灰花纹鹅膏茵(Amanita fuliginea)子实体发育及核相观察
2014-04-29郝小花田连福杨校谭虹张志光李东屏
郝小花 田连福 杨 校 谭 虹 张志光 李东屏
摘 要:采用石蜡切片、压片等方法对灰花纹鹅膏菌(Amanita fuliginea)子实体发育过程进行了观察,并用核荧光染料Hoechst 33258对灰花纹鹅膏菌子实体发育过程中的核相变化进行了观察。结果表明,灰花纹鹅膏菌子实体发育是典型的半被果型发育,菌褶是子实层同时发育类型,属于同型菌髓。在于实体发育过程,单倍的二核担子核配后形成短暂的二倍体单核担子,经过减数分裂形成四核担子,有近40%的担子中核再进行一次有丝分裂形成8核担子,担子进一步发育产生担孢子,其中,二核担孢子约占95%,单核担孢子约2.4%,三核孢子2%左右,4核担孢子约0.4%。灰花纹鹅膏菌基因组中染色体数目2n=10条。
关键词:灰花纹鹅膏菌;子实体;核相:染色体
中图分类号:Q942.4
文献标识码:A
文章编号:1007-7847(2014)06-05ll-05
灰花纹鹅膏菌(Amanita fuliginea Hongo)是一种剧毒蘑菇,近年来在我国南方地区发生了数十起误食该菌引起的中毒事件,中毒人数352人,其中死亡 79。其所含的鹅膏肽类毒素是致死毒素,同时该毒素足生命科学研究中重要的试剂,在抗肿瘤等牛物医学领域也有潜在的用途。我们在对其所产牛的毒素、毒理及中毒治疗等进行研究的同时,亦在生理生态、细胞发育等进行研究。鹅膏菌类真菌,由于不能人工栽培,取材困难,因此,其发育过程的研究极少有报道。而且,灰花纹鹅膏菌像大多数其他担子菌一样,染色体很小,难于用一般的染色方法进行核相和染色体数日观察。以往一股采用繁琐的苏木精或吉姆萨染色法,重现性较差,且研究的种类也主要见于易于培养和取材的担子菌,如草菇、平茹、香菇、鬼伞等。本文采用石蜡切片和荧光染色方法,对灰花纹鹅膏菌子实体发育及担子减数分裂过程中核相变化进行观察,该研究为进一步闸明灰花纹鹅膏菌的发育机制,进而为实现灰花纹鹅膏菌的人工栽培提供了理论参考。
1 材料与方法
1.1 材料
灰花纹鹅膏菌(Amanita fuliginea)菌丝体、不同发育阶段的子实体采集于湖南省长沙县。卡诺氏固定液、荧光染料Hoechst 33258 (Bisbenzimide,Sigma,美国)、磷酸缓冲液(pH 6.8)。
1.2 方法
子实体的发育观察采用石腊制片方法,横切或纵切.切片厚度5μm。
担子核相观察采用荧光染色法。将子实体少许置载玻片上,用摄子捣碎,滴一滴染液,混匀,盖好盖玻片,静置10 min,在荧光显微镜下观察。
2 结果与分析
2.1 灰花纹鹅膏茵子实体的发育观察
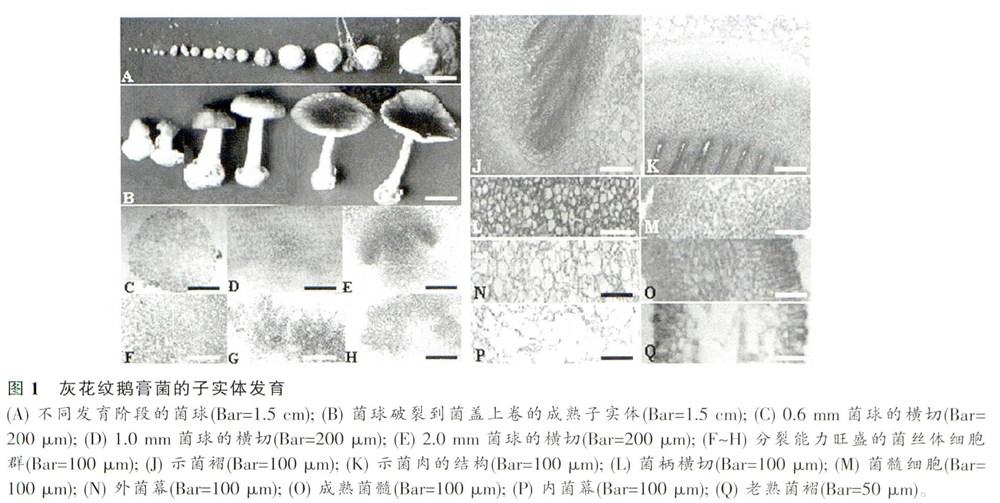
土壤中灰花纹鹅膏菌的营养菌丝发育到一定阶段后,某些部位的菌丝会产生分化,向心生成直径0.5 mm左右的白色菌球,结构上较疏松,由直径3~4 μm的菌丝体扭结而成,内外菌丝形态上无甚变化(图1A、C、F),但菌球中心点细胞是分裂活性中心。继续发育至直径lmm左右的菌球时,内外层菌丝体形态上开始出现分化。在菌球中心处产生结构致密的球状原基。该群细胞较细短,因相互挤压而使菌丝细胞变得有些弯曲,其内原生质浓,核荧光显色显示细胞核分裂旺盛(图ID、G)。外层的菌丝细胞略粗,排列较疏松,核相清晰,菌丝问夹杂少量膨大的菌丝细胞球,组成原始外菌幕(图1D)。随着球状原基内层细胞不断分裂增多,其外围细胞也不断生长膨大,形成另一层由球形细胞和菌丝细胞组成的中层,愈是向外.其球形细胞的比例愈高,体积愈大,最后发育成外菌幕(图1E),原始外菌幕在菌体发育中被撕毁。而球状原基内层细胞的不等分裂,逐步发育分化出盖原基和柄原基(图1E、H)。盖原基进一步发育出现菌肉原基,子实层体原基和盖表皮原基以及内菌幕,随后在子实层体原基中开始形成子实层,子实层出现前无菌褶腔形成,子实层体原基长出侧生菌丝,几次近端分叉,末端细胞即为原担子,大量原担子紧密而规则地排成层即构成子实层(图1J~K)。子实层是同时发育类型。其褶缘囊状体是假囊状体,是与子实层原基同一来源的菌髓末端细胞膨大,液泡化的结果。子实层上无侧缘囊状体。随着子实层的发育及菌盖的膨大,菌柄也逐渐加粗伸长。在菌盖展开菌柄伸长中,外菌幕被撕开残留于菌柄基部一起构成菌托,内菌幕发育成菌环(图1B),菌环由两种排列方向的菌丝细胞组成,靠菌褶的一层菌丝体来源于褶缘不育菌丝,靠近菌柄的一层菌丝体来源于内菌幕。
2.2 子实体发育过程中核相变化及染色体数目观察
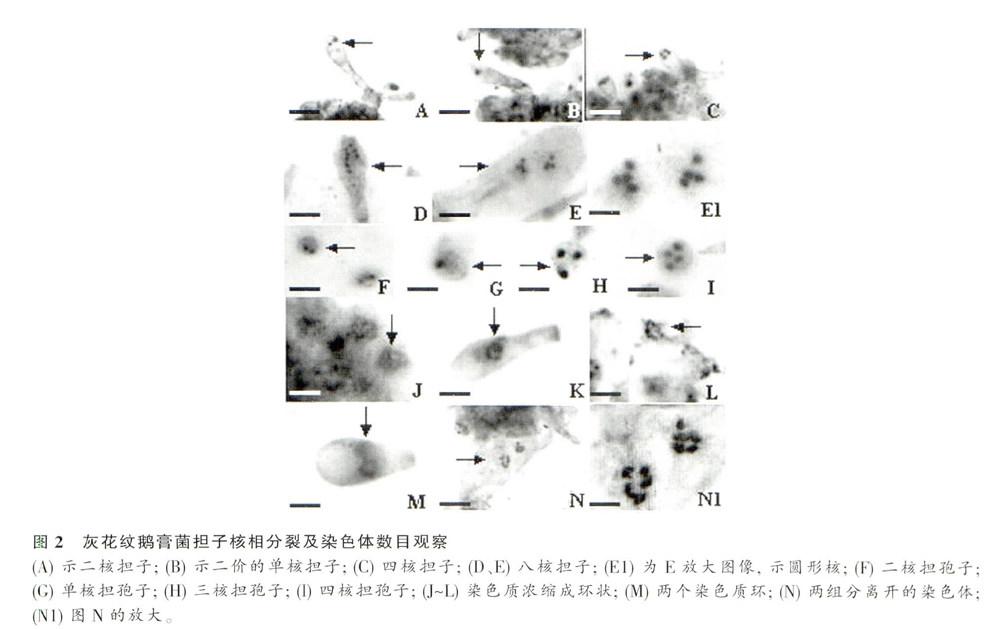
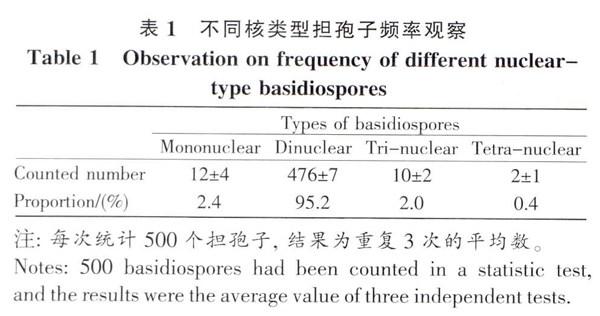
灰花纹鹅膏子实体发育过程中,只有担子发生减数分裂,像大多数担子菌一样,由二核担子(图2A)的两个单倍性的核发生核配,融合为一个二倍性核的单核担子(图1B),紧接着发生减数分裂变为单倍性的四核担子(图2C)。约有40%的担子为8核(图2D~EI),是四核担子再进行一次有丝分裂所致。但也有少量5核~7核担子;担子发育的过程中,会在顶端产生4个担孢子,担子中的核会迁移到担孢子中。95%以上的扭孢子为二核(图2F),少数担孢子为单核和三核孢子(图2G~H),偶见四核担孢子(图2I)(表1)。二核单孢子产生一方面是8核担子中8核进入4个担孢子.每个担孢子二核;另一方面是因为4个单核孢子产生后,在小梗上各进行了一次有丝分裂所致。而单核、三核和四核担孢子,是由于8核担子在进入担孢子中的少数出错行为。
担子在减数分裂过程中染色体排列呈现典型“土星环”结构(图2K—N)。图2J为减数第一次分裂的粗线期前后;图2K为单土星环的结构;图2L示减数分裂中后期两组姊妹染色单体成环状呈一定角度分离;图M示减数分裂后期分开的两组染色单体在担子中呈双环结构;图2N、Nl示担子减数分裂至终变期,两组二价染色单体,n=5。因此,灰花纹鹅膏基因组中二倍体染色体数目为10条。
担子减数分裂发生频率与其发育阶段有密切联系。由表2可以看出,菌球期的担子主要为核配前担子(88.2%),剩下11.8%为已发生核配的担子;发育到伸长期菌球破裂阶段,则大部分担子停留在核配期(53.9%),少部分已到减数分裂期(6.5%);而到伸长期菌球半开阶段,各种类型的担子都可以观察到,其中57%的担子处在减数分裂期,便于进行分裂相和染色体数目观察。至菌盖平展的成熟期,部分担子已产生担孢子,核已迁移到担孢子中。在菌盖边卜卷成熟期,绝大部分的担子都已发生担孢子。因此,要观察灰花纹鹅膏菌担子的核相和核分裂相,选择菌球半开的伸长期的子实体为材料比较适宜。
3 讨论
担子菌个体发育的形式十分多样,正确认识其子实体生长发育形成过程,对于分种及属内系统分类有重要意义。鹅膏菌属真菌的开裂子实层发育方式(schizohymenium)在伞菌目中是独一无二的,其子实层及子实体的具体发育过程至今尚无确切的结果。Corner (1994)认为本属真菌子实层体的形成方式与其他伞菌相同,即菌褶的形成是通过离心隆起(centrifugae ridges)实现的,隆起的下端(褶缘)是分生中心,杨祝良(1995)对毒蝇鹅膏(A.muscaria)和变红鹅膏(A.rubescens)的个体做了显微观察,认为褶缘并非子实层分生区。他认为子实层发生有先后之分,它在子实层原基中相隔一定距离不同点启动后向菌内或相反方向推进,之后连成褶片。而我们对子实体进行连续切片观察表明,灰花纹鹅膏子实层的发生无明显先后之分,褶缘细胞并不形成担子,它们是与菌柄菌幕同一起源的菌髓细胞膨大形成;其附近的子实层也不是分生中心。菌褶的形成很可能亦不是离心隆起方式实现的;因为刚分化出菌褶原基时,菌髓细胞与菌肉细胞和菌柄细胞是紧密相联的细长菌丝体,结构上无显著差异,随着盖原基的发育,菌髓菌丝细胞逐渐与菌肉细胞及菌柄菌幕细胞垂直,由于菌丝细胞分裂变长的速度快于变粗的速度,菌肉组织的横向增长快于子实层体原基的增粗,因此出现菌褶腔。辐射状排列的菌褶形成依赖于基因时序表达的精确调控。
灰花纹鹅膏菌担子发育,一般形成4个二核担孢子。观察灰花纹鹅膏菌的担孢子,绝大多数为二核担孢子,应是灰花纹鹅膏菌正常发育的担孢子类型;在灰花纹鹅膏菌担孢子发生过程中,也有极少数为单核、三核或四核担孢子,不是灰花纹鹅膏菌正常发育的担孢子类型,它们的产生,可能是担子中的细胞核在向担孢子迁移的过程中出错所致。通过对分离培养的灰花纹鹅膏菌菌丝体的观察,未发现锁状联合之类的特征性结构,推测其产生的二核担孢子为同核孢子。但遗憾的是目前尚不能人工培养出灰花纹鹅膏菌子实体,不能对二核担孢子的交配类型进行验证。
荧光染料Hoechst 33258能特异性地与核DNA结合,因此排除了常规染色法如苏木精或吉姆萨(Giemsa)染液染色中对中心粒及核仁染色的干扰,重现性好,且通过荧光信号进行检测,可以观察到在细胞中保持自然状态的核或染色体,是研究真菌减数分裂等核行为及观察染色体数目的有效试剂。在灰花纹鹅膏菌菌丝体和担子中,有丝分裂和减数第一次分裂时期都能观察到染色体排列成环状,而且还观察到担子中第一次减数分裂形两个“上星环”后,第2次减数分裂随即发生,呈现一个担子中4个“土星环”的结构。通过荧光染色方法,观察到灰花纹鹅膏菌基因组染色体数目为10条。
