暖温带和北亚热带过渡区白云山植物物种多样性与环境因子相关性
2014-05-08许传阳陈志超郝成元丁效东
许传阳,陈志超*,郝成元,丁效东
1. 河南理工大学测绘与国土信息工程学院,河南 焦作 454000;2. 广东省生态环境与土壤研究所,广东 广州 510650
植被是区域环境的重要组成部分,正确认识其群落组成、结构特征及主要生境因子,对于区域生态建设和生物多样性保护具有非常重要的科学意义;山地地形和气象复杂、植被结构及土壤类型迥异,是植物地理学研究的永恒主题,也是保护生物学的关注焦点之一(Messerli和Ives,1997;Norton等,2003)。多角度、综合研究山地植物多样性的空间分布及生境特征是山地生态学研究的核心内容(方精云等,2004)。
生态学上,植物种类多样性在不同尺度上影响着生态系统的物质流、能量流、信息流,对于生态系统结构、功能、格局、演替都非常重要(宋永昌,2001)。其中,海拔梯度被公认为是温度、水分和光照等环境要素的综合,因而几乎所有的梯度分析大都是以海拔梯度进行(Jankowski和Weyhenmeyer,2006;张昌顺等,2012),但坡度坡向(Stemberg和 Maxim,2001)、大气湿度(Degorski,2003)、土壤有机质(Elgersma和Dhillion,2002)、土壤酸碱度(Partel等,2004)、土壤电导率(Solon等,2007)等其他要素也可能作为主要环境梯度因子引导植物物种多样性发生有规律的变化。然而迄今为止,作为中国南北区域最重要分界的“秦岭——淮河”主要组成部分的伏牛山山地,特别是位于其北坡的白云山国家森林公园的植物种类多样性还尚未见到系统研究报道。段春燕等(2002)通过野外实地调查,整理出白云山国家森林公园珍稀濒危植物21科26种,并列出了名录和主要植被状况;廖秉华等(2008)使用群落生态学方法在伏牛山北坡研究了海拔梯度对植物功能群动态变化及种群相互作用的影响,得出海拔是最重要环境因子的结论;卢训令等(2010)通过设立植物样方、群落调查和多样性分析对物种多样性的垂直分布格局进行了研究,得到了研究区过渡带性质,以及物种丰富度沿海拔升高呈降低态势的负相关关系;陈志超等(2013)基于ArcGIS9.3的空间分析功能,对伏牛山南北两侧的水热因子、土地覆被和净初级生产力时空格局进行了对比分析,得出伏牛山山地效应显著、南北植被分异明显的结论。
鉴于此,本研究从植物物种多样性及其与主要环境因子之间的关系度量出发,探析伏牛山山地效应及其影响下的生境特征与植物种类多样性的分布关系,以揭示山地植被与地理环境之间的相互作用,以期为秦岭山地自然地带过渡性研究提供必要补充。
1 研究区概况
伏牛山为秦岭山脉的东延余脉,相对高差1000~1700 m,被称为“黄河和长江分水岭之一”,范围包括 110°30′~113°05′E、33°10′~34°10′N;呈北西西——南东东走向,东西绵延接近400 km,南北宽 40~70 km。山地以南,年活动积温 4815~4907 ℃,年均降水量931 mm;山地以北,年活动积温3750~4068 ℃,年均降水量832 mm。既位于我国南、北温度带过渡区(暖温带和北亚热带),也处在东、西地形地貌过渡带(第2阶梯和第3阶梯),更是我国中部地区保存最为完整的自然综合基因库,在中国生物多样性研究中占有重要地位(丁圣彦和卢训令,2006)。其北坡的白云山国家级森林公园位于伏牛山腹地,境内海拔1500 m以上的山峰37座,其中玉皇顶(海拔2216 m)为第一高峰;其植物区系起源古老,中国特有、残属种较多,以华北成分为主。
2 研究方法
2.1 样地设置
野外植物样方调查和土壤取样工作已于 2011年5月31日至6月4日完成,共计5天,且土壤采样满足采样前5 d无有效降水的要求。选择人类活动影响较小、林木郁闭度相近且具有地貌代表性及样方可操作性强的地点设置样地,海拔每升高50 m设置一个样地。选择位于伏牛山北坡的白云山玉皇顶路线,在海拔1650 m至2158 m的山体上,地理坐标为东经 111°49´38"~111°50´28"、北纬33°38´25"~33°39´33"之间,土壤类型以棕壤和暗棕壤为主,共设置样地15个,其基本数据见表1。
2.2 指标测定
2.2.1 物种多样性
乔木层样地面积大小为10 m×10 m。同时,在每个乔木层样地的中心及四角分别设2 m×2 m的灌木层样方和1 m×1 m的草本层样方。统计各样地内乔木层、灌木层、草本层植物种类数,其中灌木层和草本层植物种类数的统计结果为5个样地的平均值。计算乔木的物种丰富度[以Margalef指数(D)和物种多样性[以Shannon-Wiener指数(H)表示];其计算公式分别为:D=(S-1)/lnN,式中,S为物种数目,N为所有物种的个体数之和;H=-∑(PilnPi),式中,Pi=Ni/N,Ni为种 i的个体数,N为所在样本的所有物种的个体数之和。
2.2.2 土壤样品采集及性状测定
土壤取样厚度为15~30 cm,每个草本层样地各取100 g,混合后称取100 g作为整个样地的土壤样品。现场测定土层厚度,将样品带回实验室,测定土壤电导率、pH以及有机质、全氮、碱解氮、速效磷、全钾和速效钾含量等8个因子。其中,土壤电导率使用电导率仪测定(土水质量体积比为 1∶5),pH值测定采用电位法(土水质量比1∶2.5),有机质含量测定采用重铬酸钾氧化法,全氮含量测定采用半微量凯氏定氮法,碱解氮含量测定采用氯化钾浸提—分光光度计法,速效磷含量测定采用碳酸氢钠浸提—钼锑抗比色法,全钾含量测定采用酸溶—火焰光度计法,速效钾含量测定采用醋酸铵浸提—火焰光度计法(中国科学院南京土壤研究所,1978;鲁如坤,1999)。
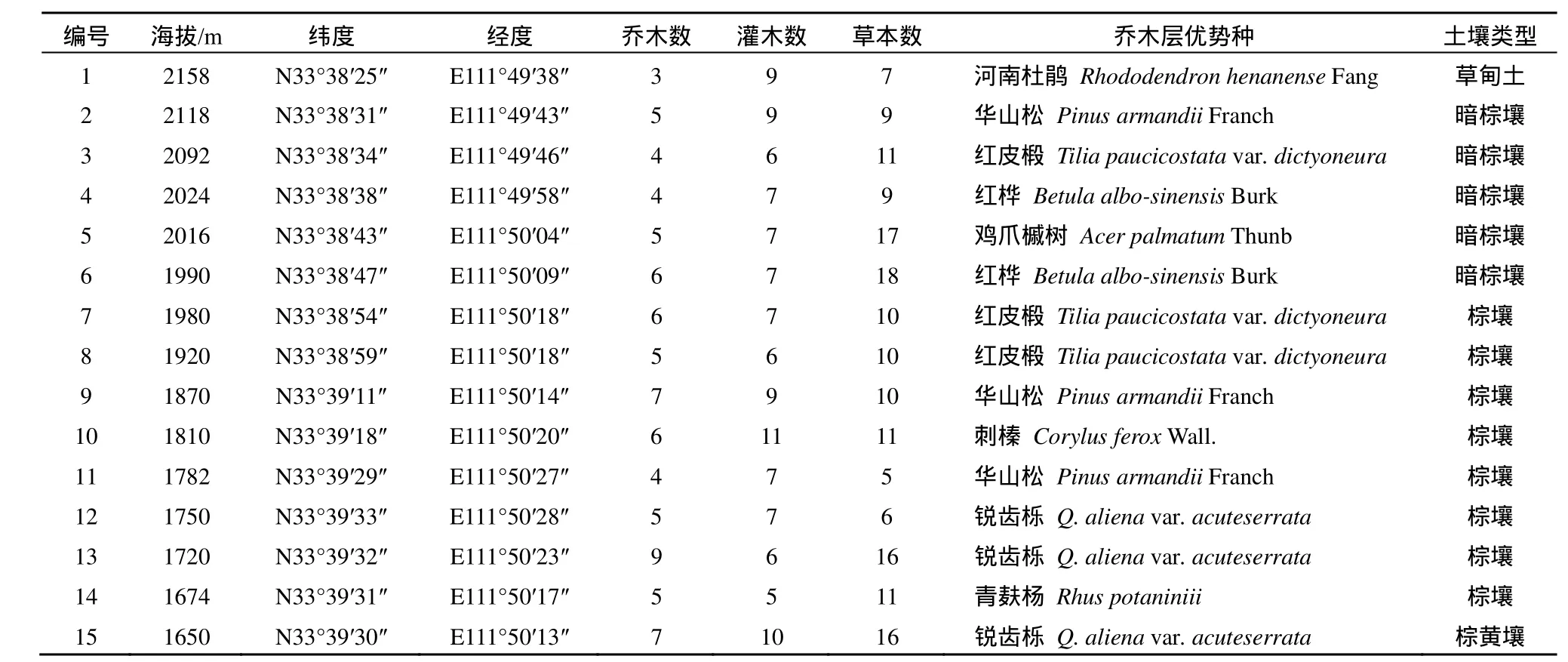
表1 15个样地概况及基本数据Table 1 Description of fifteen study sites and their basic data
2.2.3 气象和地形数据获取
使用LM-8000手持式四合一气象仪(台湾路昌电子企业股份有限公司)现场测定风速、大气湿度、光照度和大气温度等4个因子;收集或计算样地的海拔、坡向、坡度、坡位、坡形、风向坡和阴阳坡等7个因子。其中,风向坡分为向风破、侧风坡和背风坡3类,阴阳坡分为阴坡、中性坡和阳坡3类。
2.3 分析方法
先利用SPSS19.0统计分析软件中的CCA(Canonical correspondence analysis)模块对15个样方内的土壤9个、气象4个、地形7个共计20个环境因子进行主成分分析,筛选出样地生态状况的主要环境因子;然后再应用Pearson相关性分析模块计算物种多样性与其主要环境因子的相关程度。
3 结果与分析
3.1 环境因子的主成分分析
通过主成分分析,把多个环境因子用几个环境因子的综合指标来代替,找出影响样地环境状况的主要因子。在SPSS19.0环境中,对15个样方的20个环境因子原始数据进行主成分分析,结果见表2。
从表2可知,前6个主成分的贡献率分别为30.971%、19.703%、12.570%、9.293%、7.664%和6.276%,累积贡献率为86.476%,其中前3个轴的累积贡献率也达到了63.244%,充分反映出各样方内环境因子的差异。第1轴负荷量较大的4个因子是海拔(0.900)、坡位(0.889)、大气湿度(-0.800)、速效磷(-0.660);第2轴负荷量较大的3个因子是碱解氮(0.863)、有机质(0.723)、坡向(0.729);第3轴负荷量较大的2个因子是土壤厚度(0.756)、大气温度(0.697)。3类要素9个因子中,土壤要素4个因子:土壤厚度、有机质、碱解氮、速效磷;地形要素3个因子:海拔、坡形、坡向;气象要素2个因子:大气温度和大气湿度。因此,土壤、地形、气象3类要素对样地环境状况的贡献值大小依次为地形>土壤>气象。

表2 20个环境因子的主成分分析结果Table 2 Result of principal component analysis between plant species diversity and its twenty environmental factors
3.2 物种多样性与环境因子之间的相关性分析
经过对20个环境因子的主成分分析,从15个样地的20个环境因子中筛选出9个主要环境因子,包括3个地形因子(海拔、坡位、坡向)、4个土壤因子(土壤厚度、有机质、碱解氮、速效磷)、2个气象因子(大气温度、大气湿度),然后对这 9个因子与样地植物物种多样性指数进行相关性分析,得出结果见表3。
3.2.1 植物物种多样性与地形因子相关性
乔木种类数与海拔和坡向呈极显著负相关和显著正相关,其物种丰富度Margalef指数分别与坡向、海拔和坡位显著正相关、极显著和显著负相关,其物种多样性Shannon-Wiener指数亦与坡向、海拔和坡位显著正相关、极显著和显著负相关。灌木种类数仅与坡向呈显著正相关,而草本种类数则与地形因子无明显相关性。总之,海拔、坡位、坡向等地形因子与乔木层种类数、丰富度以及多样性指数相关性显著甚至极显著,与灌木层和草本层种类数无显著相关性。
3.2.2 植物物种多样性与土壤因子相关性
乔木种类数与土壤厚度和速效磷分别呈极显著和显著正相关,其物种丰富度Margalef指数与土壤厚度和速效磷均呈极显著正相关,其物种多样性Shannon-Wiener指数则与土壤厚度和速效磷分别呈极显著和显著正相关。灌木种类数与土壤因子相关性不明显,而草本种类数与有机质、碱解氮仅呈显著正相关。即无论乔木种类数、丰富度和多样性指数均与土壤厚度和速效磷等土壤因子相关性较强,草本种类数则仅与土壤有机质和碱解氮相关性显著,而灌木种类数与土壤因子无显著相关性。
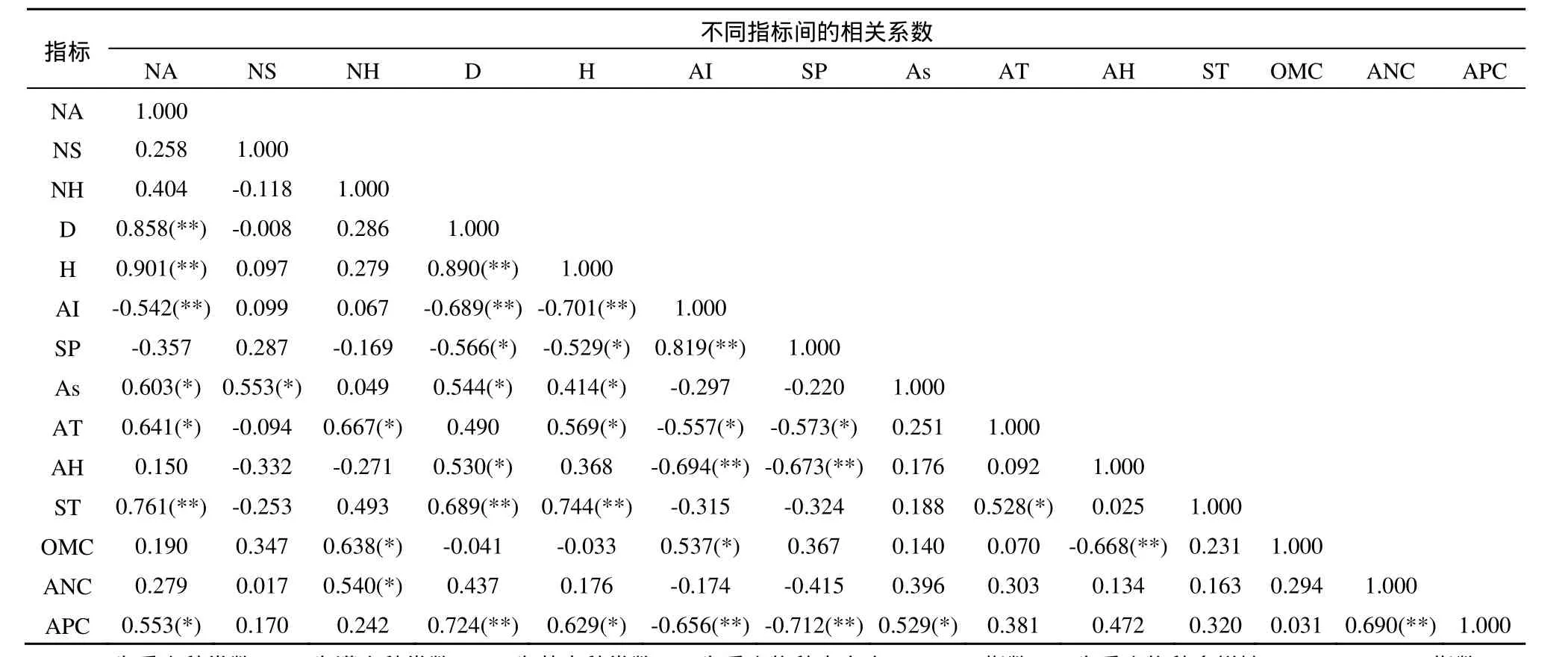
表3 白云山样地植物物种多样性与其主要环境因子的相关系数Table 3 Correlation coefficient between species diversity and main environmental factors on Mt. Baiyunshan
3.2.3 植物物种多样性与气象因子相关性
乔木种类数与大气温度显著正相关,其物种丰富度Margalef指数与湿度呈显著正相关,其物种多样性Shannon-Wiener指数与大气温度显著正相关。而灌木种类数与气象因子无明显相关性,草本种类数也仅与大气温度呈显著正相关。相对来说,乔木、灌木、草本的物种多样性均与大气温度、大气湿度等气象因子相关性不大,甚至无相关性。
总之,白云山植物物种多样性,特别是乔木层种类数、丰富度 Margalef指数以及物种多样性Shannon-Wiener指数与海拔、坡位、坡向等地形因子显著或极显著相关,与土壤厚度、速效磷、有机质等部分土壤性状相关性较强,而与微域气象因子相关性均不明显。
4 讨论和结论
4.1 讨论
环境因子的主成分和相关分析结果表明,地形、土壤和气象是影响植物群落变化的主要环境因子。地形因子中的海拔影响最大、坡向和坡位次之,土壤因子中的土壤厚度和速效磷含量亦是主要控制性要素,而大气温度、大气湿度为主的气象因子对植物物种多样性空间格局影响不大。即海拔、坡向、坡位、土壤厚度、速效磷以及有机质、碱解氮等成为影响植物群落变化的限制性因子,与前人研究结果基本一致(郝占庆等,2003;张婷等,2007;潘红丽等,2009)。特别是乔木物种丰富度和物种多样性与海拔的负相关关系最明显,这一点符合植物群落物种多样性随海拔高度的升高而降低的格局,许多研究都支持这一规律(沈泽昊等,2001;张峰等,2002;段文军和王金叶,2013)。但是分析结果中灌木和草本物种数均与海拔没有显著相关性,可能是因为在研究区中海拔高度在1650 m至2158 m之间的环境异质性不是太大,对灌木和草本的多样性影响较小,在局域环境中不如其他因子的影响那么大有关(南海龙等,2006;杨再鸿等,2007;郑敬刚等,2009)。
4.2 结论
本文以影响中国暖温带和北亚热带过渡区的白云山植物物种多样性变化的环境因子为研究对象,得出其植物物种多样性与其主要环境因子之间的关系特征有如下3点结论。
(1)依据主成分分析结果,影响白云山植物物种多样性空间格局的3类环境要素贡献率大小排布依次为地形要素、土壤性状、气象因子。
(2)乔木层种类、丰富度Margalef指数以及物种多样性Shannon-Wiener指数均与海拔和土壤厚度极显著相关,与坡向和速效磷显著相关。
(3)草本层物种种数与大气温度、土壤有机质、土壤碱解氮等微域气象或局域土壤性状关系密切,而影响灌木多样性变化的主要环境因子尚不明确。
致谢:野外工作得到河南师范大学生命科学学院李发启教授全程技术指导、河南伏牛山国家级自然保护区白云山管理局张培敏高级工程师和焦建峰工程师的鼎力支持,在此一并感谢!
DEGORSKI M. 2003. Pedodiversity as a part of geodiversity in creation of landscape structure. Multifunctional Landscape [J]. Monitoring,Diversity and Management, (2): 105-121.
ELGERSMA T A, DHILLION S S. 2002. Geographical variability of relationships between forest communities and soil nutrients along a temperature-fertility gradient in Norway [J]. Forest Ecology and Management, 158: 155-168.
JANKOWSKI T, WEYHENEYER G A. 2006. The role of spatial scale and area in determining richness-altitude gradients in Swedish lake phytoplankton communities [J]. Oikos, 115(3): 433-442.
MESSERLI B, IVES J D. 1997. Mountains of the world: a global priority[M]. The Parthenon Publishing Carnforth: 2-4.
NORTON J B, SANDOR J A, WHITE C S. 2003. Hillslope soils and organic matter dynamic within a Native American agro-ecosystem on the Colorado plateau [J]. Soil Science Society America Journal, 67:225-234.
PARTEL M, HELM A, INGERPUU N, et al. 2004. Conservation of Northern European plant diversity: with correspondence with soil Ph[J]. Biological Conservation, 120: 525-531.
SOLON J, DEGORSKI M, ROO-ZIELINSKA E. 2007. Vegetation response to a topographical-soil gradient [J]. Catena, 71: 309-320.
STERNBERG M, MAXIM S. 2001. Influence of slope aspect on Mediterranean woody formations: Comparison of a semiarid and an arid site in Israel [J]. Ecological Research, 16: 335-345.
陈志超, 郝成元, 刘昌华, 等. 2013. 伏牛山阻隔作用下水热因子、土地覆被与NPP的时空特征[J]. 生态环境学报, 22(5): 761-766.
丁圣彦, 卢讯令. 2006. 伏牛山和鸡公山自然保护区植物区系比较[J].地理研究, 25(1): 63-64.
段春燕, 侯小改, 王广收, 等. 2002. 白云山国家森林公园珍稀濒危植物资源初步调查[J]. 洛阳农业高等专科学校学报, 22(1): 17-19.
段文军, 王金叶. 2013. 猫儿山自然保护区森林群落垂直格局与主导因素分析[J]. 生态环境学报, 22(4): 563-566.
方精云, 沈泽昊, 崔海亭. 2004. 试论山地的生态特征及山地生态学的研究内容[J]. 生物多样性, 12(1): 10-19.
郝占庆, 郭水良, 叶吉. 2003. 长白山北坡木本植物分布于环境关系的典范对应分析[J]. 植物生态学报, 27 (6) :733-741.
廖秉华, 胡楠, 丁圣彦, 等. 2008. 伏牛山北坡植物功能群组成沿海拔梯度的动态变化[J]. 河南大学学报: 自然科学版, 38(6): 618-621.
卢训令, 胡楠, 丁圣彦, 等. 2010. 伏牛山自然保护区物种多样性分布格局[J]. 生态学报, 30(21): 5790-5798.
鲁如坤. 1999. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社:107-109.
南海龙, 韩海荣, 马钦彦, 等. 2006. 太岳山针阔混交林林隙草木和灌木物种多样性研究[J]. 北京林业大学学报, 28(2): 52-56.
潘红丽, 李迈和, 蔡小虎, 等. 2009. 海拔梯度上的植物生长与生理生态特性[J]. 生态环境学报, 18 (2): 722-730.
沈泽昊, 方精云, 刘增力, 等. 2001. 贡嘎山东坡植被垂直带谱的物种多样性格局分析[J]. 植物生态学报, 25(6): 721-732.
宋永昌. 2001. 植被生态学[M]. 上海: 华东师范大学出版社: 353-360.
杨再鸿, 杨小波, 余雪标, 等. 2007. 海南桉树林林下植物多样性特点的简单相关分析[J]. 浙江林学院学报, 26(6): 725-730.
张昌顺, 谢高地, 陈龙, 等. 2012. 地形和乔木结构对热带北缘与南亚亚热带过渡带林下植被分布的影响[J]. 资源学报, 34(7): 1232-1239.
张峰, 张金屯, 上官铁梁. 2002. 历山自然保护区猪尾沟森林群落植物多样性研究[J]. 植物生态学报, 26(S): 46-51.
张婷, 张文辉, 郭连金, 等. 2007. 黄土高原丘陵区不同生境小叶杨人工林物种多样性及其群落稳定性分析[J]. 西北植物学报, 27(2):340-347.
郑敬刚, 张有福, 王云, 等. 2009. 太行山中段植被分布特征及其多样性研究[J]. 河南科学, 27(3): 292-294.
中国科学院南京土壤研究所. 1978. 土壤理化分析[M]. 上海: 上海科学技术出版社: 62-68.
