温度和寄主植物对B/Q隐种烟粉虱生长发育和繁殖的联合影响
2014-04-11焦晓国周小毛王少丽吴青君张友军
易 胜, 焦晓国, 谢 文, 周小毛, 王少丽, 吴青君, 张友军*
(1.湖南农业大学农药研究所,长沙 410128;2.中国农业科学院蔬菜花卉研究所,北京 100081)
温度和寄主植物对B/Q隐种烟粉虱生长发育和繁殖的联合影响
易 胜1,2, 焦晓国2, 谢 文2, 周小毛1, 王少丽2, 吴青君2, 张友军2*
(1.湖南农业大学农药研究所,长沙 410128;2.中国农业科学院蔬菜花卉研究所,北京 100081)
摘要比较研究了不同温度和寄主植物对B/Q隐种烟粉虱若虫存活率、生长发育和雌虫繁殖和寿命的影响。结果表明,无论是B隐种还是Q隐种,一品红上的若虫存活率在27℃显著低于20℃。在同一寄主植物上,B隐种若虫和Q隐种若虫在27℃条件下发育速率显著快于20℃。无论是B隐种或Q隐种,饲养在同一寄主上,雌虫寿命和产卵量都与温度存在显著负相关。在同一温度和同一寄主上,Q隐种除若虫发育历期慢于B隐种外,Q隐种若虫存活率、雌虫产卵量和寿命都显著高于B隐种。可见,Q隐种在两种测定温度和两种测定寄主上较之B隐种具有竞争优势。研究结果表明B/Q隐种之间竞争取代可能受寄主、温度或其他外源因子的影响。
关键词烟粉虱; 温度; 寄主植物; 生长发育; 繁殖; 竞争取代
烟粉虱是严重为害蔬菜、花卉和主要田间作物的一种多食性全球重要害虫,是由形态上不易分辨的多种隐种组成的复合种[1-2]。在众多隐种中,仅B/Q隐种烟粉虱是重要外来入侵害虫,B隐种又称“MEAM1”(Middle East-Asia Minor 1),Q隐种又称“MED”(Mediterranean)。B/Q隐种烟粉虱在世界各地分布广泛,但各地分布比例不同且竞争替代结果也复杂多样。在西班牙东南部,Q隐种逐渐取代B隐种[3-4];在美国,B隐种在室外较Q隐种具有竞争优势,相反在温室内,Q隐种较B隐种具有竞争优势;在以色列,B/Q隐种能共存[5];而在东亚及中国大部分地区,Q隐种已经逐渐取代早期入侵的B隐种成为烟粉虱危害的主要致害隐种[6-7]。这说明影响B/Q隐种分布比例和竞争替代结果的因子多样且复杂。先前室内研究表明,B隐种由于非对称性交配干扰而对Q隐种具有竞争优势[8-10];而在西班牙东南部和中国大部,一般把室外Q隐种具有竞争优势归因于Q隐种较B隐种对烟碱类杀虫剂抗性更强[11]。除这些内源影响因子外,外源因子,如气候、温湿度和寄主植物分布等也可能影响两种隐种的竞争替代结果[12-14]。
植食性昆虫经受众多不同环境因子的复杂作用,而这些因子对植食性昆虫存活、生长发育和繁殖等生活史性状能产生复杂影响[15]。目前,众多研究集中于单一生态因子对植食性昆虫的影响[16],不同生态因子对植食性昆虫联合影响少见报道。温度和寄主是诱导植食性昆虫产生表型变异的两个重要因子[17]。通常,高温会导致植食性昆虫幼虫发育历期缩短,成虫体型变小[18]以及繁殖力提高[19]。而寄主植物对植食性昆虫生活史性状变异也有一定影响,因为植物营养成分和次生代谢产物浓度依寄主植物物种不同而异[20]。植食性昆虫取食低质量寄主植物通常会导致幼虫存活率低[21],生长速率慢,发育历期长,成虫个体小和雌虫生殖力低。尽管温度或寄主植物对植食性昆虫存活、生长发育和生活史性状研究多见,温度和寄主植物对植食性昆虫生活史性状联合影响却少见研究[22]。尽管先前研究结果表明烟粉虱生活史性状受温度[23-26]和寄主植物显著影响[27-30],但两因子对B/Q隐种的联合影响却鲜有报道。本文探讨比较B/Q隐种对两种寄主(棉花和一品红)和两个温度(20℃和27℃)的联合反应,以期为B/Q隐种的竞争取代机制提供理论依据。
1 材料及方法
1.1 供试昆虫来源和寄主植物
B隐种2004年采集于北京地区甘蓝(Brassica oleracea‘Jingfeng1’)上,室内用棉花(Gossypium hirsutum‘Baoling DP99B’)继代饲养至今。Q隐种2008年采自一品红(Euphorbia pulcherrima),室内用一品红继代培养至今。每一隐种种群采用外罩防虫网的养虫笼培养(50 cm×40 cm×60 cm),培养条件为(25±2)℃,60%±10%(RH)和L∥D=14 h∥10 h光照。每饲养3代,采用mtCOI引物进行PCR扩增,监测鉴别烟粉虱纯度[31]。
1.2 若虫存活率和发育历期
隐种和寄主植物的组合分为4个处理,BC(B隐种雌虫卵产于棉花上);BP(B隐种雌虫卵产于一品红上);QP(Q隐种雌虫卵产于一品红上)和QC(Q隐种雌虫卵产于棉花上)。把16株无虫的一品红或棉花放入饲养B隐种或Q隐种的养虫笼中接卵,接卵10 h后,把带卵寄主植物从养虫笼中移出。每一植株选取3~4片伸展的叶片并标记。未标记叶片上的卵在双目显微镜下去除。选择叶片下端有卵地方采用记号笔标记,利于跟踪每头烟粉虱由卵到成虫羽化的过程。每选择叶片上卵量大约20~50粒,多余卵量去除。4个处理(BC,BP,QP和QC)中,每一处理采用4株植物分别放入20℃和27℃人工气候箱中培养,培养箱保持RH60%±10%和L∥D=14 h∥10 h光照。当某一寄主上发现有烟粉虱伪蛹时,将该寄主植株采用防虫网罩住,然后每天收集羽化的成虫并冰冻于-20℃冰箱中。计算不同温度下,不同烟粉虱隐种若虫在不同寄主植物上发育历期(卵至成虫)。若虫存活率采用羽化成虫数与初始卵量比值表示。在每一处理中,每一选择叶片为一次重复,每处理12~13个重复。
1.3 雌虫产卵量和寿命
寄主植物和隐种4种组合如前所述。把刚羽化的雌雄成虫配对,采用微虫笼接于寄主植物叶片下端,每植株接一个微虫笼,每处理在20℃或27℃条件下重复23~25次。每天观察雌虫存活状态,每隔2 d把微虫笼位置转移一次,记载每次雌虫产卵量。
1.4 数据统计方法
以隐种、温度和寄主植物为固定因子,采用普通线性模型(GLM)测定不同因子对烟粉虱若虫存活率、发育历期、雌虫产卵量和寿命的影响。若虫存活率数据在统计前采用平方根反正弦变换,所有数据采用SPSS 11.5(SPSS Inc,Chicago,IL,USA)进行统计分析。
2 结果与分析
2.1 若虫存活率和发育历期
若虫存活率在不同隐种、不同寄主植物和不同温度下差异显著(表1)。在同一温度和同一寄主植物上培养时,Q隐种存活率显著高于B隐种(图1)。在棉花上,B/Q隐种若虫在27℃条件下存活率都显著高于20℃;相反在一品红上,B/Q隐种若虫在20℃条件下存活率均显著高于27℃(图1)。同时,寄主植物和温度互作对烟粉虱若虫存活率也有显著影响(表1)。
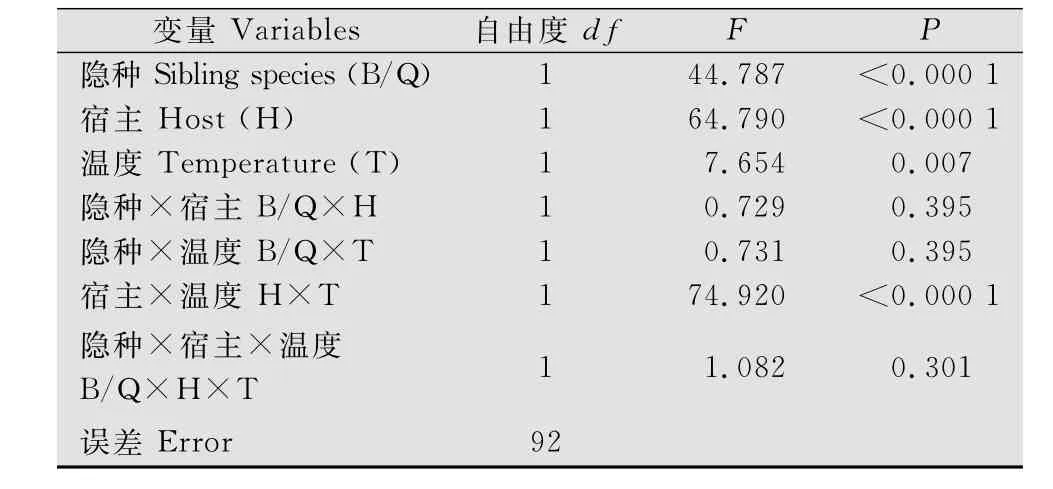
表1 温度和寄主植物对B/Q隐种若虫存活率的影响Table 1 Effects of temperature and host plants on immature survivorship(egg to adult)of B and Q sibling species of B.tabaci

图1 温度和寄主植物对B/Q隐种若虫存活率(means±SD)的影响Fig.1 Immature survivorship(means±SD)of B and Q sibling species of B.tabaci as affected by temperature and host
B/Q隐种卵至成虫发育历期在27℃下显著短于20℃(图2)。在同一寄主植物上,B隐种发育速率显著快于Q隐种(图2)。同时,B/Q隐种在棉花上发育速率都快于一品红(图2)。隐种和温度互作以及寄主植物和温度互作对若虫发育时间也存在显著影响(表2)。

表2 温度和寄主植物对B/Q隐种若虫发育历期的影响Table 2 Effects of temperature and host plants on immature development time(egg to adult)of B and Q sibling species of B.tabaci

图2 温度和寄主植物对B/Q隐种若虫发育历期(means±SD)的影响Fig.2 Developmental time(means±SD)(egg to adult)of B and Q sibling species of B.tabaci as affected by temperature and host
2.2 雌虫产卵量和寿命
雌虫产卵量受隐种、寄主植物和温度的显著影响(表3)。Q隐种产卵量显著高于B隐种(图3)。无论是B还是Q隐种,在20℃雌虫产卵量显著高于27℃,在棉花上产卵量显著高于一品红(图3)。隐种和温度、隐种和寄主植物、隐种和温度及寄主植物等互作都对烟粉虱产卵量有显著影响(表3)。

图3 温度和寄主植物对B/Q隐种雌虫产卵量(means±SD)的影响Fig.3 Fecundity(means±SD)of B and Q sibling species of B.tabaci as affected by temperature and host
雌虫寿命受隐种、寄主植物和温度的显著影响(表4)。在同一温度或同一寄主植物上,Q隐种寿命显著长于B隐种(图4)。无论是B还是Q隐种,雌虫寿命在20℃下显著长于27℃,在棉花上寿命显著长于一品红(图4)。寄主植物和温度、隐种和温度及寄主植物等互作都对烟粉虱寿命都有显著影响(表4)。

表3 温度和寄主植物对B/Q隐种雌虫产卵量的影响Table 3 Effects of temperature and host plants on female lifetime fecundity of B and Q sibling species of B.tabaci

表4 温度和寄主植物对B/Q隐种雌虫寿命的影响Table 4 Effects of temperature and host plants on female adult longevity of B and Q sibling species of B.tabaci
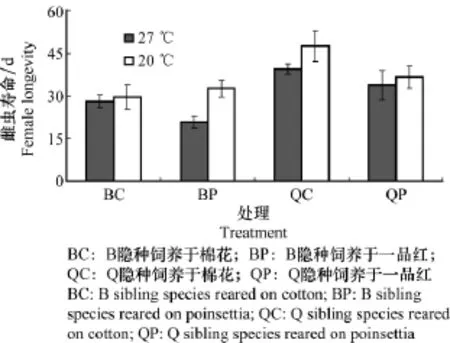
图4 温度和寄主植物对B/Q隐种雌虫寿命(means±SD)的影响Fig.4 Female longevity(means±SD)of B and Q sibling species of B.tabaci as affected by temperature and host
3 结论与讨论
本文研究结果表明,寄主植物和温度能显著影响B/Q隐种的存活率、生长发育和繁殖等生活史性状。先前研究发现不同寄主植物对B/Q隐种许多适合度参数均有显著影响[32],尽管棉花和一品红是B/Q隐种的适宜寄主,但棉花和一品红对B/Q隐种烟粉虱的影响却存在显著差异。例如,无论是B或Q隐种,在一品红上较之棉花上,若虫存活率低,发育时间延长,雌虫寿命和产卵量都低。笔者先前研究发现,棉花和一品红中主要营养物质数量和质量并没有显著差异[33],但两种寄主植物中次生物质却有明显差异,如总酚(一类抑制植食性昆虫取食的重要次生抗虫物质),其在一品红中含量是棉花中7倍。所以较之棉花,一品红中高含量酚类可能是导致其对B/Q隐种烟粉虱适宜性较低的重要原因。无论是B还是Q隐种,在一品红上取食的若虫存活率在27℃下显著低于20℃。Wagner对棉花上烟粉虱B隐种测定结果表明,次生物质对植食性昆虫不利影响程度与温度存在正相关关系[34]。本文结果也表明,次生物质对烟粉虱不利影响程度在高温下显著大于在低温条件下,两者结果一致。同时在先前提及的生物学参数方面,B/Q隐种对寄主植物和温度的反应基本一致。在测定的4个生物学参数方面,寄主植物和温度的互作显著影响到其中3个参数,包括若虫存活率、若虫发育历期和雌虫寿命。在这些生物学参数中,寄主和温度互作对若虫存活率的影响非常有意思,B/Q隐种的若虫在棉花上,27℃存活率显著高于20℃;相反在一品红上,烟粉虱若虫存活率在20℃显著高于27℃。寄主次生物质和热量动态复杂的相互作用可能导致该差异。其具体作用机制还需进一步研究。总之,本研究结果表明,温度和寄主植物对烟粉虱生物学性状的影响有些较为简单,有些可能存在复杂的相互作用,它们的影响可能依所测定的烟粉虱性状而异。在这里要指出的是B/Q隐种存活率数据变异过大,可能与处理重复数过小有关。也可能与选取的寄主植物叶龄差异过大有关,不同龄期的叶子中次生化学物质可能存在明显差异,进而导致B/Q隐种存活率数据出现较大变异。
已有研究报道表明,烟粉虱若虫存活、生长发育和生物学性状受不同寄主植物,甚至同一寄主植物不同品种和温度的显著影响。目前我们仅测定比较B/Q隐种在两种寄主和两种温度下的生物学表现,尽管发现B/Q隐种对不同温度和寄主的反应模式相似,但隐种之间还是存在一定差异,如在同一温度和同一寄主上,Q隐种若虫存活率、雌虫寿命和产卵量都相应高于B隐种。该结论至少可以部分解释在西班牙东南部和中国大部Q隐种烟粉虱正在取代B隐种烟粉虱的事实[7]。
先前研究结果认为不同地区烟粉虱不同隐种之间竞争优势可能受内源因素影响,如非对称性交配干扰[8]、对杀虫剂的抗性差异[11]和隐种本身的遗传多样性。不同隐种之间竞争优势也可能受外源因素影响,如寄主植物、气候和杀虫剂的喷洒等。且本研究结果发现,在不同测定温度下,Q隐种在棉花和一品红上表现均要优于B隐种。我们先前研究结果也表明,当同时把10对B/Q隐种烟粉虱分别释放于3种寄主植物一品红、甘蓝和黄瓜上,经过一段时间之后发现,B隐种烟粉虱在甘蓝和黄瓜上占优势,而Q隐种烟粉虱在一品红上占优势(未发表资料),可见外源因素温度和寄主植物也能影响B/Q隐种之间的竞争结果。
总之,本研究结果发现Q隐种在两种测定温度和两种测定寄主上较之B隐种具有竞争优势。B/Q隐种烟粉虱之间竞争结果可能受寄主、温度和其他外源因子的影响。但本研究结果还需要进一步采用更多寄主植物和更多温度梯度进行验证,以期为B/Q隐种的竞争取代机制提供理论依据。
参考文献
[1] De Barro P J,Liu S S,Boykin L M,et al.Bemisia tabaci:a statement of species status[J].Annual Review of Entomology,2011,56:1-19.
[2] De Barro P J.The Bemisia tabaci species complex:questions to guide future research[J].Journal of Integrative Agriculture,2012,11(2):187-196.
[3] Pascual S,Callejas C.Intra-and interspecific competition between biotypes B and Q of Bemisia tabaci(Hemiptera:Aleyrodidae)from Spain[J].Bulletin of Entomological Research,2004,94(4):369-375.
[4] Pascual S.Short communication.Mechanisms in competition,under laboratory conditions,between Spanish biotypes B and Q of Bemisia tabaci(Gennadius)[J].Spanish Journal of Agricultural Research,2006,4(4):351-354.
[5] Horowitz A R,Denholm I,Gorman K,et al.Biotype Q of Bemisia tabaci identified in Israel[J].Phytoparasitica,2003,31(1):94-98.
[6] Chu D,Wan F H,Zhang Y J,et al.Change in the biotype composition of Bemisia tabaci in Shandong Province of China from 2005 to 2008[J].Environmental Entomology,2010,39(3):1028-1036.
[7] Pan H,Chu D,Ge D,et al.Further spread of and domination by Bemisia tabaci(Hemiptera:Aleyrodidae)biotype Q on field crops in China[J].Journal of Economic Entomology,2011,104(3):978-985.
[8] Liu S S,De Barro P J,Xu J,et al.Asymmetric mating interactions drive widespread invasion and displacement in a whitefly[J].Science,2007,318(5857):1769-1772.
[9] Crowder D W,Horowitz A R,De Barro P J,et al.Mating behaviour,life history and adaptation to insecticides determine species exclusion between whiteflies[J].Journal of Animal E-cology,2010,79(3):563-570.
[10]Crowder D W,Sitvarin M I,Carrière Y.Plasticity in mating behaviour drives asymmetric reproductive interference in whiteflies[J].Animal Behaviour,2010,79(3):579-587.
[11]Horowitz A R,Kontsedalov S,Khasdan V,et al.Biotypes B and Q of Bemisia tabaci and their relevance to neonicotinoid and pyriproxyfen resistance[J].Archives of Insect Biochemistry and Physiology,2005,58(4):216-225.
[12]De Barro P J,Bourne A,Khan S A,et al.Host plant and biotype density interactions–their role in the establishment of the invasive B biotype of Bemisia tabaci[J].Biological Invasions,2006,8(2):287-294.
[13]Iida H,Kitamura T,Honda K.Comparison of egg-hatching rate,survival rate and development time of the immature stage between B-and Q-biotypes of Bemisia tabaci(Gennadius)(Homoptera:Aleyrodidae)on various agricultural crops[J].Applied Entomology and Zoology,2009,44(2):267-273.
[14]De Barro P J,Bourne A.Ovipositional host choice by an invader accelerates displacement of its indigenous competitor[J].Biological Invasions,2010,12(9):3013-3023.
[15]Stillwell R C,Wallin W G,Hitchcock L J,et al.Phenotypic plasticity in a complex world:interactive effects of food and temperature on fitness components of a seed beetle[J].Oecologia,2007,153(2):309-321.
[16]Nylin S,Gotthard K.Plasticity in life-history traits[J].Annual Review of Entomology,1998,43(1):63-83.
[17]Awmack C S,Leather S R.Host plant quality and fecundity inherbivorous insects[J].Annual Review of Entomology,2002,47(1):817-844.
[18]Kingsolver J G,Huey R B.Size,temperature,and fitness:three rules[J].Evolutionary Ecology Research,2008,10(2):251.
[19]Stillwell R C,Fox C W.Complex patterns of phenotypic plasticity:interactive effects of temperature during rearing and oviposition[J].Ecology,2005,86(4):924-934.
[20]Hwang S Y,Liu C H,Shen T C.Effects of plant nutrient availability and host plant species on the performance of two Pieris butterflies(Lepidoptera:Pieridae)[J].Biochemical Systematics and Ecology,2008,36(7):505-513.
[21]Shobana K,Murugan K,Naresh Kumar A.Influence of host plants on feeding,growth and reproduction of Papilio polytes(The common mormon)[J].Journal of Insect Physiology,2010,56(9):1065-1070.
[22]Stamp N E,Yang Y.Response of insect herbivores to multiple allelochemicals under different thermal regimes[J].Ecology,1996:1088-1102.
[23]Enkegaard A.The poinsettia strain of the cotton whitefly,Bemisia tabaci(Homoptera:Aleyrodidae),biological and demographic parameters on poinsettia(Euphorbia pulcherrima)in relation to temperature[J].Bulletin of Entomological Research,1993,83(4):535-546.
[24]Wang K,Tsai J H.Temperature effect on development and reproduction of silverleaf whitefly(Homoptera:Aleyrodidae)[J].Annals of the Entomological Society of America,1996,89(3):375-384.
[25]Drost Y C,van Lenteren J C,van Roermund H J W.Life-history parameters of different biotypes of Bemisia tabaci(Hemiptera:Aleyrodidae)in relation to temperature and host plant:a selective review[J].Bulletin of Entomological Research,1998,88(03):219-230.
[26]Muñiz M,Nombela G.Differential variation in development of the B-and Q-biotypes of Bemisia tabaci(Homoptera:Aleyrodidae)on sweet pepper at constant temperatures[J].Environmental Entomology,2001,30(4):720-727.
[27]Tsai J H,Wang K.Development and reproduction of Bemisia argentifolii(Homoptera:Aleyrodidae)on five host plants[J].Environmental Entomology,1996,25(4):810-816.
[28]Muniz M.Host suitability of two biotypes of Bemisia tabaci on some common weeds[J].Entomologia Experimentalis et Applicata,2000,95(1):63-70.
[29]Gachoka K K,Obeng-Ofori D,Danquah E Y.Host suitability of two Ghanaian biotypes of Bemisia tabaci(Homoptera:Aleyrodidae)on five common tropical weeds[J].International Journal of Tropical Insect Science,2005,25(4):236.
[30]Zang L S,Chen W Q,Liu S S.Comparison of performance on different host plants between the B biotype and a non-B biotype of Bemisia tabaci from Zhejiang,China[J].Entomologia Experimentalis et Applicata,2006,121(3):221-227.
[31]Zhang L P,Zhang Y J,Zhang W J,et al.Analysis of genetic diversity among different geographical populations and determination of biotypes of Bemisia tabaci in China[J].Journal of Applied Entomology,2005,129(3):121-128.
[32]Bethke J A,Paine T D,Nuessly G S.Comparative biology,morphometrics,and development of two populations of Bemisia tabaci(Homoptera:Aleyrodidae)on cotton and poinsettia[J].Annals of the Entomological Society of America,1991,84(4):407-411.
[33]Jiao X G,Xie W,Wang S L,et al.Host preference and nymph performance of B and Q putative species of Bemisia tabaci on three host plants[J].Journal of Pest Science,2012,85(4):423-430.
[34]Wagner T L.Temperature-dependent development,mortality,and adult size of sweet potato whitefly biotype B(Homoptera:Aleyrodidae)on cotton[J].Environmental Entomology,1995,24(5):1179-1188.
中图分类号:Q 968
文献标识码:A
DOI:10.3969/j.issn.0529-1542.2014.03.004
收稿日期:2013-07-17
修订日期:2013-10-10
基金项目:国家科技支撑计划课题(2012BAD1901);公益性行业(农业)科研专项(201303028)
*通信作者Tel:010-82109518,E-mail:zhangyoujun@caas.cn
Combined effects of temperature and host plants on the development and reproduction of B and Q sibling species of Bemisia tabaci
Yi Sheng1,2, Jiao Xiaoguo2, Xie Wen2, Zhou Xiaomao1, Wang Shaoli2, Wu Qingjun2, Zhang Youjun2
(1.Institute of Pesticide Science,Hunan Agricultural University,Changsha 410128,China;2.Institute of Vegetables and Flowers,Chinese Academy of Agricultural Sciences,Beijing 100081,China)
AbstractIn the present study,we conducted a reciprocal cross experiment with two hosts(cotton and poinsettia)between B and Q sibling species of Bemisia tabaci to investigate the combined effects of rearing temperature(20℃and 27℃)and host plants on development and several life history traits of B and Q B.tabaci.We found that the survivorship of B.tabaci immatures(egg to adult),irrespective of B and Q,was independent of rearing temperature and host plants.Females and males of either biotype reared at 27℃developed substantially faster than those at 20℃.However,female lifetime fecundity and female adult longevity at 20℃were significantly higher than those at 27℃on both host plants.Cotton was more suitable for either B or Q relative to poinsettia.Q gains advantages over B in terms of higher lifetime fecundity and female adult longevity,except slower development at the same rearing temperature and on the same host,which may at least in part shed some light on the displacement of B by Q.
Key wordsBemisia tabaci; temperature; host plant; development; reproduction; competitive displacement
