Dependences of longleaf pine (Pinus palustris) natural reproduction on environments above ground
2014-04-20QiujuGuoDexiangWangXunruAiLanYao
Qiuju Guo · Dexiang Wang · Xunru Ai · Lan Yao
Introduction
With decline in acres of longleaf pine (Pinus palustris) across its range, longleaf pine became a focal point for the Center in the late 1980’s and early 1990’s. Longleaf pine timber stands have been maintained by the use of prescribed fire, the removal of other species during timber harvest. Today, many longleaf stands exist (both natural and planted) that are maintained with regular prescribed fire to produce an open, lush understory of native plants and natural regeneration.
Natural regeneration of longleaf pine is often characterized by limited seed production and low seedling survival. Longleaf pine is known as a poor and periodic seed producer (Landers et al 1995; Boyer 1998). Seeds of high quality are produced every 5-7 years (Hardin et al. 2001). Seedlings develop among grasses for the first 3-25 years. During this period, seedlings concentrate on root growth, with a slow rate of stem growth. The delayed in stem growth allows more space for competition from other fastgrowing species in the absence of wildfire or prescribed fire. Seedlings are susceptible to defoliation by brown-spot needle blight infection. Researchers have attempted to overcome these difficulties to encourage growth of more natural seedlings by studying how overstory and understory environments influence seedling survival and growth (Pecot et al. 2007; Knapp et al. 2008). Some evidence showed that longleaf pine seedlings were negatively affected, both directly and indirectly, by competition from overstory and understory plants (Kush and Meldahl 2004; Dyson 2010). The overstory of longleaf pine facilitated seedling survival within a range of basal areas but natural seedling growth declined as overstory stocking increased. The overstory of longleaf pines also influences the growth of seedlings through competition for light and nitrogen (Brockway and Outcalt 2000; McGuire et al 2001; Battaglia et al. 2002, 2003).
When undisturbed by fire, understory reduces the growth of seedlings through root competition (Rodriguez-Trejo et al. 2003; Brockway et al. 2005a). Root competition is an important factor causing slow growth of longleaf pine seedlings. Sufficient available nutrients can be provided for the growth of longleaf pine seedlings (Mulligan and Kirkman 2002) by removal of grasses and weeds (Addington et al. 2006).
Brockway et al. (2005b) suggested that natural regeneration of longleaf pine can be achieved through selection of forest structure (such as forest type and density) and spatial pattern. Variable retention silvicultural is an effective approach for rapidly restoring microclimate factors associated with structurally complex forests (Brockway et al. 2006). Better understanding of the influences of overstory retention and understory environments on seedling regeneration will increase the ability to successfully manage and restore a forest (Brockway et al. 2005c).
Although selection silvicultural techniques have been successfully used in this ecosystem, the relationships between natural seedling reproduction and overstory or understory characters (canopy, stand density, understory competition and light environment) sustained by the prescribed fire every 2 or 3 years are still not clear. This work focused on the influences of above ground environmental factors (overstory and understory) on natural regeneration of longleaf pine seedlings in the selected and control stands in three main distribution areas in south Alabama, United States. Six above-ground indices were used in this project and three methods of regression analysis were used to quantify relations between natural seedling reproduction and important environmental factors. Results are expected to provide references for future work on longleaf pine reproduction.
Methods
Study sites
This project was conducted at three study sites: Blackwater River State Forest (Alabama part), Escambia Experimental Forest, and Auburn University’s Solon Dixon Forestry Education Center. The prescribed fires for increased longleaf pine reproduction are habitually performed during November through April at Blackwater River State Forest and Escambia Experimental Forest sites, and 5 years’ absence of prescribed fire in Dixon Center in 2010.
The Blackwater River State Forest, about 2096 ha, is located at Santa Rosa County (30.8°N, 86.8°W), in the Southern Pine Hills region of northwest Florida, United States, in the Alabama Area longleaf pine site zone (Craul et al. 2005). Topography is characterized as gently sloping to sloping, with elevations between 60-75 m. Average annual temperature is 19.4°C, and rainfall averages 1650 mm per year. The average growing season is 300 days. The primary soil phase on the study site is well drained soils and loamy subsoil. The overstory on the Blackwater forest was dominated by longleaf pine developed from natural seeding after the original forest was cut in the 1920s. Some oak species such as southern red oak (Quercus falcata Michx.), turkey oak (Q. laevis Walt.), and sand post oak (Q. margaretta Ashe) were present in the midstory and overstory. The understory was largely composed of wiregrass (Aristida spp.) and associated species; woody plants including sassafras (Sassafras albidum Nutt.), common persimmon (Diospyros virginiana L.), blueberry (Vaccinium spp.) and various clonal oaks (Quercus spp.) were also common. In order to promote tree growth, thinning cutting was proceeded from winter 1990 to spring 1991 in Blackwater River State Forest. The single-tree, group-tree selection cutting and non-cutting were operated in autumn 2006 (Jones et al. 2003).
The Dixon Center includes 53 ha of managed pine upland and hardwood bottomlands and is located in Covington and Escambia counties in the Gulf Coastal Plain of south-central Alabama (31.1°N, 86.8°W). Soils of the region are deep and well drained with sandy loam surface soils over sandy clay loam or loam. Topography of the region is highly and varies from deeply dissected to gently rolling hills. Elevation ranges from 20 m to 137 m a.s.l. The climate is mild with long summers and cool short winters. Precipitation is evenly distributed throughout the year with rare prolonged droughts. Average annual rainfall is 150 cm with 53% occurring between April and September. The overstory was dominated by longleaf pine, the dense shrub understory was dominated by yaupon (Ilex vomitoria) along with lesser abundeance of blueberries and gallberry (I. glabra). Selective cutting of different models was carried out during fall 2004.
Escambia Experimental Forest (EEF) is located in southcentral Escambia County, Alabama (31.1°N, 87.1°W). It occupies 30 ha in the central location in the longleaf pine belt. Soils are generally favorable to tree growth, sandy and well-drained, but with adequate moisture (Kush et al. 2000). Topography is flat to rolling; most slopes are between 3° and 10°. Elevation varies from 31 m to 83 m a.s.l. The climate is humid and mild with plentiful rainfall well distributed throughout the year. Annual precipitation is 1560 mm well distributed throughout the year. The average annual range of temperatures is from -6°C to 37°C. The growing season is 250 days (Kush et al. 1999). Ninety percent of overstory trees were longleaf pines. Yaupon, blueberries, inkberries (Ilex glabra) and brackenferns (Pteridium aquilinum) dominated the understory. The clear cutting was carried out in 1948, and the woodland was basically dominated by natural longleaf pine mixed with some oaks in 1966. In 1986, another harvest cutting was conducted at the study site. Research selective cutting was made in total area of 30 ha in spring 2006.
Plot description
In summer 2010, 48 sample plots were randomly demarcated at three study sites, 27 at Escambia, 12 at Blackwater and 9 at Dixon center (Table 1). Each circular plot was 0.05 ha (12.62 m radius). To ensure that all plots were initially under a closed forest canopy, plots were established no less than 12.6 m from adjacent plots or from natural forest gaps (0.025 ha). Four subplots (0.001 ha, 1.78 m radius) in four cardinal directions were demarcated in each plot, each subplot center was the middle point of the main plot radius (Fig. 1).

Fig. 1: Plot and subplots setting

Table 1: Plots description
Data collection and analysis
Plot Overstory tree inventories were completed for each 0.05 ha plot. All trees >5 cm DBH were measured: height (m), DBH (cm), and species were recorded. Tree inventory data was summarized as average DBH (cm), stand density (number of trees ·ha-1) and basal area (m2·ha-1) for each plot.
Subplot (1) Understory composition (species and coverage) was recorded for each subplot. The plant layers were recorded as tree regeneration, shrub and grass layers. Coverage was calculated by view estimation in each subplot. (2) Numbers of longleaf pine seedlings in each subplot were tallied by two classes (grass stage and bolting stage classified by RCD of 15 mm). Five seedlings were randomly selected in each class for the subplots having five or more seedlings; for those having less than five seedlings, all were selected. The root collar diameter (RCD) of tallied seedlings was recorded. (3) Canopy closure was estimated using a convex spherical densiometer (Lemon 1956). Readings were taken directly over plot centers, subplot centers and edge of the four cardinal directions, and average canopy closure was used (Buckley et al. 1999). A previous study suggested that different observers could introduce bias into densiometer readings (Vales and Bunnell 1988), thus, a single individual collected all the densiometer data in the study. (4) Light intensity, measured as photosynthetically active radiation (PAR), was directly quantified by an AccuPar linear PAR/LAI ceptometer (Decagon Devices, Inc., Pullman, WA, USA). Nine readings were taken in each plot (the same reading points with the densiometer) at a height of 1.25 m above the ground. Every reading was an average of 80 individual sensors along the 0.8 m long ceptometer array. Care was taken to ensure that the ceptometer was leveled and pointed under the brightest light source to avoid shadow casting on the sensors from the observer.
Statistical analyses were conducted using SAS version 9.1 software (SAS Institute, Cary, NC, USA). For all tests the statistical significance level was α=0.05. Regression analyses were used to relate above ground indices and seedling reproduction numbers.
Results
Regeneration on three sites
The plots were not evenly set in the three study sites (Table 1) because each was part of a different longleaf pine distribution area. The Dixon center had 9 plots and the average DBH of longleaf pine was 21.12 cm, seedlings numbered 278 per ha and the average seedling RCD was the smallest of all sites (6.23 mm).
Twelve plots were sampled at Blackwater, where the basal area was greater than at Dixon center, but lower than at Escambia. Seedlings had the largest RCD (15.20 mm) at Blackwater. Escambia had 27 fixed plots and the overstory longleaf pine here had the greatest mean DBH (27.44 cm), and also the highest number of seedlings (38230/ha).
Above ground indices
Six basic above ground indices, viz. basal area, stand density, average height, canopy closure, PAR and understory coverage, were measured (Table 2). Escambia had the greatest basal area but the lowest stand density of 219 trees per hectare. The average height of longleaf pine was 19.86 m. Canopy closure at Escambia was as low (43.6%) as was that at Blackwater (43.6%). PAR at Escambia (546 μmol·m-2·s-1) was the highest of the three study sites. Understory coverage at Escambia was 15.3% lower than that at Dixon center and 7.8% higher than at Blackwater. At Dixon center, stand density was highest (293 trees per hectare) and canopy closure and understory coverage were also the highest of the three sites.
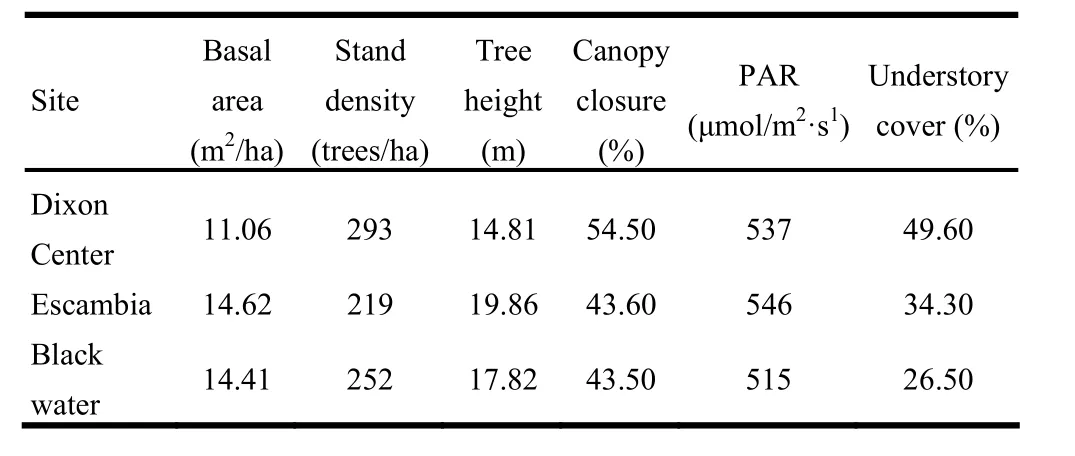
Table 2: Six stand indices at three study sites
Regression analysis
Backward regression analysis
Backward analysis starts with all candidate variables and tests each of them one by one for statistical significance, and deletes any variable that is not statistically significant at α=0.1. Three indices (canopy closure, stand density and basal area) remained after the above procedure (Table 3) and were used in building the regression model of relations between above ground indices and seedling number. The model was expressed as:

where SN is seedling number, CC is the canopy closure, SD is stand density, BA is the basal area
This regression yielded R-square of 0.2818 despite a P value<0.001, indicating certain extent of validity.
Forward regression analysis
Forward regression selection starts with no variables in the model and selects variables one by one from those satisfying statistically significance.

Table 3: Backward analysis for six above ground indices
The p value of the forward regression model was <0.01 (0.0018). Although the R-square value was not high, the p value proved this model was statistically significant (Table 4). Three indices: canopy closure, stand density and basal area were included in this model as no other variables met the 0.10 significance level for entering the model. The model can be expressed as:

where SN -- Seedling Number, CC -- Canopy Closure, SD --Stand Density, BA -- Basal Area
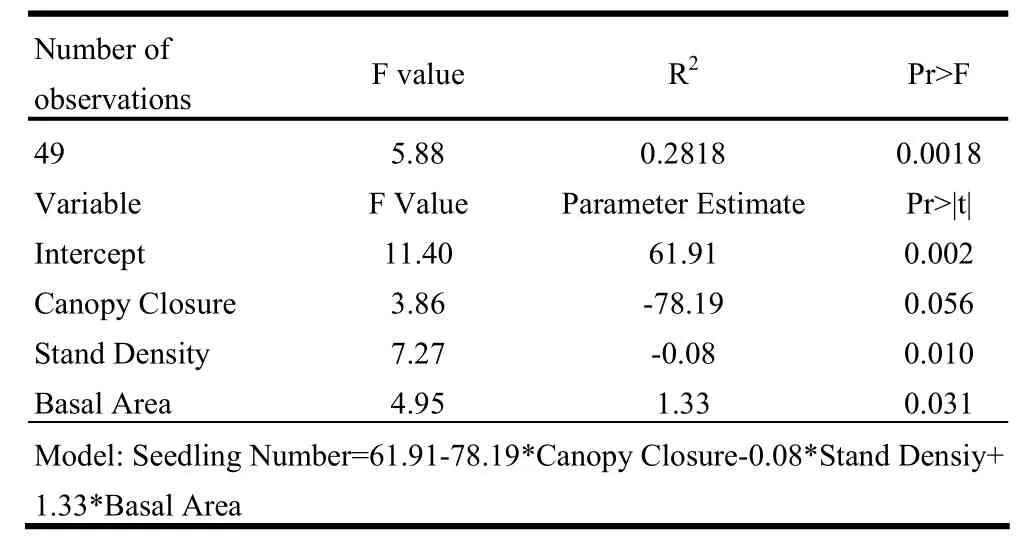
Table 4: Forward regression analysis for six above ground indices
Stepwise regression analysis
Stepwise regression is commonly used for developing statistical models when there is a large number of potential explanatory variables and no underlying theory on which to base the model selection. The procedure is used primarily in regression analysis, and the basic approach is applicable to many forms of model selection. At each stage of analysis, after a new variable is added, a test is made to check if some variables can be deleted without appreciably increasing the residual sum of squares (RSS). The procedure terminates when the measure is maximized, or when the available improvement falls below a critical value.
Three variables: canopy coverage, stand density and basal area were finally selected from the six above-ground indices in this model (Table 5). The model had similar p and R-square values to those of the backward and forward regression analysis models. The other three above ground variables: average tree height, PAR and understory coverage, were excluded from the model.

Table 5: Stepwise regression analysis for six above ground indices
Discussion
Differences of longleaf pine seedling regeneration in study sites
Longleaf pine seedlings need open spaces to regenerate. The collected data showed overstory and understory parameters were differed at the three study sites. The highest stand density, canopy closure and understory coverage at Dixon center explains the lowest abundance of seedlings here. Escambia is a historical longleaf pine growing area. The forest system here is the typical second growth forest. Silvicultural techniques were applied at Escambia in a timely and systematic program, and this enabled appropriate levels of the basal area and stand density at this site. Prescribed fire was used once every 2-3 years at Escambia to reduce understory cover. Thus, seedlings were most numerous at this site.
The lowest understory coverage and the largest average root collar diameter of seedlings were recorded at Blackwater. The lowest understory coverage was mostly because of the occasional tornados, but this needs to be confirmed by more studies. Understory coverage is an important index of sun light interception. At lower the understory coverage, more sun light is available to seedlings and their growth will be faster. This may explains the biggest root collar of natural seedling at this site among the three sites. The soil chemistry, water availability, or other factors which could produce some part of the growth, so it should be clarified in the future work.
The regression model of longleaf pine seedling reproduction
All six above-ground indices from 48 plots were analyzed to produce a regression model of longleaf pine natural seedling reproduction. Three modeling approaches (backward, forward and stepwise) of index selections in regression analysis were used in this work, and these approaches yielded the same three indices as primary determinants of seedling number: canopy closure, basal area and stand density. The other three indices did not meet the 0.10 significance level required by the model. The model can be expressed as:
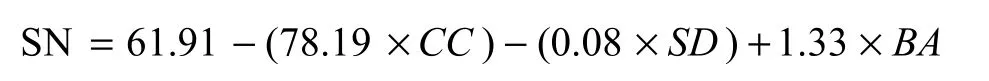
where SN -- Seedling Number, CC -- Canopy Closure, SD --Stand Densiy, BA -- Basal Area
From the above equation it can be seen that the number of seedlings declined with increasing canopy closure and increasing stand density, but increased with increasing overstory basal area. The F and R2values of this regression model suggest modestly significant impact of the above-ground variables on seeding reproduction, but the p value of this model implies, to a certain extent, that the model was scientifically meaningful. Thus, the model might be applied to estimate the number of longleaf pine natural seedling reproduction when some basic stand indices are available.
The model suggests that control of stand density and canopy closure will stimulate longleaf pine seedling reproduction if the basal area meets the management objective. Canopy closure and stand density are related variables: canopy closure increases with increasing stand density and the combined increase reduces the open space and root gap available for seedling reproduction. The R2value suggests there are some other important factors influencing the seedling reproduction, it needs to be explored in the next step.
Escambia had the highest number of seedlings. This resulted mainly from the selection methods: single-tree and group-tree selection. The two selection methods log some trees and open some extra space for seedling reproduction and growth. Additionally, prescribed fire is set every 2-3 years at Escambia, which eliminates some understory plants, and are able to provide more nutrients and decrease infections for longleaf pine seedlings. This indicates that selection techniques and prescribed fire should be considered to increase seedling numbers in longleaf pine ecosystems such as Dixon center.
Conclusion
Three above-ground indices, viz. canopy closure, basal area and stand density, were used to predict longleaf pine seedling numbers. Basal area was positively correlated, and canopy closure and stand density were negatively correlated with seedling numbers. The selection methods and prescribed fire improved the natural longleaf pine seedling reproduction in Escambia and Blackwater.
Acknowledgements
Thanks to USDA for providing the funding to our research. Many thanks are due to the school of forestry and wildlife science at Auburn University, and especially Dr. Edward F. Loewenstein for his contributions. Thanks also to John Kush, Ferhat Kara, David Dyson and James Ransom for their advice and kind help in the field work.
Addington RN, Donovan LA, Mitchell RJ, Vose JM, Pecot SD, Jack SB, Hacke UG, Sperry JS, Oren R. 2006. Adjustments in hydraulic architecture of Pinus palustris maintain similar stomatal conductance in xeric and mesic habitats. Plant Cell Environment, 29(4): 535-545.
Battaglia MA, Mitchell RJ, Mou PP, Pecot SD. 2003. Light transmittance estimates in a longleaf pine woodland. Forest Science, 49(5): 752-762.
Battaglia MA, Mou P, Palik B, Mitchell RJ. 2002. The effect of spatially variable overstory on the understory light environment of an opencanopied longleaf pine forest. Canadian Journal of Forest Research, 32: 1984-1991.
Boyer WD. 1998. Longleaf pine regeneration and management: An overstory overview. In: Kush JS (Comp.), Proceedings of the longleaf pine ecosystem restoration symposium, presented at the Society of Ecological Restoration 9th Annual International Conference. Longleaf Alliance Report No. 3, pp: 14-19.
Brockway DG, Outcalt KW, Boyer WD. 2006. Longleaf pine regeneration ecology and methods. In: Jose S, Jokela EJ, Miller DL (eds.), The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration. New York, Springer, pp: 95-133.
Brockway DG, Outcalt KW, Guldin JM, Boyer WD, Walker JL, Rudolph DC, Rummer RB, Barnett JP, Jose S, Nowak J. 2005b. Uneven-aged management of longleaf pine forests: a scientist and manager dialogue. USDA Forest Service Southern Research Station Gen. Tech. Rep. SRS-78. Asheville, NC, 38 pp.
Brockway DG, Outcalt KW, Tomczak DJ, Johnson EE. 2005a. Restoring longleaf pine forest ecosystems in the southern U.S. In: Stanturf JA, Madsen P (eds.), Restoration of Boreal and Temperate Forests. CRC Press, Boca Raton, FL, pp: 501-519.
Brockway DG, Outcalt KW, Tomczak DJ, Johnson EE. 2005c. Restoration of longleaf pine ecosystems. USDA Forest Service Southern Research Station General Technique Reprt SRS-83. Asheville, NC, 34 pp.
Brockway DG, Outcalt KW. 2000. Restoring longleaf pine wiregrass ecosystems: Hexazinone application enhances effects of prescribed fire. Forest Ecology and Management, 137: 121-138.
Buckley DS, Isebrands JG, Sharik TL. 1999. Practical field methods of estimating canopy cover, PAR, and LAI in Michigan oak and pine stands. Northern Journal of Applied Forestry, 16: 25-32.
Craul PJ, Kush JS, Boyer WD. 2005. Longleaf pine site zones. General Technical Report SRS-89, U.S. Department of Agriculture, Forest Service, Southern Research Station, Asheville, N.C. 14pp.
Dyson DS. 2010. Influence of Competition on Longleaf Pine Seedling Recruitment in Selection Silviculture. Ph. D Thesis: Auburn University, 148pp.
Hardin JW, Leopold DJ, White FM. 2001. Harlow and Harrar’s textbook of dendrology, 9th edition. McGraw-Hill, New York, pp: 142-146.
Jones RH, Mitchell RJ, Stevens GN, Pecot SD. 2003. No burns of fine root dynamics across a gradient of gap sizes in a pine woodland. Oecologia, 134: 132-143.
Knapp BO, Wang GG, Walker JL. 2008. Relating the survival and growth of planted longleaf pine seedlings to microsite conditions altered by site preparation treatments. Forest Ecology and Management, 255: 3768-3777.
Kush JS, Meldahl RS, Avery C. 2004. A restoration success: longleaf pine seedlings established in a fire-suppressed, old-growth stand. Ecological Restoration, 22(1): 6-10.
Kush JS, Meldahl RS, Boyer WD. 1999. Understory plant community response after 23 years of hardwood control treatments in natural long leaf pine (Pinus palustris) forests. Canadian Journal of Forest Research, 129(7): 1047-1054.
Kush JS, Meldahl RS, Boyer WD. 2000. Understory plant community response to season of burn in natural longleaf pine forests. Tall timbers fire ecology conference. Allen Press, Tallahassee, FL.pp:32-39.
Landers JL, Van Lear DH, Boyer WD. 1995. The longleaf pine forests of the Southeast: Requiem of renaissance. Journal of Forestry, 93(11): 39-44.
Lemmon PE. 1956. A spherical densiometer for estimating forest overstory density. Forest Science, 2: 314-320.
McGuire JP, Mitchell RJ, Moser EB, Pecot SD, Gjerstad DH, Hedman CW. 2001. Gaps in a gappy forest: plant resources, longleaf pine regeneration, and understory response to tree removal in longleaf pine savannas. Canadian Journal of Forest Research, 31: 765-778.
Pecot SD, Mitchell RJ, Palik BJ, Moser EB, Hiers JK. 2007. Competitive responses of seedlings and understory plants in longleaf pine woodlands: separating canopy influences above and below ground. Canadian Journal of Forest Research, 37: 634-648.
Rodriguez-Trejo DA, Duryea ML, White TL, English JR, McGuire J. 2003. Artificially regenerating longleaf pine in canopy gaps: initial survival and growth during a year of drought. Forest Ecology and Management, 180: 25-36.
Vales DJ, Bunnell FL. 1988. Comparison of methods for estimating forest overstory cover. I. Observer effects. Canadian Journal of Forest Research, 18: 606-609.
杂志排行

Journal of Forestry Research的其它文章
- The effects of understory vegetation on P availability in Pinus radiata forest stands: A review
- Phytosociology, structure and dynamics of Pinus roxburghii associations from Northern Pakistan
- Characterization of expressed genes in the establishment of arbuscular mycorrhiza between Amorpha fruticosa and Glomus mosseae
- Changes induced by osmotic stress in the morphology, biochemistry, physiology, anatomy and stomatal parameters of almond species (Prunus L. spp.) grown in vitro
- Effects of gaps on regeneration of woody plants: a meta-analysis
- A modified Murashige and Skoog media for efficient multipleshoot induction in G. arborea Roxb.