高山被孢霉△6一脂肪酸脱氢酶基因在红冬孢酵母中的表达
2014-04-29李凌彦胡彬彬赵吉然魏云林林连兵季秀玲张琦
李凌彦 胡彬彬 赵吉然 魏云林 林连兵 季秀玲 张琦


摘要:以绿色荧光蛋白(green fluorescent protein,GFP)作为报告基因,将质粒pRH2304转化红冬孢酵母YM25235进行表达分析,荧光显微观察结果表明GFP在YM25235获得表达,建立了红冬孢酵母YM25235遗传转化方法 在此基础上,以高山被孢霉△6一脂肪酸脱氢酶基因取代pRH2304中的GFP基因,构建重组质粒pRH2304MAD6,将其转化红冬孢酵母YM25235进行表达分析。PCR结果表明,高山被孢霉△6一脂肪酸脱氢酶基因已经整合到YM25235基因组中,进一步的脂肪酸气相色谱分析结果表明,该基因编码产物催化n-6途径中的亚油酸转化成γ-亚麻酸,占细胞总脂肪酸的4.35%,但没有检测到催化n-3途径中的α-亚麻酸转化成十八碳四烯酸。
关键词:红冬孢酵母;高山被孢霉;△6一脂肪酸脱氢酶基因;γ一亚麻酸;表达
中图分类号:Q786
文献标识码:A
文章编号:1007-7847(2014)04-0304-06
多不饱和脂肪酸(polyunsaturdted fatty acids,PUFAs)是指含有两个或两个以上双键、碳原子数为18~22的直链脂肪酸,除了作为结构成分和信号分子对维持细胞膜正常功能和一些基本的细胞过程具有重要的调节作用外,PUFAs在人体内合成变化会导致多种疾病,比如心血管疾病、肥胖、非胰岛素依赖性糖尿病、高血压、神经性疾病和癌症等。PUFAs按照脂肪链甲基端引入的最后一个双键位置分为n-3和n-6两大类,亚油酸LA(linoleic acid C18:2,n一6)和α-亚麻酸ALA(α-Linolenic acid, 18:3 n-3)是n-6和n-3 PUFAs合成的前体,两者在不同脂肪酸脱氢酶(Desaturase)和延长酶(Elongase)的交替催化下分别转化成γ-亚麻酸GLA(γ一Linolenic acid, C18:3 n-6)、花生四烯酸AA (Arachidonic acid, C20:4 n-6)、EPA(Eicosatetraenoic acid, C20:4 n一3)和DHA (docosa-hexaenoic acid, C22:6 n-3)等多种PUFAS。同一种脱氢酶或者延长酶都可利用n-3和n-6途径中的脂肪酸作为底物进行催化,但是由于酶的底物偏好性特点会导致一条途径的脂肪酸转化增强,而另一条途径的脂肪酸转化减弱。
某些微生物产生并储存的油脂占其生物总量的20%以上,最高可达细胞干重的80%,这类微生物称为产油微生物。目前已报道的产油酵母包括耶罗维哑酵母属(Yarrowia)、假丝酵母属(Candida)、红酵母属(Rhodotorula)、红冬孢酵母属(Rhodosporid-ium)、隐球酵母属(Cryptococcus)、毛孢子菌属(Tri-chosporon)和油脂酵母属(Lipomyces)等一些种类,这些产油酵母油脂中富含PUFAs,而且油脂成分与常见的植物油相类似,因此可作为生活中植物食用油的替代品,具有重要的潜在应用价值,但是不含GLA。GLA是一种人体必须脂肪酸,是合成AA的前体,属于n-6类PUFAs,AA可进一步转化成前列腺素、白细胞三烯以及脂氧合酶或是环氧合酶的产物,这些产物在降血脂、抗脂质过氧化、抗炎、减肥、抑制过敏、增强胰岛素和抗血栓性心血管疾病等方面具有重要的生理功能,但是现有GLA商业来源难以满足日益增长的市场需求,借助于基因工程手段构建产高GLA菌株可作为潜在的替代来源之一。本研究将高山被孢霉(Mortierella alpina)△6-脂肪酸脱氢酶基因转化到产油酵母——红冬孢酵母(Rhodosporidium, Kra-tochvilovae)YM25235中进行表达,使其产GLA,类似的研究在红冬孢酵母属中未见报道。
1材料和方法
1.1材料
1.1.1菌株、质粒和培养条件
红冬孢酵母(Rhodosporidium Kratochvilovae)菌株YM25235由昆明理工大学嗜极微生物研究室保存,质粒pRH2304(图1)购自新加坡民族大学,具有高山被孢霉(Mortierella alpina)△6一脂肪酸脱氢酶基因(MAD6,GenBank: AF307940)的T载体pTMAD6由南开大学李明春教授惠赠;菌株YM25235在YM培养基(酵母膏0.3%、胰蛋白胨0.5%、麦芽糖0.5%,pH自然)中30℃条件下培养。
1.1.2主要试剂
酵母总DNA提取试剂盒为上海生工产品DNA聚合酶、各种限制性内切酶、T4连接酶和DNA胶回收试剂盒购自大连宝生物工程公司,潮霉素以及油脂分析所用的油酸、亚油酸、γ-亚麻酸和α-亚麻酸甲酯标准品均购自美国Sigma公司。
1.2红冬孢酵母YM25235感受态细胞制备
挑取YM25235单菌落接种于5mL YM液体培养基中,150r/min、30℃培养过夜,以1%接种量转接到100mL YM液体培养基中,150r/min、30℃培养至OD=1.3~1.5,4℃、5000r/min离心10min收集菌体,用100mL预冷的无菌水洗涤菌体两次,然后用20mL lmol/L预冷的山梨醇洗涤1次,最后悬浮于200μL lmoI/L预冷的山梨醇中待用。
1.3pRH2304质粒转化
取5μg pRH2304,通过电击法转化80μL制备的感受态YM25235,然后涂布于含400mg/L潮霉素的YM固体抗性平板上并置于30℃培养直至阳性克隆出现,最后通过荧光显微镜用油镜观察RtGFP基因是否在YM25235细胞中表达。
1.4pRH2304MAD6重组表达质粒的构建
参考已报道MAD6基因序列,设计合成引物对:MAD6F:5'-CATACCATGGCTGCTGC TC-CCAGTGTGA-3'(下划线为BamH I位点)和MAD6R:5'-CTCGATATCTTACTGTG CCTTGCC-CATC-3'(下划线为EcoR V位点),以含有MAD6基因的T载体作为模版进行PCR扩增,PCR扩增条件为:95℃预变性2min,95℃变性1min,56℃退火1min,72℃延伸1.5min(30个循环);72℃再延伸10min。扩增产物用BamH I/EcoR V进行酶切并回收酶切产物,同时用BamH I/EcoR V酶切pRH2304并回收大片段,用T4连接酶将两种回收产物连接并转化大肠杆菌DH5a,由此获得重组质粒为pRH2304MAD6,MAD6基因取代原来GFP(green fluorescent protein)基因,所得质粒经酶切分析和测序,验证其正确性。
1.5pRH2304MAD6的质粒转化
按1.2中的方法将重组质粒pRH2304MAD6转化感受态YM25235,涂布于含400mg/L的YM固体抗性平板上培养,挑取抗性克隆进行液体培养,用试剂盒进行基因组DNA提取,用引物对MAD6F和MAD6R阳性克隆的筛选,同时用引物对RKD12F:5'-ATGGCCGCTAC CCTCCGCCAGC-3'和RKD12R:5'-CTAGAGTCCCTCGACGCCCGAG-T-3'来扩增YM25235 △12-脂肪酸脱氢酶基因组序列来证明所获得的阳性克隆为YM25235菌株。
1.6脂肪酸分析
将转基因酵母种子液接种于50mL YM液体培养基中,置于30℃、150r/min培养48h。离心收集酵母细胞,50℃,烘干后研磨,取100mg干菌粉加入5mL 5%的KOH-CH30H溶液中,70℃反应3~5h。冷却到室温后,用6mol/L的盐酸调节溶液的pH值为2.0,加入4mL 14% BF3(BF3/乙醚)-CH3OH溶液,70℃继续反应1.5h,合成脂肪酸甲酯。再加入饱和的NaCl溶液10mL,充分混匀后转移到分液漏斗中。用8mL l:4的氯仿:环己烷抽提两次,合并提取液,然后加入适量无水Na2S208干燥提取液,静置1h,过滤去除Na2S2O8,用氮气将含有脂肪酸甲酯的滤液吹干,正己烷回溶后通过气相色谱(GC)检测转基因红冬孢酵母YM25235脂肪酸含量的变化。
2结果
2.1红冬孢酵母YM25235质粒转化体系的建立
将pRH2304通过电击法转化红冬孢酵母YM25235,通过荧光显微镜检测具有潮霉素抗性的阻性克隆,结果显示pRH2304质粒中的RtGFP基因在YM25235中获得表达,使细胞发出绿色荧光(图2B),表明pRH2304可以转化红冬孢酵母YM25235并使目的基因获得表达,为下一步高山被孢霉MAD6基因的表达鉴定了基础。
2.2重组表达质粒pRH2304MAD6的构建
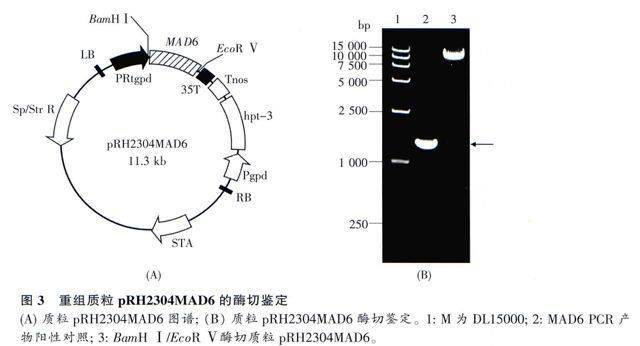
通过PCR、酶切和连接方法,将MAD6基因序列插入pRH2304启动子PRtgpd和终止序列35T之间,取代原来的RtGFP基因序列,构建出重组表达质粒pRH2304MAD6(图3A)。用BamH I/EcoR V对pRH2304MAD6进行双酶切结果显示,小片段与MAD6基因的PCR产物大小一致(图3B,箭头所示),而大片段约为11kb,大小与预期相符,说明质粒构建正确,进一步的测序结果也证明这一点。
2.3红冬孢酵母YM25235转化子的鉴定
将重组质粒pRH2304MAD6通过电击法转化红冬孢酵母YM25235,提取潮霉素抗性的克隆子基因组DNA,用引物对RKD12F和RKD12R进行PCR扩增,结果显示,从抗性克隆子基因组DNA中也能扩增出1.6kb的△12-脂肪酸脱氢酶基因组序列(图4A),说明该克隆子为红冬孢酵母。进一步用引物对MAD6F和MAD6F进行PCR扩增,结果显示抗性克隆子基因组DNA中能扩增出1.4kb的MAD6基因的序列(图4B),表明pRH2304MAD6已经转化到YM25235中并已整合到其基因组中。
2.4红冬孢酵母YM25235阳性转化子的脂肪酸分析
将重组质粒pRH2304MAD6转化YM25235,经验证正确后提取转基因酵母细胞的总脂肪酸进行GC分析。如图5所示,以pRH2304转化的酵母菌细胞作为对照,只有转化了MAD6基因的酵母YM25235中产生保留时间为15.13min的新的特殊峰(图5B,黑色箭头所示),该峰保留时间与GLA甲酯标准品的一致,表明合成了新的GLA,其合成量占细胞总脂肪酸的4.35%(表1)。
3讨论
△6-脂肪酸脱氢酶能同时催化n-6和n-3途径中底物LA和ALA生成GLA和OTA,但是一些研究表明该酶对n-6和n-3途径中的底物具有偏好,比如苔藓(Ceratodon purpureus)△6-脂肪酸脱氢酶对n-6途径中底物LA具有很强的偏好,而樱草(Primula vialii)和少根根霉(Rhizopus ar-thizus)的△6-脂肪酸脱氢酶却偏好n-3途径中的ALA。红冬孢酵母YM23235自身能合成LA和ALA,两者均可作为△6一脂肪酸脱氢酶的潜在底物,但是从我们的分析结果看出,高山被孢霉△6-脂肪酸脱氢酶转化的酵母油脂中只合成了GLA,没有检测到OTA,可以说明高山被孢霉△6-脂肪酸脱氢酶对n-6途径中底物LA具有很强的偏好。目前,除了少根根霉(Rhizopus arrhizus)的△6-脂肪酸脱氢酶外,未见丝状真菌△6一脂肪酸脱氢酶偏好n一3途中的ALA作为底物合成OTA的研究报道。
人体中△6-脂肪酸脱氢酶可将外源性摄人的LA转化成GLA并进一步转化成AA等其他n-6类PUFAs,来满足机体的各种需要,然而老龄化、营养缺乏、吸烟和过度酗酒等原因会导致△6一脂肪酸脱氢酶活性降低,导致GLA合成不足,引起多种疾病。因此,必须在食物中添加外源性的GLA,但现有GLA商业来源难以满足日益增长的市场需求,通过基因工程手段使微生物生产特殊的油脂是另一种重要的潜在来源。Huang等尝试将△12和△6-脂肪酸脱氢酶在酿酒酵母(Saccha-romyces cerevisiae)和耶罗维亚酵母(Yarrowialipolytica)中共表达产生GLA,我们前期的工作也分别将高山被孢霉和少根根霉△6一脂肪酸脱氢酶基因导入毕赤酵母中表达,但是脂肪酸转化率低、油含量低以及组分复杂等原因使得进一步应用难以推广。作为潜在的食用油商业化来源之一,产油酵母其油脂成分与常见的植物油类似,不含GLA等高不饱和的PUFAs。本研究通过基因工程手段将高山被孢霉△6-脂肪酸脱氢酶基因导入产油酵母YM25235中进行表达,使其产GLA,改良其油脂成分和营养价值,为将来大规模应用打下基础。
