的生长,为耐牧性和一年生植物的生长提供了机会的空间,导致物种丰富度升高(袁建立等,2004)。本研究发现,围栏外草本半灌木总丰富度显著大于围栏内,围栏内多年生禾草和一年生植物物种丰富度显著小于围栏外。以冰草为例,围栏内冰草的频数远远小于围栏外样地(表 1,围栏内外频数比 7:26)。
围栏内外在禁牧和放牧不同处理下,表现出不同的草原退化方式。放牧能够抑制灌木的扩张(Metera等, 2010),而长期的围封禁牧导致灌木和半灌木功能的增加,是草原灌木化的表现。Casasús等人(Casasús等, 2007)研究认为,中度放牧有利于抑制灌木植被在草原中扩张。熊好琴等人(熊好琴等, 2011)通过对毛乌素不同围封年限下植被的群落特征的研究,发现围封5年后,油蒿等灌木在群落占据主要地位,而在围封 16年后,草本盖度大大增加而半灌木盖度却减少。本研究中,在围封6年后,围栏内半灌木盖度和生物量显著高于围栏外,表现出灌木化的趋势。同时放牧、刈割增加了耐牧物种和不适口植物生长的可能(罗天相和刘莎, 2007),导致草原退化指示物种增加(刁兆岩等, 2011),是草原退化的表现。围栏外,一年生植物盖度和生物量显著高于围栏内,一年生植物的频数远远大于围栏内(表 1,围栏内外频数比 13:71),且独行菜、猪毛菜等仅出现于围栏外。
4.4 物种丰富度和生物量的关系
草原生态系统功能受物种多样性控制(张全国和张大勇, 2003),但围封禁牧处理影响了两者关系的显著性。自然群落中,多样性和生产力的关系是在演化过程中物种与环境干扰之间相互作用下的产物(马文静等, 2013)。本研究发现,围栏内外草本和半灌木总生物量随物种丰富度增加,但只有围栏外的关系达到显著,表明围封调制了生物多样性与生态系统功能的关系。
5 结论
围封对草地的影响具有双面性(闫玉春等,2009):一方面,合理的围封措施是草地生态保护和恢复的重要手段,对物种多样性、草地生产力等有着深远影响;另一方面,根据草地生态系统的可持续性原理,草地围封不应是无限期的。长期的围封反而不利于草场的繁殖更新,导致草场退化。本研究发现,完全禁牧6年后,尽管生物量有所恢复,但灌木和半灌木的增加却代表了草原的退化。因此,对于干旱半干旱典型草原的恢复,不能简单地采用长期完全禁牧措施,应当根据区域环境、植被以及社会经济情况,制定季节性放牧或者间歇性禁牧的恢复措施,从而保证草场的可持续性利用。
CASASÚS I, BERNUÉS A, SANZ A, et al. 2007. Vegetation dynamics in Mediterranean forest pastures as affected by beef cattle grazing[J].Agriculture, Ecosystems and Environment, 121 (4): 365-370.
CONNELL J H. 1978. Diversity in Tropical Rain Forests and Coral Reefs[J].Science, 199 (4335): 1302-1310.
CZEGLÉDI L, RADÁCSI A. 2005. Overutilization of pastures by livestock[J]. Grassland Studies, (3): 29-35.
DOLEK M, GEYER A. 2002. Conserving biodiversity on calcareous grasslands in the Franconian Jura by grazing: a comprehensive approach[J]. Biological Conservation, 104 (3): 351-360.
GROVER H D, MUSICK H B. 1990. Shrubland encroachment in southern New Mexico, U.S.A. An analysis of desertification processes in the American southwest[J]. Climatic Change, 17 (2): 305-330.
HOWARD K S C, ELDRIDGE D J, SOLIVERES S. 2012. Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrubland[J]. Basic and Applied Ecology,13 (2): 159-168.
MACGILLIVRAY C W, GRIME J P. 1995. Testing Predictions of the Resistance and Resilience of Vegetation Subjected to Extreme Events[J]. Functional Ecology, 9 (4): 640-649.
METERA E, SAKOWSKI T, SLONIEWSKI K, et al. 2010. Grazing as a tool to maintain biodiversity of grassland - a review[J]. ANIMAL SCIENCE PAPERS AND REPORTS, 28 (4): 315-334.
PECO B, SÁNCHEZ A M, AZCÁRATE F M. 2006. Abandonment in grazing systems: Consequences for vegetation and soil[J]. Agriculture,Ecosystems and Environment, 113 (1-4): 284-294.
POSCHLOD P, WALLISDEVRIES M F. 2002. The historical and socioeconomic perspective of calcareous grasslands:lessons from the distant and recent past[J]. Biological Conservation 104 (3): 361-376.
SCIMONE M, ROOK A J, GAREL J P, et al. 2007. Effects of livestock breed and grazing intensity on grazing systems: 3. Effects on diversity of vegetation[J]. Grass and Forage Science, 62 (2): 172-184.
TILMAN D, DOWNING J A. 1994. Biodiversity and stability in grasslands[J]. Nature, 367 (6461): 363-365.
WAIDE R B, WILLIG M R, STEINER C F, et al. 1999. The relationship between productivity and species richness[J]. Annual Review of Ecology and Systematics, 30: 257-300.
WEBER G E, MOLONEY K, JELTSCH F. 2000. Simulated long-term vegetation response to alternative stocking strategies in savanna rangelands[J]. Plant Ecology, 150 (1-2): 77-96.
白永飞, 李凌浩, 王其兵, 等. 2000. 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究[J]. 植物生态学报,(6):667-673.
单贵莲, 徐柱, 宁发. 2010. 典型草原不同演替阶段群落结构与物种多样性变化[J]. 干旱区资源与环境, (2): 163-169.
单贵莲, 薛世明, 陈功, 等. 2012. 季节性围封对内蒙古典型草原植被恢复的影响[J]. 草地学报, (5): 812-818.
刁兆岩, 冯朝阳, 徐立荣, 等. 2011. 草地围封内外植物多样性变化研究:以辉河自然保护区为例[J]. 水土保持研究, (5): 148-151.
高军靖, 吕世海, 刘及东, 等. 2013. 不同干扰类型对呼伦贝尔草甸草原群落结构及物种多样性影响的比较[J]. 环境科学研究, (7):765-771.
郭轶瑞, 赵哈林, 赵学勇, 等. 2007. 科尔沁沙质草地物种多样性与生产力的关系[J]. 干旱区研究,(2): 198-203.
韩国栋, 焦树英, 毕力格图, 等. 2007. 短花针茅草原不同载畜率对植物多样性和草地生产力的影响[J]. 生态学报, (1): 182-188.
江小雷, 岳静, 张卫国,等. 2010. 生物多样性,生态系统功能与时空尺度[J]. 草业学报, (1): 219-225.
江小蕾, 张卫国, 杨振宇, 等. 2003. 不同干扰类型对高寒草甸群落结构和植物多样性的影响[J]. 西北植物学报, (9): 1479-1485.
李博. 1997. 中国北方草地退化及其防治对策[J]. 中国农业科学, (6):2-10.
罗天相, 刘莎. 2007. 中度放牧干扰对草地生物多样性影响的思考[J].安徽农业科学, (21): 6567-6568,6612.
马文静, 张庆, 牛建明, 等. 2013. 物种多样性和功能群多样性与生态系统生产力的关系:以内蒙古短花针茅草原为例[J]. 植物生态学报,(7): 620-630.
蒙旭辉, 李向林, 辛晓平, 等. 2009. 不同放牧强度下羊草草甸草原群落特征及多样性分析[J]. 草地学报, (2): 239-244.
乔春连, 王基恒, 葛世栋, 等. 2012. 围封和放牧条件下高寒矮嵩草草甸土壤性质的比较[J]. 草业科学, (3): 341-345.
尚占环, 姚爱兴, 龙瑞军. 2005. 干旱区山地植物群落物种多样性与生产力关系分析[J]. 干旱区研究, (1): 74-78.
孙涛, 毕玉芬, 赵小社, 等. 2007. 围栏封育下山地灌草丛草地植被植物多样性与生物量的研究[J]. 云南农业大学学报, (2): 246-250+279.
覃光莲, 杜国祯, 李自珍, 等. 2002. 高寒草甸植物群落中物种多样性与生产力关系研究[J]. 植物生态学报, (S1): 57-62.
王宏, 李晓兵, 李霞, 等. 2008. 中国北方草原对气候干旱的响应[J]. 生态学报, (1): 172-182.
王长庭, 龙瑞军, 王启基, 等. 2005. 高寒草甸不同草地群落物种多样性与生产力关系研究[J]. 生态学杂志, (5): 483-487.
熊好琴, 段金跃, 张新时. 2011. 围栏禁牧对毛乌素沙地植物群落特征的影响[J]. 生态环境学报, (2): 233-240.
闫玉春, 唐海萍. 2007. 围栏禁牧对内蒙古典型草原群落特征的影响[J].西北植物学报, (6): 1225-1232.
闫玉春, 唐海萍, 辛晓平, 等. 2009. 围封对草地的影响研究进展[J]. 生态学报, (9): 5039-5046.
袁建立, 江小蕾, 黄文冰, 等. 2004. 放牧季节及放牧强度对高寒草地植物多样性的影响[J]. 草业学报, (3): 16-21.
张步翀, 李凤民, 黄高宝. 2006. 生物多样性对生态系统功能及其稳定性的影响[J]. 中国生态农业学报, (4): 12-15.
张继义, 赵哈林, 张铜会, 等. 2004. 科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态.[J] 植物生态学报, (1): 86-92.
张全国, 张大勇. 2002. 生物多样性与生态系统功能:进展与争论[J]. 生物多样性, (1): 49-60.
张全国, 张大勇. 2003. 生物多样性与生态系统功能:最新的进展与动向[J]. 生物多样性, (5): 351-363.
张伟娜, 干珠扎布, 李亚伟, 等. 2013. 禁牧休牧对藏北高寒草甸物种多样性和生物量的影响[J]. 中国农业科技导报, (3): 143-149.
张勇. 2011. 基于温带半干旱草原围封坡面样地连续观测的植被与土壤水分关系[D]. 北京:北京师范大学.
郑伟, 董全民, 李伟雄, 等. 2012. 放牧强度对环青海湖高寒草原群落物种多样性和生产力的影响[J]. 草地学报, (6): 1033-1038.
周华坤, 赵新全, 唐艳鸿, 等. 2004. 长期放牧对青藏高原高寒灌丛植被的影响[J]. 中国草地, (6): 2-12.
朱丽, 李广宇, 王芳, 等. 2012. 围栏封育对草地生物多样性的影响[J].草业与畜牧, (5): 1-3,29.
朱新萍, 贾宏涛, 蒋平安, 等. 2012. 长期围栏封育对中天山草地植物群落特征及多样性的影响[J]. 草业科学, (6): 989-992.
朱源, 康慕谊, 刘全儒, 等. 2007. 贺兰山高山草甸生物多样性和地上生物量的关系[J]. 应用与环境生物学报, (6): 771-776.
左万庆, 王玉辉, 王凤玉, 等. 2009. 围栏封育措施对退化羊草草原植物群落特征影响研究[J]. 草业学报, (3): 12-19.
