西湖风景名胜区森林群落结构特征与动态
2014-04-27张洋慎佳泓张方钢
张洋,慎佳泓,张方钢
(1.浙江自然博物馆,浙江杭州 310014;2.湖州市环境保护局,浙江湖州 313000)
西湖风景名胜区森林群落结构特征与动态
张洋1,慎佳泓2,张方钢1
(1.浙江自然博物馆,浙江杭州 310014;2.湖州市环境保护局,浙江湖州 313000)
以西湖风景名胜区森林植被为研究对象,在样方法野外调查的基础上,进行组平均聚类,划分出8个群落类型,分别为马尾松Pinusmassoniana林、马尾松-苦槠Castanopsis sclerophylla林、朴树Celtis sinensis-珊瑚朴C.julianae林、樟树Cinnamomum camphora-麻栎Quercus acutissima林、木荷Schima superba-苦槠林、青冈Cyclobalanopsis glauca林、毛竹Phyllostachys edulis林和毛竹-槲栎Quercus aliena林。用“空间序列代替时间变化”的方法,对优势种群的年龄结构、存活曲线进行研究。结果表明:马尾松-苦槠林、木荷-苦槠林、青冈林为增长型的种群,马尾松林属于衰退型种群,朴树-珊瑚朴林属于稳定型向衰退型过渡的类型。樟树-麻栎林为不稳定类型,有发展成为樟树纯林的可能。采用方差/均值比率法、负二项式法和平均拥挤指数等方法,研究优势种群的空间分布格局,种群随机分布的是马尾松林、马尾松-苦槠林、樟树-麻栎林和毛竹-槲栎林,集群分布的是朴树-珊瑚朴林、木荷-苦槠林、青冈林和毛竹林。图3表2参24
森林生态学;森林群落;大小级结构;存活曲线;空间分布格局;演替;西湖风景名胜区
结构特征是森林群落的重要属性之一。种群是构成群落的基本单位。种群的年龄结构是种群内不同年龄的个体数量的分布情况,不仅对群落结构有直接影响,更能很好地反映种群的动态变化[1]。种群分布格局是种群个体在植物群落中的空间分布[2]。它与环境进程和外力作用密切相关,如种间和种内竞争、种子散布、干扰、环境异质性等[3]。植物空间结构的分析是研究种群特征、种群和环境相关性的重要方法,有助于揭露群落演替的潜在进程和机制[3]。分析森林群落优势种群的年龄结构和分布格局,对于研究种群的更新方式和更新动态、群落演替趋势[4-6],揭示珍稀濒危植物种群衰退的外在原因[1,7-8],具有重要的意义。同时,通过阐明种群与环境的相互作用与揭示格局的成因,为保护措施的制定和恢复重建提供一定的科学依据。西湖风景名胜区是著名旅游胜地,其植被在20世纪40年代遭到严重破坏,后经几十年的封山育林,现已郁郁葱葱。20世纪80年代以来,该区植被的主要研究重点为次生植被性质、类型[9-11]。近年来,主要集中在物种多样性和人为干扰的影响[12-13],以及生态服务功能价值评估[14]。本研究对西湖风景名胜区的森林群落结构特征进行了研究,旨在分析该区典型优势种群的年龄结构、空间分布特点及其动态变化,环境及人为干扰对森林群落产生的影响,进而初步揭示西湖风景区森林植被的演替进程,并为制定合理有效的森林和生物多样性保护措施提供参考。
1 研究地概况
西湖风景名胜区的森林植被主要分布在西湖山区一带,整个山区总面积为40.21 km2,本研究中24个研究样地均设在此,约位于30°11′191″~30°16′N,120°04′743″~120°08′E。
西湖山区位于杭州城区西部,地处亚热带季风气候区,四季分明,夏季炎热多雨,冬季温暖湿润,年相对平均湿度80%左右,年平均降水量为1 399 mm。年平均气温16.2℃,7月平均气温28.6℃,最冷的1月平均气温3.7℃,≥10℃的平均积温为5 101.9℃,无霜期246 d,生长期长达311 d[15]。
西湖山区的地质构造属杨子淮台地东南边缘浙西一部分,以低山丘陵为主,由古生代沉积岩和中生代火山岩构成。土壤种类繁多,资源丰富,成土母质良好,有机质含量较高,酸碱适中,主要有红壤、石灰(岩)土、水稻土、淤灌土等[16]。
该区主要森林植被类型为常绿阔叶林。但每年冬天的短暂低温,不利于喜热常绿植物的生长,大量分布的常绿植物以较能耐寒的为主,于是就出现了许多落叶阔叶林或常绿与落叶阔叶混交林[15]。
2 研究方法
2.1 样地设置
根据卫星图片,结合对西湖风景名胜区的实地勘察,在该区选了14个研究地点,分别是北高峰、梅家坞、龙井、九溪、灵隐寺、灵隐茶园、虎跑、老和山、理安寺、云栖、飞来峰、外大桥、飞来峰、美人峰等地。在上述地点,选择典型地段设置20 m×20m样地,共24个样地。各样地分布概况见表1。24个样地的优势树种主要为马尾松Pinusmassoniana,苦槠Castanopsis sclerophylla,朴树Celtis sinensis,珊瑚朴C.julianae,樟树Cinnamomum camphora,麻栎Quercus acutissima,木荷Schima superba,青冈Cyclobalanopsis glauca,毛竹Phyllostachys edulis和槲栎Q.aliena。

表1 植被类型及所属样地概况Table 1 Vegetation typesand environmental data at 24 plots
2.2 数据采集
20 m×20 m的样地分成16个5 m×5 m的小样方进行调查。记录所在样地的群落生态学特征及周围生境,包括样地的群落高度、郁闭度、土壤状况、海拔、坡度、坡向、岩石裸露、干扰情况及乔木层、灌木层、草本层的高度和盖度。在5 m×5 m的小样方内,对胸径>3 cm或树高>5m的乔木进行每木调查,记录每个样方中乔木个体的种类、胸径、高度和冠幅;在每个5m×5m的小样方的右下角2 m×2 m处进行灌木的每木调查,记录灌木的种类、高度、盖度;草本则在右下角1m×1 m处进行调查,记录种类、高度、盖度。记录样方内的层间植物种类。此外,在整个5 m×5 m内记录其他植物种类。
2.3 数据处理
2.3.1 种群大小级划分由于立木种群的年龄结构在野外不易测定,通常采取空间代替时间的方法,即用立木的大小级来代替不同的年龄结构[2]。1级:H<0.33m;2级:H≥0.33m,DBH<2.5 cm;3级:2.5 cm≤DBH<7.5 cm;4级:7.5 cm≤DBH<22.5 cm;5级:DBH≥22.5 cm。
2.3.2 重要值重要值是反映植物种类在群落中地位重要程度的一个综合性指标,一般由各种类的相对多度、相对显著度和相对频度推算而得[17]:

首先将24个样方合并,由公式(1)计算立木层树种的重要值VI,选择在前40位的种作为聚类种。然后由式(2)计算40个种在各样方中的重要值,构成40×24矩阵,进行聚类。
2.3.3 聚类运用SPSS 16.0软件,选择Ward系统聚类法,得到聚类树状图。
2.3.4 种群分布格局的测定[18]①本研究采用泊松(Poisson)之方差/均值比率法V/m,并将比值与期望值之间的差异进行t检验。其中:V为样本方差,m为样方个体数均值;②负二项式(negative binomial parameter):K=m/(V/m-1)。若K>0,种群为集群分布,且随着K值的增大而集群程度降低;若K<0,则为随机分布。③平均拥挤指数(index o fmean crowing):M=m+(V/m-1)。聚块指数:C=M/ m。若C=1为随机分布,若C>1为聚集分布,若C<1为均匀分布。
3 结果和分析
3.1 群落分类及结构特征
根据聚类结果(图1)对24个样地进行分类,可分为8种群落类型,分别为马尾松林(样地1,样地2),马尾松-苦槠林(样地3,样地4,样地5),朴树-珊瑚朴林(样地6,样地7),樟树-麻栎林(样地8,样地9,样地10,样地11),木荷-苦槠林(样地12,样地13,样地14,样地15,样地16),青冈林(样地17,样地18,样地19,样地20),毛竹林(样地21,样地22,样地23)和毛竹-槲栎林(样地24)。
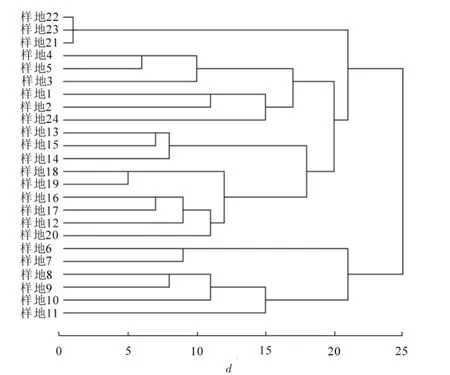
图1 群落聚类结果Figure 1 Clustering result for the communities
3.2种群格局分析
3.2.1 种群年龄结构年龄结构把每一年龄组个体的数量描述为1个年龄群对其他年龄群的比率。增长型种群为金字塔形,表示种群中有大量幼体,是迅速增长的种群。稳定型种群为圆锥形,种群数量稳定。衰退型种群为倒金字塔型,幼体少而老年个体比较大,数量趋于下降的种群[19]。除去人工经营的毛竹林和村庄附近的毛竹-槲栎林,将其他6个种群的大小级结构进行整理(图2)。从图2可以看出,马尾松-苦槠林、木荷-苦槠林、青冈林年龄结构接近金字塔型,林下有大量幼苗和幼树,可视为增长型的种群。马尾松-苦槠林下全部为苦槠幼苗,没有马尾松幼苗,可推测随着时间推移,马尾松会逐步被苦槠取代,发展成为苦槠纯林。马尾松林林下无马尾松幼苗,种群将无法天然更新,属于衰退型种群。朴树-珊瑚朴林为落叶阔叶林,幼苗和幼树比例较小,属于稳定型向衰退型过渡的类型。樟树-麻栎林为常绿落叶阔叶混交林,幼苗中麻栎占大多数,而3龄级中无麻栎,说明较大的郁闭度可能会影响落叶树种在林下的更新,从而发展成为樟树林。
3.2.2 存活曲线为了更进一步对各个优势种群年龄结构动态进行比较,最常见的方法是绘制存活曲线。存活曲线直观地反映着种群个体在特定年龄级的存活状况[20]。特定年龄或年龄组的出生率或死亡率不能确定时,可以根据存活曲线的类型来判断种群是增长、下降还是稳定的动态特征。如果存活曲线是直线型,表明种群为稳定型;若为凹形,则为增长型种群;若为凸型,则为衰退型种群[21]。以特定年龄级的存活个体数(n)的自然对数为纵坐标,以对应年龄级为横坐标,绘制存活曲线(图3)。马尾松-苦槠林、木荷-苦槠林线型近似凹形,属于增长型种群。青冈林的线型开始平缓且呈直线型,从1~4龄级的死亡率基本一致,种群较为稳定。朴树-珊瑚朴林和樟树-麻栎林线型相似,都经历一个“下降—上升—再下降”的波动过程,也佐证了其种群不稳定的格局特征。马尾松林林下无幼苗,线型不完整。

图2 优势种群年龄大小级结构Figure 2 Size structure of the dominant populations

图3 优势种群存活曲线Figure 3 Survival curves of the dominantpopulations
3.2.3 种群分布格局种群发育过程中,分布格局随时间推移而发生动态变化。本研究采用空间代替时间的方法,讨论各个优势种群的分布格局和动态,采用方差/均值比率法,以t检验确定显著度,并用负二项式(K),平均拥挤指数(M)和聚块指数(C)等其他方法辅助分析。结果见表2。从t检验的结果看,种群随机分布的是马尾松林、马尾-苦槠林、樟树-麻栎林和毛竹-槲栎林,集群分布的是朴树-珊瑚朴林、木荷-苦槠林、青冈林和毛竹林。从K值来看,马尾松林属于随机分布,其他属于集群分布,且K值越大的聚集程度越小,马尾松-苦槠林的K值达到33.27,可见集群程度很低,其次毛竹林、苦槠-木荷林的集群程度也较低。从C值来看,马尾松林属于平均分布,其他种群为集群分布。
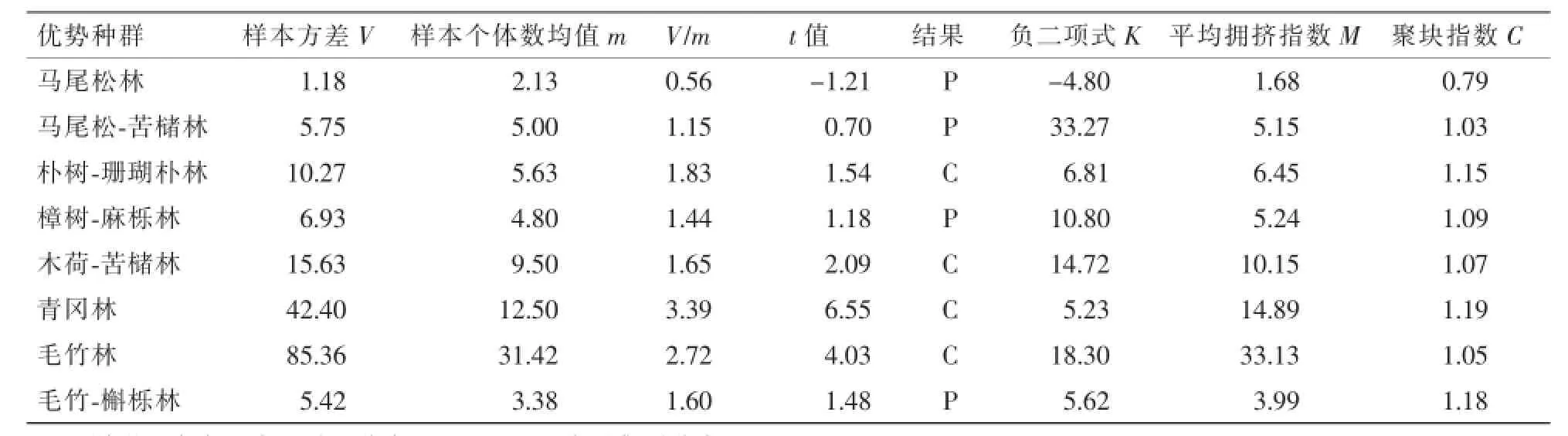
表2 西湖风景区优势种群的分布格局Table 2 Distribution pattern of the dominant populations in theWest Lake Scenic of Hangzhou
4 讨论与结论
年龄结构是种群的重要特征。许多学者都采用大小级结构和存活曲线来研究种群结构和动态,进而研究森林群落的演替[2]。①西湖风景名胜区的马尾松林林下几无幼苗与幼树,年龄结构不完整,这可能与其地理位置和人为干扰有关。马尾松林多位于山顶及山岗上部,松果可能随着山坡向下滚落,加上雨水的冲刷,种子难以停留在林下;同时,西湖风景区山顶多为登山爱好者的通道,人为干扰也会对种子的扩散与萌发造成影响。马尾松群落天然更新存在困难。②马尾松-苦槠林为针阔叶混交林,从大小级结构和存活曲线来看,为增长型种群。但其林下全部为苦槠幼苗和幼树,无马尾松小苗,可见随着马尾松大树的死亡,会逐渐发展成为常绿的苦槠林。③朴树-珊瑚朴林为落叶阔叶林,幼苗和幼树比例较小;群落郁闭度较大,会影响落叶树的种子在林下萌发与生长[2],一旦被耐阴的常绿树种入侵,则会慢慢被取代。④樟树-麻栎林为常绿落叶阔叶混交林,幼苗中麻栎占大多数,说明麻栎天然下种能力强,但是在3龄级中无麻栎,可能是较大的郁闭度影响落叶树种在林下的更新,或者资源有限产生自疏效应,导致小树无法继续生长发育成为中龄树,从而可能发展成为樟树林。⑤木荷-苦槠林和青冈林为常绿阔叶林,是亚热带地区最常见建群树种和优势树种,从年龄结构和存活曲线可以看出其在西湖风景区的种群较为稳定,且林下有大量幼苗幼树,是未来会持续增长的种群类型。
树种的生物学、生态学特征决定了其在不同演替阶段群落中的地位和作用是主导还是从属,在群落演替过程中是替代还是被替代,树种的种群结构类型正是其所处地位和作用以及发展过程和趋势的综合表现[22]。从上面可见,西湖风景区的先锋种马尾松林属于衰退种群,马尾松-苦槠林有发展为苦槠纯林的倾向,落叶树种群落不稳定,常绿阔叶林种群稳定且会持续增长。西湖风景区的森林群落有明显的次生特征,目前正处于针叶林和落叶阔叶林衰退向常绿阔叶林发展的演替过程中,且常绿阔叶林未达到最成熟稳定状态,在向顶级常绿阔叶林过渡。
林分环境的差异,种群生物学特性不同,以及林分范围上的资源分布不均,都是导致空间分布格局多样的原因[3]。森林中大小不一的林窗,林窗内光照充足,有利于强阳性的先锋种快速生长,从而集群分布在林分中。随着林木的生长,对光照、养分、水分、生长空间的需求增大,自然稀疏效应产生,从而导致种群分布格局由集群趋向于随机[23-24]。西湖风景名胜区的优势种群中,呈现随机分布的马尾松林、马尾松-苦槠林就属于上述情况。木荷-苦槠林、青冈林的幼苗幼树比例较高,植物的种子因重力会落在母体植物附近,当种子萌发就会产生一簇幼小的植物,幼苗、幼树一般成斑块状聚集[22]。毛竹通过根茎从母株蔓延开去,也会形成簇生的个体,呈现聚集分布。
简敏菲等[18]研究认为:种群在散布和发展时期的分布格局表现为集群分布,而在衰退期常表现为随机分布。西湖名胜区优势种群的分布格局符合此理论,常绿的木荷-苦槠林和青冈林属于增长型年龄结构,集群程度高,种群繁衍后代较为容易。马尾松林、马尾松-苦槠林、樟树-麻栎林和毛竹-槲栎林,属于衰退和不稳定的种群,其空间格局呈现随机分布趋势。
本研究采用聚类方法得出优势种群,方法相对简单、有效。研究种群年龄结构和空间分布,分析西湖风景区森林群落的现状及演替趋势,可针对具体情况制定相应的生物多样性保护措施。该区属于城市森林范畴,山上公路、游步道、茶园等都会对森林群落有人为干扰影响[12]。如何加强森林保护,减少人为干扰,是需要思考的。本研究受调查样地数量的影响,不能涵盖所有森林群落类型,群落特征难以描述详尽。各个优势种群具体的分布格局动态,也会因地理位置和海拔高度的不同而发生变化,这些还有待深入研究的。
[1]杨宁,邹冬生,杨满元.贵州雷公山秃杉的种群结构和空间分布格局[J].西北植物学报,2011,31(10):2100-2105.
YANG Ning,ZOU Dongsheng,YANGManyuan.Structure and spatial distribution pattern ofTaiwania flousianapopulation in Leigong Mountain,Guizhou[J].Acta Bot Boreal-Occident Sin,2011,31(10):2100-2105.
[2]孙澜,苏智先,张素兰.嘉陵江流域南充金城山森林群落结构特征分析[J].植物研究,2008,28(3):364-369.
SUN Lan,SU Zhixian,ZHANG Sulan.Forest community structural features of Jincheng Mountain in Nanchong Region in Jialing River Basin[J].Bull Bot Res,2008,28(3):364-369.
[3]杜志,亢新刚,孟京辉,等.长白山杨桦次生林主要树种的空间分布格局及其关联性[J].东北林业大学学报,2013,41(4):36-42.
DU Zhi,KANG Xingang,MENG Jinghui,et al.Spatial distribution patterns and associations of dominant tree species inPoplar-Birchsecondary forest standin ChangbaiMountains[J].JNortheast For Univ,2013,41(4):36-42.
[4]范繁荣,潘标志,马祥庆.白桂木的种群结构和空间分布格局研究[J].林业科学研究,2008,21(2):176-181.
FAN Fanrong,PAN Biaozhi,MA Xiangqing.A study on structure and spatial distribution pattern ofArtocarpus hypargyreuspopulation[J].For Res,2008,21(2):176-181.
[5]郭连金,徐卫红.武夷山米槠种群结构及谱分析[J].植物研究,2007,27(3):325-330. GUO Lianjin,XU Weihong.Structure and spectral analysis ofCastanopsis carlesiipopulation in Wuyishan Nature Reserve[J].Bull Bot Res,2007,27(3):325-330.
[6]ARISTAM.The structure and dynamicsofan Abies pinsapo forest in southern Spain[J].For EcolManage,1995,74:81-89.
[7]李辛雷,孙振元,李纪元,等.广东鹅凰嶂自然保护区杜鹃红山茶种群结构与空间分布格局[J].应用生态学报,2013,24(8):2115-2121.
LIXinlei,SUN Zhenyuan,LI Jiyuan,et al.Population structure and spatial distribution pattern ofCamellia azaleain E’huangzhang Nature Reserve of Guangdong,China[J].Chin JAppl Ecol,2013,24(8):2115-2121
[8]彭焱松,张晓波,桂忠明,等.庐山香果树毛竹混交林空间格局研究[J].广西植物,2013,33(4):502-507.
PENG Yansong,ZHANG Xiaobo,GUIZhongming,et al.Spatial distribution pattern inEmmenopterys henryiandPhyllostachys edulismixed forest in Lushan Mountain[J].Guihaia,2013,33(4):502-507.
[9]陈启瑺.杭州西湖山区次生植被性质研究(一)[J].杭州大学学报,1987,14(4):473-481.
CHEN Qichang.Characteristics of vegetation on hills and lower mountains in vicinity of West Lake in Hangzhou(Ⅰ)[J].JHangzhou Univ Nat Sci,1987,14(4):473-481.
[10]陈启瑺.杭州西湖山区此生植被性质研究(二)[J].杭州大学学报,1988,15(1):81-87.
CHENQichang.Characteristics of vegetation on hills and lowermountains in vicinity ofWest Lake in Hangzhou(Ⅱ)[J].JHangzhou Univ Nat Sci,1988,15(1):81-87.
[11]陈启瑺.杭州西湖山区植被的分类[J].植物生态学与地植物学学报,1988,12(4):265-271.
CHEN Qingchang.Classification of vegetation on hills and lowermountainous in vicinity ofWest Lake in Hangzhou[J].Acta PhytoecolGeobat Sin,1988,12(4):265-271.
[12]张洋,慎佳泓,张方钢,等.西湖风景名胜区森林群落物种多样性及人为干扰的影响[J].浙江大学学报:理学版,2008,35(5):567-575.
ZHANG Yang,SHEN Jiahong,ZHANG Fanggang,et al.Forest community species diversity and influence of hu-man disturbance in mountainous area of the West Lake in Hangzhou,East China[J].J Zhejiang Univ Sci Ed,2008,35(5):567-575.
[13]陈攀,慎佳泓,胡广,等.西湖风景名胜区不同类型森林群落的空间分布及β多样性[J].生态学报,2009,29(6):2929-2937.
CHEN Pan,SHEN Jiahong,HU Guang,et al.Spatial distribution and beta diversity of forest communities in mountainous areas of theWest Lake in Hangzhou,East China[J].Acta Ecol Sin,2009,29(6):2929-2937.
[14]唐宇力,任笑一,范丽坤,等.杭州西湖风景名胜区山林生态服务功能价值评估[J].浙江林业科技,2012,32(5):54-59.
TANG Yuli,REN Xiaoyi,FAN Likun,et al.Assessment of ecosystem service of the West Lake Scenic Area in Hangzhou[J].JZhejiang For Sci Technol,2012,32(5):54-59.
[15]蔡壬侯,何绍箕.杭州西湖山区的植被类型及其分布[J].杭州大学学报,1980,7(4):100-107.
CAI Renhou,HE Shaoqi.Vegetation types and its distributing in the mountainous areas of the West Lake in Hangzhou[J].JHangzhou Univ,1980,7(4):100-107.
[16]刘经雨.杭州西郊的土壤及其分布规律[J].杭州大学学报,1978,5(1):86-104.
LIU Jingyu.Soil and its distributing in the west Hangzhou[J].JHangzhou Univ,1978,5(1):86-104.
[17]宋永昌.植被生态学[M].上海:华东师范大学出版社,2005:90-91,128-131.
[18]简敏菲,刘琪璟,朱笃.江西九连山常绿阔叶林优势种群的年龄结构和分布格局[J].广西植物,2010,30(3):348-354.
JIAN Minfei,LIU Jingqi,ZHU Du.Age structure and distribution pattern of the dominant population in the evergreen broad-leaved forest of Jiulian Mountain in Jiangxi Province[J].Guihaia,2010,30(3):348-354.
[19]DEEVEY E SJr.Life tables for natural populations of animals[J].JVert Paleontol,1947,22(4):283-314.
[20]HETT JM,LOUCKSO L.Age structure models of balsam fir and eastern hem lock[J].JEcol,1976,64(3):1029-1044.
[21]LEAKW B.Age distribution in virgin red spruceand northern hardwords[J].Ecology,1975,56(6):H1451-1454.
[22]熊能,金则新,陈琢,等.千岛湖姥山马尾松种群结构和分布格局研究[J].植物研究,2010,30(5):537-542.
XIONG Neng,JIN Zexin,CHEN Zhuo,et al.Population structure and distribution pattern ofPinusmassonianaon Laoshan Island of Qiandao Lake Region[J].Bull Bot Res,2010,30(5):537-542.
[23]缪宁,刘世荣,史作民,等.川西亚高山红桦-岷江冷杉林优势种群的空间格局分析[J].应用生态学报,2009,20(6):1263-1270.
LIAONing,LIU Shirong,SHIZuomin,et al.Spatial patterns of dominant tree species in sub-alpine Betula-Abies forest in west Sichuan of China[J].Chin JAppl Ecol,2009,20(6):1263-1270.
[24]闫海冰,韩有志,杨秀清,等.华北山地典型天然次生林群落的树种空间分布格局及其关联性[J].生态学报,2010,30(9):2311-2321.
YAN Haibing,HAN Youzhi,YANG Xiuqing,et al.Spatial distribution patterns and associations of tree species in typical natural secondary mountain forest communities of Northern China[J].Acta Ecol Sin,2010,30(9):2311 -2321.
Structure and dynamics of forest community in the West Lake Scenic of Hangzhou,China
ZHANG Yang1,SHEN Jiahong2,ZHANG Fanggang1
(1.Zhejiang Museum of Natural History,Hangzhou 310014,Zhejiang,China;2.Huzhou Environmental Protection Bureau,Huzhou 313000,Zhejiang,China)
Structure is one of themost important characteristics of a forest community,and population is its basic unit.The study on the age structure and the spatial distribution has importantmeanings to the research of population dynamics and community succession.This study was designed to help clarify vegetative ecological characteristics and change rules as well as to provide reference for establishing plans to protect the forest and the biodiversity in this area.Using quadrate investigation,the forest community in the West Lake Scenic of Hangzhou was divided into eight forest types including Pinusmassoniana(A),P.massoniana-Castanopsis sclerophylla(B),Celtis sinensis-C.julianae(C),Cinnamomum camphora-Quercus acutissima(D),Schima superba-C.sclerophylla(E),Cyclobalanopsis glauca(F),Phyllostachys edulis(G)andPh.edulis-Q.alien a(H),and analyzed by group average clustering to confirm dominant populations.The size stuctrue and survival curves of the dominant populations were studied with a spatial sere substituted for a temporal sere.Also,the spatial distibution pattern of themain dominant populationswas determined using a Poisson distrbution,a negative binomial parameter,and amean crowing index.The results of the size structure and survival curves showed that B, E and Fwere growing populations,A was a declining pupulation,C was a transitional type going from stable todeclining,and D which was not stable could transform into a pureCinnamomum camphoraforest.The spatial distribution patterns showed that A,B,D,and H were random distributions;C,E,F,and G were clumped distributions.Forest vegetation was in successional change with the declining coniferous forest and deciduous broad-leaf forest developing into an evergreen broad-leaf forest and the evergreen broad-leaf forest transforming into a climax forest.Group average clustering was relatively simple and effective for confirming dominant populations.This study could help analyze the present situation and successional trends of the forest community in theWest Lake Scenic of Hangzhouto protectbiodiversity.[Ch,3 fig.2 tab.24 ref.]
forest ecology;forest community;size structure;survival curves;spatial distribution pattern;succession;West Lake Scenic of Hangzhou
Q948;S718.5
A
2095-0756(2014)04-0589-08
2013-11-04;
2014-01-06
浙江省科技计划项目(2007C22084);杭州市科技发展计划项目(2004113B05)
张洋,馆员,从事植被生态学和保护生物学研究。E-mail:zhangyang1905755@163.com。通信作者:张方钢,研究馆员,从事植物分类学与生态学研究。E-mail:zhangfg-88@163.com
