丹波黑大豆GmbHLH30转录因子耐铝功能初步研究
2014-04-29宋倩钱绍方陈宣钦陈丽梅李昆志
宋倩 钱绍方 陈宣钦 陈丽梅 李昆志


摘要:铝毒是酸性土壤作物生长的主要限制因素。前期研究发现,铝胁迫下,耐铝型丹波黑大豆SSH(sup-pression subtractive hybridization,SSH) cDNA文库中bHLH30转录因子基因上调表达,推测该基因与丹波黑大豆耐铝性相关。克隆GmbHLH30基因,构建GmbHLH30植物表达载体pK2-35S-GmbHLH30,并在烟草中过量表达获得转GmbHLH30的转基因烟草植株。在铝胁迫下,转GmbHLH30的转基因烟草相对根伸长率比野生型烟草大,可溶性糖和脯氨酸含量高,H2O2水平低。表明GmbHLH30基因的过量表达可以增强植物的耐铝能力,暗示GmbHLH30转录因子参与调控植物的耐铝特性。
关键词:黑大豆:bHLH30; 铝胁迫; 转基因烟草; 铝耐受性
中图分类号:Q945.78
文献标识码:A
文章编号:1007-7847(2014)04-0332-06
铝毒害是酸性土壤限制植物生长的主要因子:铝通常以无毒害的氧化物和铝硅酸盐的形态存在土壤中。然而,当土壤的pH<5时,铝从土壤中溶解出来,形成有毒害的Al3+。当Al离子浓度达到微摩尔浓度时,Al3+就能抑制植物的根生长,影响对养分的吸收,从而影响植物生长。
已有研究证实植物能够通过分泌有机酸、酚类化合物、磷酸等物质在一定程度上抵抗铝的毒害作用。Miyasaka等发现在不含铝的营养液中培养8d的菜豆暴露在铝胁迫条件下时,铝耐受型的菜豆根际的柠檬酸含量是未经铝处理菜豆的70倍;无论铝敏感型菜豆是否经过铝胁迫处理,其根际的柠檬酸含量是铝处理过的耐受性菜豆的十分之一,因此认为柠檬酸能在一定程度上减缓铝对植物的毒害作用。此外,Kidd等报道铝能够诱导玉米中的儿茶酚和黄酮类的酚醛树脂(槲皮黄酮)的释放。
植物对铝的耐受机制,根本上是由遗传机制调控的。在铝胁迫环境条件下,某些基因特异性表达,表达产物可能参与到耐铝信号途径中,或者作为酶调节一些代谢途径,从而增强植物对铝的耐受性。植物转录因子在植物生长发育及其对外界环境的反应中起着重要的作用。碱性螺旋一环一螺旋(basic helix-loop-helix,bHLH)转录因子普遍存在于真核生物中,具有两个功能区。位于N端的碱性区域,又称为亮氨酸拉链区域,主要参与DNA结合;在C端为HLH区域,主要为二聚化区域,形成同源或异源二聚体。bHLH转录因子的亮氨酸拉链区域能够识别DNA的核心序列基序,包括已知的E-box (5'-CANNTG-3'),及最常见G-box(5'-CACGTG-3')。bHLH转录因子参与植物的生长发育过程、植物激素信号途径以及非生物胁迫等过程。拟南芥中的bHLH92转录因子在NaCl、干旱、甘露糖醇以及冷处理实验中都具有较高的表达量,说明了bHLH92能够响应非牛物胁迫。水稻中的RERJ1基因,编码bHLH转录因子,该转录因子能够响应机械损伤和干旱胁迫。酵母中的PHO4的磷酸化与去磷酸化作用控制PHO的调节作用,PHO4是一个bHLH转录因子,它能与第二个转录因子PHO2相互作用,在磷饥饿条件下调节下游基因的表达。然而,bHLH转录因子对抗铝毒性的研究很少见报道。
在先前的研究中,用铝对丹波黑大豆(铝耐受性)和云南小黑豆(铝敏感型)进行胁迫处理,然后通过正向SSH cDNA文库分析,发现在丹波黑大豆的SSH cDNA文库中bHLH30转录因子的基因呈上调表达,但是在云南小黑豆中并没有出现这个转录因子的基因。因此,我们推测GmbHLH30转录因子可能参与到大豆对铝毒的抵御途径中。本研究从丹波黑大豆中克隆出GmbHLH30基因,构建植物表达载体pK2-35S-GmbHLH30,并在烟草中过量表达CmbHLH30基因,对转基因烟草的相对根伸长率、可溶性糖含量、脯氨酸含量以及过氧化氢含量进行测定,以鉴定转基因烟草是否具有耐铝性,从而揭示丹波黑大豆bHLH30转录凶子基因耐铝毒的功能。
1材料与方法
1.1植物材料的培养
本实验采用从日本引进的铝耐受型丹波黑大豆(Tamba)作为实验材料,在25℃恒温条件下将大豆催芽,待其萌发后挑选生长状况一致的豆芽进行水培。选取生长14d形态大小均一致的幼苗,用0.5mmol/L pH4.5的CaCl2溶液预处理12h后,再用50μmol/L pH4.5的AlCl3溶液(含0.5mmol/L CaCl2)分别处理大豆0、2、4、8、12、24h。
本研究所采用的野生型烟草(Nicotianatabacum cv.Xanth)种子均由本实验室所繁殖。烟草种子先用按1:200比例稀释的农药(medience-sporetexnase)浸泡24h杀灭真菌,再加入2mL1% SDS与5% NaClO的混合液进行消毒处理,播种于MS培养基上。
1.2引物合成
根据丹波黑大豆SSH cDNA文库测序所得的GmbHLH30 EST片段,在大豆数据库中查找bHLH基因全长序列,设计引物,在5'一端引物加BamH I酶切位点,3'-端引物加Xho I酶切位点,引物序列如下:基因全长克隆引物:GmbHLH30 5/:5/-GGATCCATGATACAGGAA-GATCAAGG-3/(BamH I),GmbHLH30 3':5'-CTCGAGAAGACCTCGATC-CGTTCC-3'(Xho I);表达谱引物:GmbHLH30 5':5'-GGATCCATGATACACJGAA-GATCAAGG-3',GmbHLH30 3':5'-GTCCAACAAGGGATTCAGA-TA-3'。
1.3GmbHLH30基因的获得
使用TRIzol@Reagent(Invitrogen)提取植物总RNA,并将RNA纯化,然后使用M-MLV反转录酶合成cDNA。采用TIANGEN公司Taq DNApolymerase进行PCR扩增GmbHLH30基因,再用琼脂糖凝胶DNA回收试剂盒回收GmbHLH30基因片段,并使用pMD18-T vector kit (TaKaRa)进行TA克隆;然后用热刺激法转化E.coli DH5α;最后采用蓝白斑筛选阳性克隆。
1.4转基因烟草根伸长的测定
分别取具有4~6条幼根的野生型烟草(WT)、GmbHLH30-10和GmbHLH30-20烟草无菌苗各4株,水培3d后,用0.5mmol/L、pH4.5的CaCl2溶液预处理12h后测定每株幼苗的根长。然后置入50μmol/L pH4.5的AlCl3(含0.5mmol/L CaCl2)溶液处理24h后再次测定其根长。相对根伸长率=(AlCl3处理后根长-AlCl3处理前根长)/CK根伸长。
1.5烟草植株中可溶性糖、脯氨酸和过氧化氢含量测定
分别取生根后1个月左右的WT、Gmb-HLH30-10和GmbHLH30-20烟草无菌苗各3株,置入0.5mmol/L pH4.5的CaCl2溶液预处理12h,再放人50μmol/L pH4.5的AlCl3(含0.5mmol/LCaCl2)溶液中处理24h,用于测定可溶性糖、脯氨酸(Pro)和过氧化氢(H2O2)含量。
采用蒽酮比色法测定可溶性糖含量,以μmol·g-1 FW表示。测定OD625值后代入标准曲线y=0.0345x+0.0204计算可溶性糖含量。
采用磺基水杨酸法测定脯氨酸含量,以μmol·g-1 FW表示.测定OD520值后代入标准曲线y=0.0223x-0.0286后计算脯氨酸含量。
过氧化氢(H2O2)含量采用二甲酚橙法进行测定,以μmol.g-1 FW表示。
1.6数据统计分析
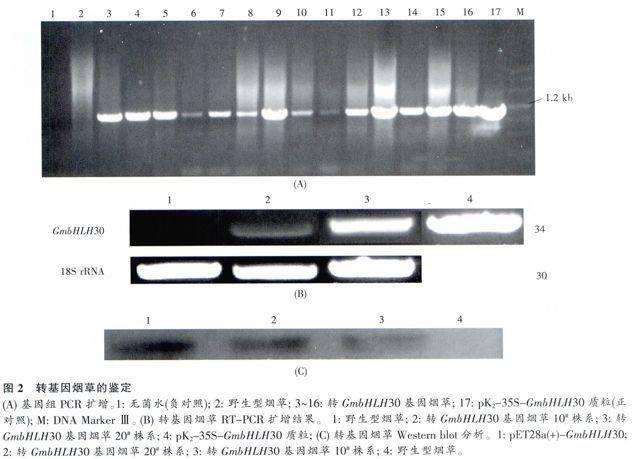
所有生理生化指标测定至少设置3次重复,用TEST法对所得数据进行统计学分析和差异显著性分析。星号表示50μmol/L AlCl3胁迫处理24h后转基因株系和WT株系间生理指标间存在显著性差异(*表示P<0.05,**表示P 2研究结果 2.1不同铝处理时间黑豆根中bHLH30基因表达谱分析 用50μmol/L AlCl3处理丹波黑大豆和云南小黑豆,处理时间为0、2、4、8、12、24h,然后以大豆28S rRNA作为内参,分别对这两种豆根进行RT-PCR分析bHLH30的表达水平。结果显示云南小黑豆处理2h时bHLH30表达量突然上升,4h又下降,8h表达量达到最大,随后表达量呈现下降趋势(图1)。而丹波黑大豆根中bHLH30的表达量开始随着处理时间的增加而增加,当达到8h时,丹波黑大豆bHLH30的表达量达到最大值,之后随着处理时间的增加其表达量逐渐下降(图1)。这表明黑豆根中bHLH30的表达受铝胁迫的诱导。 2.2GmbHLH30过表达转基因烟草的筛选与鉴定 通过Gatewav技术将GmbHLH30基因亚克隆到植物表达载体pK2GW7中构建植物表达载体pK2-35S-GmbHLH30,通过农杆菌介导的叶盘法转化烟草。在基因组水平(图2A)、RNA水平(图2B)和蛋白水平(图2C)上对转基因烟草进行鉴定。在基因组水平检测时,转基因株系GmbHLH30-9、GmbHLH30-10、GmbHLH30-11、GmbHLH30-13、GmbHLH30-20等植株中均检测出了GmbHLH30基因。在这些植株中,选取GmbHLH30-10与GmbHLH30-20株系进行RNA水平与蛋白水平的检测,结果表明GmbHLH30基因成功转入烟草并能够在转基因烟草中成功表达。在后续的实验中均选取GmbHLH30-10与GmbHLH30-20株系作为研究对象。 2.3GmbHLH30过表达转基因烟草的抗铝生理特性分析 在铝胁迫下,铝对植物根部的毒害最明显。为了验证转基因烟草对铝是否具有耐受性,我们对其根相对伸长率进行测定。结果表明在50μmol/LAlCl3胁迫处理24h后,转GmbHLH30基因烟草的相对根伸长率比野生型大,其相对根伸长率比野生型大1.75~2倍(图3A)。这说明转基因植株对铝的耐受性比野生型植株强。 转GmbHLH30基因烟草可溶性糖、脯氨酸含量测定结果见图3B、C。结果表明50μmol/L AlCl3胁迫处理24h后转GmbHLH30基因烟草根和叶中可溶性糖含量都比野生型的高,其中叶中的可溶性糖含量显著增加,而脯氨酸含量在转GmbHLH30基因的植物根中比野生型的低,叶片中则显著增加。这说明铝胁迫下转基因烟草通过调节可溶性糖、脯氨酸含量的变化来维持植物体内渗透压的稳态来提高转GmbHLH30基因烟草的耐铝能力。 在铝胁迫下,通常会导致植物体内产生氧化损伤。通过测定植株体内H2O2含量水平来反映出转基因植株与野生型植株中氧化损伤水平差异。野生型烟草和转基因烟草根和叶中H2O2含量测定结果见图3D。研究结果显示与野生型相比,50μmol/L AlCl3胁迫处理24h后转GmbHLH30基因烟草根和叶中H2O2含量均下降且维持较低水平。说明转基因烟草在铝胁迫下能够存一定程度上清除体内的H2O2,降低氧化损伤的程度,最终提高自身的耐铝能力。 3讨论 前期研究已证明丹波黑大豆的耐铝能力比云南小黑豆强。本研究中,用50μmol/l,的AlCl3处理丹波黑大豆(RB)和云南小黑豆(SB)后对其表达谱进行分析。研究结果表明Al胁迫处理丹波黑大豆0到8h,bHLH基因表达水平逐渐增强,8h达到最大值,随后表达逐渐减弱。云南小黑豆在2h表达量上升,4h下降,到8h达到最大值,之后逐渐减弱(图1)。这可能是由于云南小黑豆对铝胁迫的耐受性较弱,对铝的外部耐受机制也比丹波黑大豆弱。因此在2h处理时,主要依靠诱导内部耐受机制抵御铝毒,随着铝胁迫时间的增加,已表达的GmbHLH30转录因子会调节相关基因的表达.在一定程度上增强植株的耐铝能力,使植株对铝毒的敏感性稍微减弱,所以云南小黑豆处理4h,GmbHLH30的表达量减少。处理8h,云南小黑豆中的GmbHLH30的表达水平较丹波黑大豆中的表达水平高。由于丹波黑大豆的耐铝能力相对较强,其外部耐受机制可能也较强,所以,在0~8h的处理过程中,GmbHLH30的表达量是逐步增加的。在处理时间为8h时,两种豆中GmbHLH30的表达量都达到最大值。但是,随后的12~24h之间,GmbHLH30的表达量都降低。这可能是由于植株可以在一定程度上通过内部耐受机制和外部耐受机制抵抗铝的毒害作用,超过一定的范围后,铝的毒害作用会对植株造成损伤,影响植株正常的新陈的代谢、生理生化过程。从而导致植株整体机能下降,使得处理时间过长,GmbHLH30转录因子表达量会下降。 由于植物的根与土壤中的铝直接接触,在铝胁迫下,铝的毒害作用对植物根部的影响较明显。本研究结果表明转基因植物的根相对伸长率比野生型烟草的大(图3A),说明转基因植物具有抵抗铝的毒害作用,使根部不受损伤,具有较大的相对根伸长率;而野生型烟草对铝毒害作用较敏感,根的生长受到抑制。有文献报道,用铝处理大麦8h后,根尖细胞中的DNA会发生断裂,并且还会形成凋亡体,使细胞发生凋亡。由于细胞死亡,从而抑制根的生长。 在各种逆境胁迫下,植物细胞通过主动积累各种有机和无机物质来提高细胞液浓度,维持膨压,保护质膜的稳定,从而维持原有的生理过程。本实验结果表明,Al胁迫对转基因炯草和野生型烟草的生理特性有显著不同的影响。铝胁迫下转基因烟草根中可溶性糖和脯氨酸的含量都较低,而在叶中的含量比野生型烟草显著的增加(图3B、C),这说明在铝胁迫下转基因植物根受到的危害比较少,具有铝毒抗性。转基因植物叶片上累积较多的可溶性糖和脯氨酸,由于这两种物质都是渗透调节物质,可降低溶质势,保持较高的膨压,维持植物的正常生长,同时这两种物质有利于转基因烟草清除体内的过氧化物质,从而提高转基因植物耐铝毒性。从H2O2含量的生理期指标来看,转基因烟草的根和叶中含量都维持较低水平(图3D),这说明转基因烟草的根部受A1胁迫危害比较少,植物能够正常生长。 综上所述,铝胁迫下转GmbHLH30基因烟草通过累积渗透调节物质可溶性糖和脯氨酸来维持植物体内渗透压的稳态,低的H2O2水平以减少氧化损伤来抵御销胁迫,提高转GmbHLH30基因烟草的耐铝能力。在铝胁迫下,促进了转基因烟草bHLH转录因子的过量表达,可能通过转录凋控可溶性糖和脯氨酸等调节渗透物质的代谢合成和累积以及清除自由氧基相关的靶基因,使烟草叶中含有较高的可溶性糖和脯氨酸含量以及较低的H2O2含量,维持植物膨压和保护细胞膜的结构,达到减轻Al对细胞的毒害,从而使植物在一定程度上忍受、减缓或抵抗逆境胁迫。bHLH转录因子具体调控哪些靶基因还有待进一步研究。
