象山港游泳动物群落功能群组成与功能群多样性*
2014-04-16姜亚洲袁兴伟焦海峰李圣法
姜亚洲 林 楠 袁兴伟 焦海峰 李圣法
象山港游泳动物群落功能群组成与功能群多样性*
姜亚洲1林 楠1袁兴伟1焦海峰2李圣法1①
(1. 中国水产科学研究院东海水产研究所 上海 200090; 2. 宁波市海洋与渔业研究院 宁波 315012)
依据2011年4月、7月和11月象山港水域桁杆拖网和单拖网渔业资源定点调查资料, 对象山港水域游泳动物群落的功能群组成及功能群多样性特征进行了系统研究。依据摄食习性, 本文将调查海域出现的96种游泳动物划分为7个功能群, 其中底栖动物/游泳动物食性、游泳动物食性、底栖动物食性和腐屑食性功能群是该水域的优势功能群。相似性分析检验结果显示: 不同季节象山港游泳动物群落的功能群结构保持相对稳定, 不存在显著性季节差异。4月、7月和11月该水域游泳动物群落的功能群多样性均值分别为1.303、1.261和1.185, 亦未呈现显著性季节差异。功能群多样性指数与大个体生物(体重大于50g)的总重量资源密度显著正相关; 功能群多样性与物种多样性的关系可用斜率为0.463、截距为0.266的线性方程进行描述, 这表明该水域物种多样性的大量丧失可能引起功能群多样性的降低。象山港水域部分功能群所含物种数明显偏少, 冗余度较低, 在强大的外界干扰下, 某些重要物种的衰退可能将使该水域功能群多样性显著降低。
象山港; 游泳动物群落; 功能群; 功能群多样性; 物种冗余
游泳动物是海洋生态系统中的高级消费者, 在海洋生态系统中有着重要的地位; 同时, 它还是人类重要的优质蛋白来源, 是海洋渔业主要的利用对象(沈国英等, 2002)。近年来, 随着我国近海渔业资源严重衰退、海洋生态系统的服务和产出功能日益降低, 游泳动物群落研究作为引入基于生态系统的渔业资源管理策略的前期理论准备和必需环节(唐启升等, 2005), 逐渐受到人们的关注。在游泳动物群落生态研究中, “功能群”是一种简化群落结构的重要研究手段(Muñoz, 1998; Garrison, 2000; Bellwood, 2003)。功能群是指在生态系统中起着相似生态作用、并占据相似生态位的若干物种的集合体(Root, 1967)。在海洋生态学研究中, 功能群可以大大简化海洋生态系统的食物网和营养动力学过程研究, 并可为群落结构同群落功能性、稳定性以及抗干扰能力关系的研究提供一种新的视角(Micheli, 2005; Arenas, 2006)。
象山港是位于我国浙江省中部偏北沿海的半封闭狭长形海湾, 港域生态环境良好, 渔业资源丰富。近年来, 随着各类海洋开发活动的进行, 象山港生态环境不断恶化, 渔业资源显著衰退, 象山港生态系统保护和修复问题亟待解决(尤仲杰等, 2011)。掌握不同类群生物尤其是游泳动物的群落结构现状是开展海洋生态系统修复工作的基础。针对该海域游泳动物群落生态研究的薄弱现状, 本研究拟利用2011年开展的象山港水域渔业资源定点调查数据, 分析象山港游泳动物群落的功能群结构, 探究该群落功能群的多样性和冗余性特征, 以期丰富该水域生态系统结构和功能的理论认知, 并为系统开展象山港海洋生态系统的管理与修复提供基础信息。
1 材料与方法
1.1 站位布设及采样分析方法
本研究所用数据来源于2011年4月7—10日、7月22—25日和11月23—26日开展的象山港渔业资源定点调查。鉴于不同渔业作业方式对不同生态类型游泳动物的采样代表性存在较大差异, 本次调查的游泳动物采样采取桁杆拖虾网调查和单拖网调查相结合的方式。其中, 桁杆拖虾网调查的目标种类主要包括甲壳类和营贴底生活的鲽形目鱼类, 调查网具的桁杆长度为7m, 囊网网目尺寸为13mm, 航速约2km/h, 调查期间每站拖曳30min; 单拖网调查的目标种类主要包括鲽形目以外的鱼类和头足类, 调查网具的网口宽度为10m, 囊网网目尺寸为13mm, 航速约为4km/h, 调查期间每站拖曳15—30min。各航次定点调查的站位设置如图1所示。其中, 桁杆拖虾网调查共设置11个调查站位; 单拖网调查由于受部分目标调查海域的水深和地形状况限制, 仅能在4个调查站位开展单拖网调查, 具体站位分别为S03、S04、S07和S09。各站位游泳动物渔获样品经速冻保存, 带回实验室后鉴定和分析, 样品均鉴定到种, 长度测量精确到1mm, 重量精确到0.1g。
1.2 数据处理
利用扫海面积法估算各调查站点不同游泳动物种类的资源密度, 其中甲壳类和鲽形目鱼类的资源密度依据桁杆拖虾网的调查数据进行估算, 其它类型鱼类和头足类的资源密度依据单拖网调查资料进行估算。求算公式为:
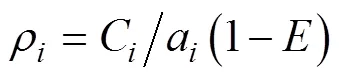
式中,为第站游泳动物的资源密度(重量密度: kg/km2; 尾数密度: 103ind/km2);C为第站每小时游泳动物的渔获量(重量密度: kg/h; 尾数密度: 103ind/h);为逃逸率, 本研究中底栖鱼类(含甲壳类)、中上层鱼类和底层鱼类(含头足类)的逃逸率分别取0.2、0.7和0.5(郑元甲等, 2003);a为调查船只每小时的扫海面积(km2/h), 即网口宽度×平均拖速。
将同一功能群中不同种类的资源密度进行汇总求和, 求得各功能群的资源密度。
1.3 数据分析
游泳动物功能群依据不同种类的摄食习性进行划分(Garrison, 2000; Floeter, 2004; 张波等, 2009)。
以不同月份桁杆拖虾网和单拖网共同调查站位(S03、S04、S07和S09)的资源密度数据为基础数据, 采用Shannon-Weiner多样性指数()来计算象山港游泳动物群落的功能群多样性和物种多样性指数, 具体计算公式如下(Micheli, 2006; 姜亚洲等, 2008; Shan, 2011; 吕振波等, 2012):
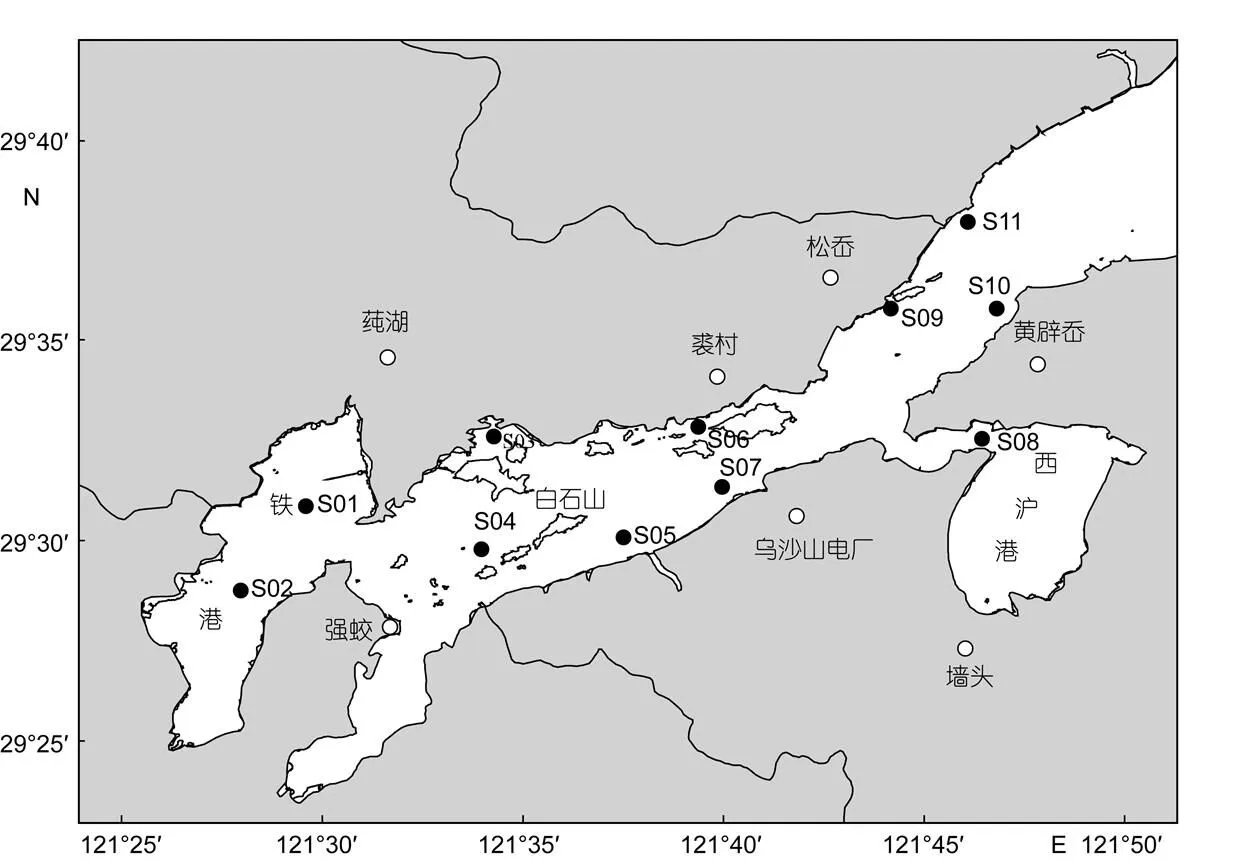
图1 2011年象山港渔业资源调查站位图
所有站位均开展桁杆拖虾网调查, 在S03、S04、S07和S09站位增加了单拖网调查
Shannon-Weiner 多样性指数() :
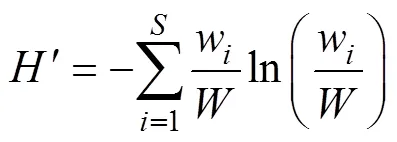
式中,为某调查站点的功能群个数(或物种数),w为该站位第种功能群(或物种)的重量资源密度,为该站位的总重量资源密度。
为分析物种多样性与功能群多样性的关系, 将以上述计算过程所获取的功能群多样性指数作为因变量、以物种多样性指数作为自变量, 分别利用直线方程、对数方程和指数方程进行回归分析, 3种回归分析结果的优劣依据拟合优度2加以判断(Micheli, 2006)。
以群落中的大个体生物(体重大于50g)的总重量资源密度作为反映群落功能的一项参数指标, 该指标可在一定程度上反映群落提供高营养层次生物的能力(Rochet, 2003; Greenstreet, 2006), 利用Pearson相关分析研究物种多样性、功能群多样性同该参数指标的关系, 以分析群落功能群多样性、物种多样性同群落功能间的关系。
利用相似性分析检验(ANOSIM)对不同月份象山港游泳动物群落的功能群结构进行差异显著性检验(Clarke, 1993), 该检验过程所用的基础数据为不同月份调查站位S03、S04、S07和S09(桁杆拖虾网和单拖网共同调查的站位)中各功能群的重量资源密度数据。
以上计算和统计过程利用SPSS14.0和Primer 5.0软件进行。
2 结果
2.1 功能群的划分
2011年象山港水域渔业资源调查共鉴定游泳动物种类96种, 其中鱼类54种, 甲壳类38种, 头足类4种。依据现有的关于游泳动物摄食习性的文献资料(魏崇德等, 1991; 张雅芝等, 1994; 徐善良等, 1996; 程济生等, 1997; 柏怀萍, 1999; 黄美珍, 2005; 倪勇等, 2006; 张波等, 2007, 2009; 庄平等, 2011; 纪炜炜, 2011; 吕振波等, 2012), 将象山港游泳动物划分7个功能群, 分别为浮游动物食性功能群、浮游动物/底栖动物食性功能群、底栖动物食性功能群、底栖动物/游泳动物食性功能群、游泳动物食性功能群、杂食性功能群和腐屑食性功能群。各功能群包含的主要种类(单航次调查中重量资源密度百分比大于0.5%的种类)如下:
浮游动物食性功能群, 主要包括黄吻棱鳀()和刺海马()等4个种类。
浮游动物/底栖动物食性功能群, 主要包括口虾蛄()、黑斑口虾蛄()、短蛸()、棘头梅童鱼()和中华管鞭虾()等12个种类。


杂食性功能群, 主要包括鲬() 和鳄鲬()等3个种类。

2.2 象山港游泳动物群落的功能群结构
ANOSIM分析结果表明: 不同季节象山港游泳动物群落的功能群结构保持相对稳定, 季节差异达不到显著性水平(=0.074,=0.308)。3个航次调查中象山港游泳动物的功能群组成状况如表1所示, 底栖动物食性、底栖动物/游泳动物食性、游泳动物食性和腐屑食性功能群是该水域的优势功能群, 其所占总重量资源密度的百分比范围分别为: 16.22%— 18.43%、32.43%—48.50%、12.45%—25.04%和1.42%— 20.43%。
在3航次调查中, 不同调查站位各功能群平均所含物种数如图2所示。底栖动物食性功能群平均所含物种数最高, 不同月份该值在11—14.25范围内; 其次为底栖动物/游泳动物食性和浮游动物/底栖动物食性功能群, 该值范围分别为5.25—7.25和2.75—4.75; 其余功能群平均所含物种数均较低, 该值均小于2。
表1 2011年4月、7月和11月象山港游泳动物群落功能群组成
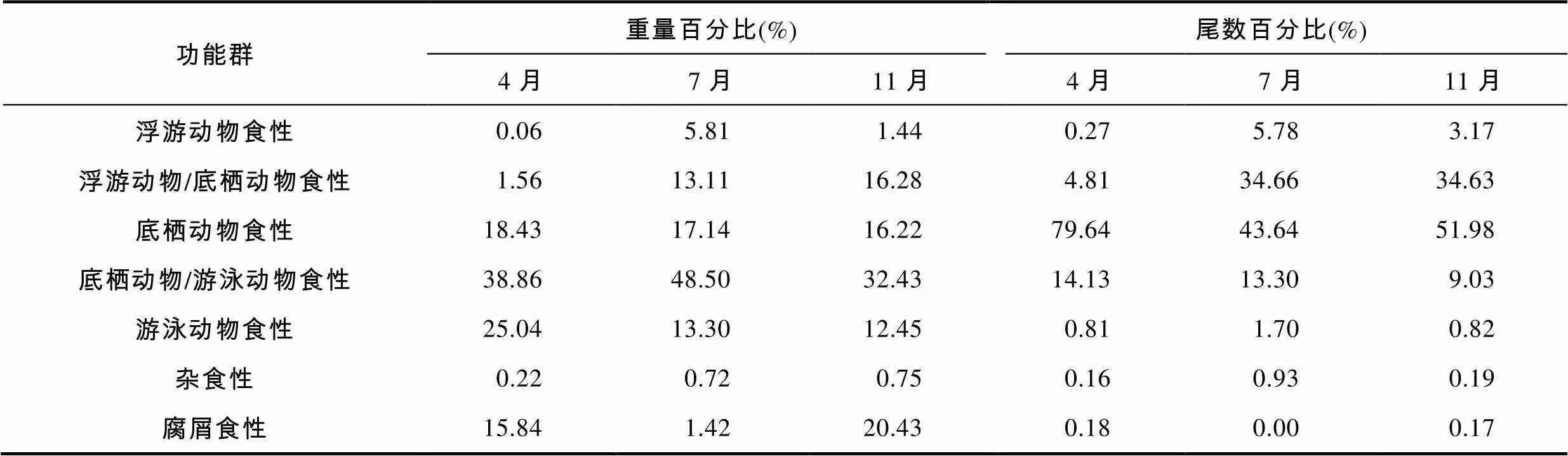
Tab.1 Proportion of different functional groups during April, July and November of 2011 in the Xiangshan Bay
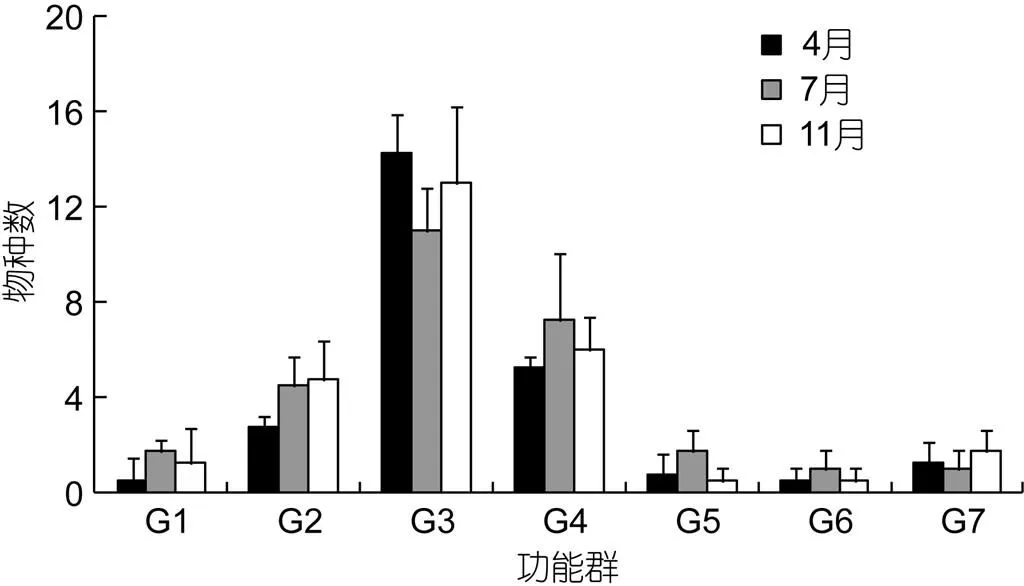
图2 2011年4月、7月和11月象山港游泳动物群落各功能群平均所含物种数
G1. 浮游动物食性功能群, G2. 浮游动物/底栖动物食性功能群, G3. 底栖动物食性功能群, G4. 底栖动物/游泳动物食性功能群, G5. 游泳动物食性功能群, G6. 杂食性功能群, G7. 腐屑食性功能群
2.3 象山港游泳动物群落的功能群多样性
4月、7月和11月象山港游泳动物群落的功能群多样性指数分别为1.303±0.151、1.261±0.333和1.185±0.343。方差分析结果显示: 功能群多样性指数不存在显著性季节差异(=0.172,=0.844)
本文对象山港游泳动物群落物种多样性与功能群多样性关系进行回归分析, 利用直线方程、对数方程和指数方程对该关系进行曲线拟合, 结果显示: 以斜率为0.463、截距为0.266的直线方程(图3)对该海域物种多样性和功能群多样性关系的拟合效果最优(拟合优度R=0.413,=7.03726,=10,=0.0242)。这表明: 如果该水域游泳动物的物种多样性降低, 该群落的功能群多样性可能会随之线性下降。
Pearson相关分析结果显示: 象山港游泳动物群落的功能群多样性指数与群落中大个体(体重大于50g)生物的总重量资源密度存在显著的正相关关系(<0.05), 其相关系数为0.471; 而物种多样性指数与其相关关系不显著(=0.667)。这表明: 该水域游泳动物群落所能提供的大个体生物的能力与功能群多样性密切相关, 功能群多样性越高, 群落所能提供的大个体生物的资源密度越大。
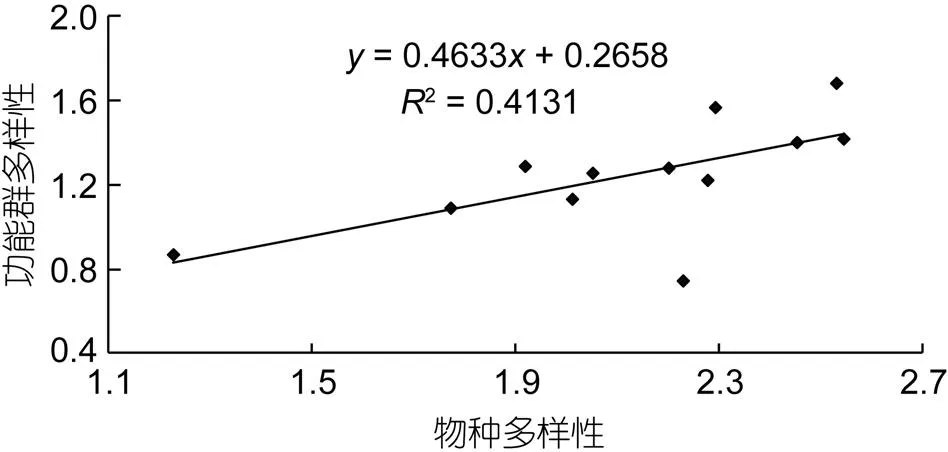
图3 象山港游泳动物群落物种多样性与功能群多样性的线性关系
3 讨论
3.1 象山港游泳动物群落功能群组成特征
象山港位于欧亚大陆东部的亚热带季风区, 冬夏季风交替显著, 四季分明(尤仲杰等, 2011)。象山港游泳动物群落的功能群组成并未受该水域水文条件季节变化的影响,不同季节其功能群结构保持相对稳定, 未呈现出显著性季节差异。底栖动物食性、底栖动物/游泳动物食性、腐屑食性和游泳动物食性功能群构成了象山港水域的优势功能群(表1)。与东海近海浮游动物食性功能群占据较大生物量比重不同, 象山港由于缺少大种群的浮游动物食性种类分布, 浮游动物食性功能群不构成该水域的优势功能群。
功能群是简化海洋生态系统食物网, 探索其物质、能量传递途径的重要手段(张雅芝等, 1994; 许思思等, 2010)。食物网中物质与能量的传递取决于各功能群的生物量构成(唐启升等, 2005)。基于象山港水域游泳动物群落的功能群组成特点, 象山港水域的物质、能量传递可大致归纳为以下3条途径: (1) 浮游植物→浮游动物→浮游动物食性功能群→游泳动物食性功能群; (2) 浮游植物和碎屑→底栖动物→底栖动物食性功能群→游泳动物食性功能群(或底栖动物/游泳动物食性功能群); (3) 碎屑→腐屑食性功能群。从各功能群的生物量配比上分析, 底栖动物食性功能群和底栖动物/游泳动物食性功能群为该水域的优势功能群, 第2条途径应是该水域的主要的物质、能量传递途径。与此同时, 腐屑食性功能群所占生物量比重亦较高, 在该水域物质、能量传递过程中亦起着重要作用。第1条传递途径受限于象山港水域浮游动物食性功能群的重量资源密度较低, 所起作用不甚明显。
3.2 象山港游泳动物群落功能群多样性与冗余性
功能群多样性是反映群落中功能群数目和均匀性的指标。从维持生态系统稳定性的角度来看, 功能群多样性比单纯的物种多样性更有意义。同一功能群的不同物种在生态系统中起着相似的作用, 它们之间的相互取代对生态系统功能不会产生太大的影响, 而如若某个功能群整体发生了变化甚至消失, 将会对整个生态系统产生巨大影响(Naeem, 2002; Bellwood, 2003 )。本研究发现, 象山港水域游泳动物群落的功能群多样性指数在1.185—1.303范围内, 明显高于与之毗邻的东海北部近海(姜亚洲等, 2008; Shan, 2011)。究其原因, 在所划分的功能群数目较东海北部近海较少的情况下, 象山港水域各功能群间的生物量分配相对均衡, 功能群组成的均匀性高于东海北部近海应是造成该水域功能群多样性较高的主要原因。在东海北部海域, 游泳动物食性、杂食性和浮游动物食性3个功能群的生物量占总渔获重量的比重合计高达95.64%, 其它功能群所占总渔获重量的比重均未超过2%(姜亚洲等, 2008); 而在象山港水域, 除浮游动物食性和杂食性功能群所占生物量百分比较低外, 其余各功能群的生物量组成较为均衡。
本文发现象山港游泳动物群落功能群多样性与群落中大个体生物的总资源密度间存在显著的正相关关系, 即功能群多样性越高, 所能提供的大个体生物的资源量越大。大个体生物的资源密度不仅可反应特定水域的渔业利用潜力; 同时, 鉴于个体较大的种类, 其营养层次通常较高, 大个体生物的资源密度亦可表征群落提供高营养层次生物的能力(Boudreau, 1991; Woodward, 2005)。高营养层次生物资源密度的高低可直接反应群落的物质、能量传递效率, 是评价群落功能的重要指标(Rochet, 2003; Greenstreet, 2006)。故此, 保护该水域游泳动物群落的功能群多样性具有双重意义, 它不仅可以保障该水域优质渔获产品的产出, 同时有利于该水域群落功能的正常发挥。
本文通过对物种多样性与功能群多样性关系的回归分析发现, 象山港水域游泳动物群落物种多样性与功能群多样性存在正相关关系(图3), 这表明该水域物种多样性大量丧失可能会引起功能群多样性的随之降低。但与东海近海相比, 该水域物种多样性与功能群多样性拟合直线的斜率较小(姜亚洲等, 2008), 这表明该水域较东海近海的物种冗余程度较大, 同等幅度的物种多样性丧失在该水域引起的功能群多样性降低幅度较小。
象山港水域游泳动物群落的功能群多样性及其冗余性虽较东海近海较高, 但在不断增强的外界干扰下, 该水域功能群多样性亦存有进一步丧失的风险。目前, 该水域一些功能群所含种类数较少(图2), 物种冗余性较低, 一些关键物种的衰退不仅可降低该水域的物种多样性, 也会导致功能群多样性的丧失。以游泳动物食性功能群为例, 游泳动物食性功能群是象山港海洋生态系统中营养级别最高的功能群, 对于控制该水域的能量传递起着重要作用。本次调查发现, 象山港游泳动物食性功能群仅含有7个种类, 且绝大多数生物量集中于中单一种类(中国花鲈), 该种类作为象山港最重要的渔业捕捞对象, 承受着较大捕捞压力, 如若其资源量出现衰竭, 游泳动物食性功能群的生物量将锐减, 进而对游泳动物群落的营养结构产生严重的负面影响。为此, 在制定象山港生态保护与修复措施的过程中, 应加大对关键物种和功能群的保护力度, 维持游泳动物群落营养结构的完整性和功能群多样性, 以确保象山港海洋生态系统的健康与稳定。
尤仲杰, 焦海峰, 2011. 象山港生态环境保护与修复技术研究. 北京: 海洋出版社, 1—444
吕振波, 李 凡, 徐炳庆等, 2012. 黄海山东海域春、秋季鱼类群落多样性. 生物多样性, 20(2): 207—214
庄 平, 罗 刚, 张 涛等, 2011. 长江口水域中华鲟幼鱼与6 种主要经济鱼类的食性及食物竞争. 生态学报, 31(20): 5544—5554
许思思, 宋金明, 段丽琴等, 2010. 渤海主要渔业资源结构的演变分析. 海洋科学, 34(6): 59—65
纪炜炜, 2011. 东海中北部主要游泳动物食物网结构和营养关系初步研究: 基于稳定同位素技术. 青岛: 中国科学院海洋研究所博士学位论文, 33—95
沈国英, 施并章, 2002. 海洋生态学. 北京: 科学出版社, 153—188
张 波, 金显仕, 唐启升, 2009. 长江口及邻近海域高营养层次生物群落功能群及其变化. 应用生态学报, 20(2): 344—351
张 波, 唐启升, 金显仕, 2007. 东海高营养层次鱼类功能群及其主要种类. 中国水产科学, 14(6): 939—949
张雅芝, 李福振, 刘向阳等, 1994. 东山湾鱼类食物网研究. 台湾海峡, 13(1): 52—61
郑元甲, 陈雪忠, 程家骅等, 2003. 东海大陆架生物资源与环境. 上海: 上海科学技术出版社, 1—835
柏怀萍, 1999. 象山港黑鲷的摄食习性. 宁波大学学报(理工版), 12(4): 42—47
姜亚洲, 程家骅, 李圣法, 2008. 东海北部鱼类群落多样性及其结构特征的变化.中国水产科学, 15(3): 453—459
倪 勇, 伍汉霖, 2006. 江苏鱼类志. 北京: 中国农业出版社, 1—963
徐善良, 王春琳, 梅文骧等, 1996. 浙江北部海区口虾蛄繁殖和摄食习性的初步研究. 浙江水产学院学报, 15(1): 30—35
唐启升, 苏纪兰, 张 经, 2005. 我国近海生态系统食物产出的关键过程及其可持续机理. 地球科学进展, 20(12): 1281—1287
黄美珍, 2005. 台湾海峡及邻近海域主要无脊椎动物食性特征及其食物关系研究. 海洋科学, 29(1): 73—80
程济生, 朱金声, 1997. 黄海主要经济无脊椎动物摄食特征及其营养层次的研究. 海洋学报, 19(6): 102—108
魏崇德, 陈永寿, 1991. 浙江动物志(甲壳类). 杭州: 浙江科学技术出版社, 1—481
Arenas F, Iñigo S, Stephen J H, 2006. The invasibility of marine algal assemblages: role of functional diversity and identity. Ecology, 87(11): 2851—2861
Bellwood D R, Hoey A, Choat J H, 2003. Limited functional redundancy in high diversity systems: resilience and ecosystem function on coral reefs. Ecology Letters, 6(4): 281—285
Boudreau P R, Dickie L M, Kerr S R, 1991. Body-size spectra of production and biomass as system-level indicators of ecological dynamics. Journal of Theoretical Biology, 152: 329—339
Clarke K R, 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology, 18(1): 117—143
Floeter S R, Ferreira C E L, Dominici-Arosemena A, 2004. Latitudinal gradients in Atlantic reef fish communities: trophic structure and spatial use patterns. Journal of Fish Biology, 64(6): 1680—1699
Garrison L P, 2000. Spatial and dietary overlap in the Georges Bank groundfish community. Canadian Journal of Fishery and Aquatic Sciences, 57(8): 1679—1691
Garrison L P, Link J S, 2000. Dietary guild structure of the fish community in the Northeast United States continental shelf ecosystem. Marine Ecology Progress Series, 202: 231—240
Greenstreet S P R, Rogers S, 2006. Indicators of the health of the North Sea fish community: identifying reference levels for an ecosystem approach to management. ICES Journal of Marine Science, 63(4): 573—593
Micheli F, Halpern B S, 2005. Low functional redundancy in coastal marine assemblages. Ecology Letters, 8(4): 391— 400
Muñoz A A, Ojeda F P, 1998. Guild structure of carnivorous intertidal fishes of the Chilean coast: implications of ontogenetic dietary shifts. Oecologia, 114(4): 563—573
Naeem S, 2002. Disentangling the impacts of diversity on ecosystem functioning in combinatorial experiments. Ecology, 83(10): 2925—2935
Rochet M J, Trenkel V M, 2003. Performance of indicators derived from abundance estimates for detecting the impact of fishing on a fish community. Canadian Journal of Fisheries and Aquatic Sciences, 60(1): 67—85
Root R B, 1967. The niche exploitation pattern of the blue-gray gnatcatcher. Ecological Monographs, 37(4): 317—350
Shan X J, Jin X S, Zhou Z P, 2011. Fish community diversity in the middle continental shelf of the East China Sea. Chinese Journal of Oceanology and Limnology, 29(6): 1199—1208
Woodward G, Ebenman B, Emmerson M, 2005. Body size in ecological networks. Trends in Ecology and Evolution, 20(7): 402—409
FUNCTIONAL GROUP COMPOSITION AND FUNCTIONAL DIVERSITY OF NEKTON COMMUNITY IN THE XIANGSHAN BAY
JIANG Ya-Zhou1, LIN Nan1, YUAN Xing-Wei1, JIAO Hai-Feng2, LI Sheng-Fa1
(1. East China Sea Fisheries Research Institute, Chinese Academy of Fisheries Sciences, Shanghai 200090, China; 2. Ningbo Academy of Oceanology and Fisheries, Ningbo 315012, China)
A functional group is a group of species that occupy the same niche in ecology and share resources in a similar way. In the concept of functional group, we simplify the structure and dynamics of nekton community to examine the energy flow and biomass within food webs in the Xiangshan Bay based on the data collected from beam-trawl and single-trawl surveys in April, July, and November of 2011. In terms of feeding habit, seven functional groups were identified: planktivores, planktivores/benthivores, benthivores, benthivores/piscivores, omnivores, piscivores, and detritivores. Similarity analysis revealed that there was no significant seasonal difference in structure of nekton community at functional group level. Benthivores/piscivores, benthivores, piscivores, and detritivores were dominant in biomass of the nekton community. The functional diversity in the bay was 1.303 in April, 1.261 in July, and 1.185 in November, with no significant monthly difference. A strong positive relationship was revealed between functional diversity and total biomass of these individuals whose body weights exceed 50g. A linear relationship between species and functional diversity could be well described in regression. In addition, the number of species in some given functional groups was small, indicative of a low level of functional redundancy. Currently, under the intense fishing pressure, some commercially important species might extinct locally, which might further lead to the removal of the whole functional group from the bay and irreversible degradation of ecosystem functions.
Xiangshan Bay; nekton community; functional group; functional diversity; species redundancy
10.11693/hyhz20120917001
* 国家公益性行业(农业)科研专项经费项目, 201003068号; 中央级公益性科研院所基本科研业务费专项(中国水产科学研究院东海水产研究所), 2011T08号。姜亚洲, E-mail: yazhoujiang@163.com
李圣法, 研究员, E-mail: shengfa@sh163.net
2012-09-17,
2012-12-17
S931
