不同生育期花生叶片蛋白质含量及氮代谢相关酶活性分析
2014-04-09翁伯琦郑向丽徐国忠王俊宏叶花兰
翁伯琦, 郑向丽, 赵 婷, 徐国忠, 王俊宏, 叶花兰
(福建省农业科学院农业生态研究所 福建省山地草业工程技术研究中心福建省丘陵地区循环农业工程技术研究中心, 福建 福州 350013)
福建是个多山省份,山地面积达10×104km2,其中海拔80 m以上的丘陵山地占89%,全省人均耕地仅为0.037 hm2, 低于FAO规定的耕地安全警戒线(0.067 hm2),土地资源紧张的现状迫使人们将农业开发目标转向占福建省总面积84.1%的丘陵山地。花生(ArachishypogaeaLinn.)是重要的油料作物和经济作物,也是发展中国家重要的食用植物油和蛋白质来源[1]。目前福建省的花生种植面积约107 km2,总产量达到25×104t[2],但只能满足自身需求的50%。因此,筛选或选育出适宜红壤丘陵山地种植的花生品种,且使花生荚果及花生秆的产量提高,是当前花生育种的重要目标之一。
大量的研究结果[3-11]表明:农作物体内的酶活性以及蛋白质代谢水平受品种基因型、光照、温度以及栽培管理措施等多种因素的影响。在花生植株中氮素营养代谢是一个重要的生理过程,直接影响花生的产量和品质[12];硝酸还原酶(NR)、谷氨酰胺合成酶 (GS)和谷氨酸脱氢酶(GDH)等是植物氮代谢过程中的关键酶,它们的活性直接影响花生对无机氮的利用;而蛋白质作为氮素代谢的终极产物,也与花生的品质正相关。近年来,有关干旱胁迫、氮肥施用量及其种类、植物生长调节剂以及微量元素对花生叶片NR及根瘤固氮酶活性方面的研究报道较多[13-17],且在不同粒型花生全生育期内各器官氮代谢相关酶活性方面也取得了一些研究结果[12],但有关红壤条件下仁秆两用型花生新品种(系)不同生育期内叶片氮代谢相关酶活性的研究尚未见报道。
作者以福建省农业科学院农业生态研究所筛选的5个珍珠豆型花生品种(系)为研究对象,对影响花生产量及品质的3个关键生育期(花针期、结荚期和饱果期)内叶片氮代谢相关酶活性和蛋白质含量的变化状况进行了分析,并比较了收获期花生的荚果产量和秆产量,旨在为红壤条件下花生的优质高产栽培及品种选育提供实验数据。
1 材料和方法
1.1 材料
供试5个花生品种(系)为福建省农业科学院农业生态研究所辐射选育的仁秆两用型花生新品系‘汕G’(‘Shan G’)(经250 KR剂量辐照后品种‘汕油71’的后代)和‘TJ’(经250 KR剂量辐照后品种‘闽科TA’的后代)及筛选的仁秆两用型花生新品种‘汕E’(‘Shan E’)(来源于‘汕油71’)、‘TH’(来源于‘闽科TA’)和‘泉花7号’(‘Quanhua No.7’)(福建省当家品种)。原始育种材料均由泉州市农业科学研究所提供。
1.2 方法
1.2.1 播种和栽培及采样方法 实验于2011年在福建省农业科学院农业生态研究所网室内进行。采用盆栽方式,花盆直径35 cm、高25 cm,每盆装干土23 kg。所用土壤为红壤,含有机质11.65 g·kg-1、速效氮73.8 mg·kg-1、速效磷3.04 mg·kg-1和速效钾60.75 mg·kg-1。按氮肥(尿素)300 kg·hm-2、磷肥(磷酸钙,12% P2O5)150 kg·hm-2和钾肥(硫酸钾,45% K2O)300 kg·hm-2的施用量在播种前施肥。于3月31日播种,每盆3穴,每穴播2粒,于“三叶一心”期间苗,每穴留苗1株。每个品种(系)为1个处理,共5个处理,每处理各设6次重复,随机排列。
观察花生植株的生长发育情况,并分别于花针期、结荚期和饱果期采集供试样品。叶片样品于上午9:00至11:00采集;采集主茎上倒数第3及第4片复叶,每盆采集3株,每处理各采集3盆;混合后用湿纱布包好并保存于-4 ℃条件下,迅速带回实验室,用于可溶性蛋白质及酶活性的测定。另外3盆则在收获期分别收获后考种计产。
1.2.2 可溶性蛋白质含量及酶活性测定 参照文献[18]中的Lorry法测定可溶性蛋白质含量,以牛血清蛋白做标准曲线,用考马斯亮蓝G-250显色,并于波长595 nm处测定OD值。参照文献[19]的方法测定叶片中硝酸还原酶(NR)活性;参照文献[20]的方法测定叶片中谷氨酰胺合成酶(GS)和谷氨酸脱氢酶(GDH)活性。
1.2.3 产量测定 大田收获时,各品种(系)取余下的3盆共9株,收获荚果及秆,分别称取单株荚果鲜质量和单株秆鲜质量。
1.3 数据处理
采用DPS 7.05数据处理系统对实验数据进行分析。
2 结果和分析
2.1 不同生育期可溶性蛋白质含量的变化
不同生育期(花针期、结荚期和饱果期)供试的5个花生品种(系)叶片中可溶性蛋白质含量的变化见表1。结果显示:不同生育期供试的5个花生品种(系)叶片可溶性蛋白质含量的变化趋势基本一致,但含量高低有差异;总体上看,各品种(系)叶片可溶性蛋白质含量在花针期较低,至结荚期达到最高,饱果期则略有下降,呈现“低—高—低”的变化趋势。
花针期和结荚期‘泉花7号’叶片可溶性蛋白质含量均最高,‘汕E’ 叶片可溶性蛋白质含量均最低;饱果期‘汕G’叶片可溶性蛋白质含量最高,而‘TH’叶片可溶性蛋白质含量最低,其中,‘TJ’、‘汕E’、‘泉花7号’和‘TH’叶片可溶性蛋白质含量分别比‘汕G’低21.55%、22.62%、23.94%和33.14%。差异显著性分析结果显示:花针期‘汕E’ 叶片可溶性蛋白质含量显著低于其他品种(系)(P<0.05);结荚期‘汕E’和‘TH’叶片可溶性蛋白质含量显著低于其他品种(系)(P<0.05);饱果期‘汕G’叶片可溶性蛋白质含量极显著高于其他品种(系)(P<0.01)。
总体上看,5个花生品种(系)中,‘汕G’和‘泉花7号’叶片可溶性蛋白质含量较高,‘汕E’ 叶片可溶性蛋白质含量较低。
2.2 不同生育期氮代谢相关酶活性的变化
2.2.1 硝酸还原酶(NR)活性的变化 不同生育期(花针期、结荚期和饱果期)供试的5个花生品种(系)叶片中硝酸还原酶(NR)活性的变化见表2。结果显示:在3个生育期内,‘泉花7号’叶片NR活性呈“高—低—高”的变化趋势,表现为花针期最高、结荚期降低、饱果期则略有上升;而其他4个品种(系)叶片NR活性均逐渐降低,变化趋势基本一致。
在花针期、结荚期和饱果期,5个品种(系)中,‘泉花7号’叶片的NR活性均最高,花针期和饱果期‘TJ’叶片的NR活性均最低,结荚期‘汕G’ 叶片的NR活性最低。差异显著性分析结果显示:在花针期,‘泉花7号’、‘汕E’和‘汕G’叶片的NR活性差异不显著(P>0.05),但均显著(P<0.05)或极显著(P<0.01)高于‘TH’或‘TJ’叶片的NR活性;在结荚期,‘泉花7号’、‘汕E’和‘TH’叶片的NR活性差异不显著(P>0.05),但均显著(P<0.05)或极显著(P<0.01)高于‘TJ’或‘汕G’叶片的NR活性;在饱果期,‘汕E’、‘汕G’、‘TH’和‘TJ’叶片的NR活性差异不显著,但均极显著(P<0.01)低于‘泉花7号’叶片的NR活性。
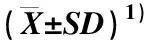
表1 不同生育期5个花生品种(系)叶片可溶性蛋白质含量的比较
表2 不同生育期5个花生品种(系)叶片硝酸还原酶(NR)活性的比较
总体上看,5个品种(系)中,‘泉花7号’、‘汕E’和‘汕G’叶片的NR活性较高,‘TJ’叶片的NR活性较低。
2.2.2 谷氨酰胺合成酶(GS)活性的变化 不同生育期(花针期、结荚期和饱果期)供试的5个花生品种(系)叶片中谷氨酰胺合成酶(GS)活性的变化见表3。结果显示:在3个生育期内,供试的5个花生品种(系)叶片GS活性的变化趋势有一定差异。
在3个生育期内,‘汕E’、‘TJ’和‘泉花7号’叶片的GS活性呈逐渐降低趋势,即GS活性在花针期最高、饱果期降至最低;‘汕G’和‘TH’叶片GS活性呈“低—高—低”的变化趋势,但花针期和结荚期GS活性差异较小,而饱果期GS活性明显降低。差异显著性分析结果显示:在花针期,‘TJ’叶片GS活性最高,极显著高于其他4个品种(系);而‘汕G’和‘TH’叶片GS活性则极显著低于其他品种(系)。在结荚期,‘汕E’叶片GS活性极显著高于其他4个品种(系),而‘TJ’叶片GS活性显著或极显著低于其他4个品种(系)。在饱果期,5个花生品种(系)叶片GS活性差异不大,仅‘汕G’叶片的GS活性显著低于其他4个品种(系)。
总体上看,5个品种(系)中,‘汕E’叶片GS活性较高,而‘汕G’和‘TH’叶片GS活性较低。
表3 不同生育期5个花生品种(系)叶片谷氨酰胺合成酶(GS)活性的比较
2.2.3 谷氨酸脱氢酶(GDH)活性的变化 不同生育期(花针期、结荚期和饱果期)供试的5个花生品种(系)叶片中谷氨酸脱氢酶(GDH)活性的变化见表4。结果显示:在3个生育期内,供试的5个花生品种(系)叶片GDH活性的变化趋势基本一致,均呈逐渐升高的趋势,在饱果期达最高,但不同品种(系)叶片GDH活性水平及变化幅度有一定差异。
在3个生育期内,‘泉花7号’叶片的GDH活性均最高,而‘汕E’叶片的GDH活性均最低。差异显著性分析结果显示:在花针期,仅‘泉花7号’叶片GDH活性分别与‘汕E’和‘TJ’有极显著或显著差异;在结荚期,‘泉花7号’、‘TH’ 和‘汕G’叶片GDH活性分别与‘汕E’和‘TJ’有极显著或显著差异,其中‘汕E’叶片GDH活性均极显著低于其他4个品种(系);而在饱果期,‘汕E’叶片GDH活性也均极显著低于其他4个品种(系)。
总体上看,5个花生品种(系)中,‘泉花7号’、‘汕G’和‘TH’叶片GDH活性较高,而‘汕E’叶片GDH活性最低。
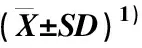
表4 不同生育期5个花生品种(系)叶片谷氨酸脱氢酶(GDH)活性的比较
2.3 不同品种(系)单株荚果和单株秆产量的比较
以单株荚果鲜质量和单株秆鲜质量作为供试5个花生品种(系)的产量指标,收获期2个产量指标的测定结果见表5。在供试的5个花生品种(系)中,‘汕G’的单株荚果鲜质量最高,分别较‘泉花7号’、‘TH’、‘汕E’和‘TJ’高1.76%、6.18%、17.65%和22.65%;按照单株荚果鲜质量从高至低排序,5个品种(系)依次为‘汕G’、‘泉花7号’、‘TH’、‘汕E’、‘TJ’,其中,‘汕G’和‘泉花7号’的单株荚果鲜质量无显著差异,但与‘汕E’和‘TJ’有极显著差异。5个品种(系)中,‘TH’的单株秆鲜质量最高,分别较‘泉花7号’、 ‘汕G’、 ‘TJ’和‘汕E’高19.30%、 19.80%、22.67%和32.63%;按照单株秆鲜质量从高至低排序,5个品种(系)依次为‘TH’、‘泉花7号’、‘汕G’、‘TJ’、‘汕E’,其中,‘TH’和‘汕E’的单株秆鲜质量分别极显著高于或低于其他品种(系)。
表5 不同花生品种(系)单株荚果鲜质量和单株秆鲜质量的比较
总体上看,‘汕G’的荚果和秆产量均较高,其次为‘泉花7号’,而‘汕E’的荚果和秆产量均较低。
3 讨论和结论
植物叶片可溶性蛋白质是多种酶系构成的非膜结合蛋白体系,反映了叶片酶蛋白功能的变化,其含量代表叶片氮代谢水平和叶片生活力的高低,也是反映叶片衰老的指标之一。上述实验结果表明:供试的5个花生品种(系)叶片可溶性蛋白质含量在结荚期均最高、饱果期降低,表明在结荚期花生叶片蛋白质合成能力较高,可以为花生结实提供氮储备;饱果期叶片可溶性蛋白质含量较低则表明植株进入衰老进程。这一现象与张智猛等[12]、李向东等[13]和王才斌等[14]获得的研究结果基本一致。
硝酸还原酶(NR)是NO3--N同化的限速酶,是植物氮代谢的关键酶;叶片中NR活性的高低控制着整个同化过程,其强弱在一定程度上反映了光合作用、呼吸作用及蛋白质合成的强度和氮代谢活性[1];此外,NR活性与土壤含氮量和施肥量也有密切关系,土壤含氮量和施肥量越高、植株体内的NR活性也越强,其氮素代谢也越旺盛[21]。在花针期、结荚期和饱果期,虽然供试的5个花生品种(系)的NR活性有一定差异,但均以花针期最高、并在结荚期有所下降,表明花针期植株体内的氮代谢均较为旺盛;但在饱果期,‘泉花7号’的NR活性却明显高于结荚期,说明在生育后期‘泉花7号’叶片的氮代谢还维持在比较高的水平。
谷氨酰胺合成酶(GS)参与多种氮代谢的调节,是氮代谢中心的多功能酶[17]。在不同生育期,供试的5个花生品种(系)叶片的GS活性有不同的变化趋势,其中,‘汕G’和‘TH’叶片GS活性在结荚期高于花针期及饱果期,表明结荚期这2个品种(系)的氨同化能力较高,而这2个品种(系)的单株荚果鲜质量也均较高,与此现象可能有一定的关系;‘汕E’、‘TJ’和‘泉花7号’叶片GS活性均表现为花针期高于结荚期和饱果期,可能与土壤基础肥力过低导致的花生叶片提早衰老有关。在花针期、结荚期和饱果期,供试的5个花生品种(系)叶片的GDH活性均逐渐升高,其中饱果期与结荚期叶片的GDH活性差异较小,表明花生叶片GDH活性的变化趋势与GS活性的变化趋势不完全一致。
在红壤条件下,不同生育期供试的5个花生品种(系)叶片可溶性蛋白质含量以及NR和GDH活性的变化趋势基本一致,但其水平有一定差异。而花生荚果产量与叶片氮代谢相关酶活性有一定的相关性,荚果产量较高的‘汕G’和‘泉花7号’,其叶片NR和GDH活性均相对较高;而荚果产量最低的‘TJ’,其叶片的NR和GDH活性也均较低。但花生荚果的产量和质量并不取决于单一或某一生理因子,而是外部环境与内在遗传及生理代谢因子共同作用的结果。因此,在花生的生长发育过程中应采取不同的管理栽培措施,提高各器官氮代谢的生理机能,延缓衰老,增强光合效率,达到增产的目的。
参考文献:
[1] 孙大容. 花生育种学[M]. 北京: 中国农业出版社, 1998.
[2] 福建省统计局. 福建经济与社会统计年鉴: 农村篇[M]. 福州: 福建人民出版社, 2004.
[3] 柴小清, 印莉萍, 刘祥林, 等. 不同浓度的NO3-和NH4+对小麦根谷氨酰胺合成酶及其相关酶活性的影响[J]. 植物学报, 1996, 38(10): 803-806.
[4] 王月福, 于振文, 李尚霞, 等. 氮素营养水平对冬小麦氮代谢关键酶活性变化和籽粒蛋白质含量的影响[J]. 作物学报, 2002, 28(6): 743-748.
[5] 王小纯, 熊淑萍, 马新明, 等. 不同形态氮素对专用型小麦花后氮代谢关键酶活性及籽粒蛋白质含量的影响[J]. 生态学报, 2005, 25(4): 802-807.
[6] 张智猛, 戴良香, 胡昌浩, 等. 氮素对不同类型玉米蛋白质及其组分和相关酶活性的影响[J]. 植物营养与肥料学报, 2005, 11(3): 320-326.
[7] 王宪泽, 田纪春, 张忠义. 不同品种小麦籽粒蛋白质及其组分积累规律的研究[J]. 山东农业大学学报, 1994, 25(4): 394-398.
[8] 赵俊晔, 于振文. 施氮量对小麦强势和弱势籽粒氮素代谢及蛋白质合成的影响[J]. 中国农业科学, 2005, 38(8): 1547-1554.
[9] 刘开昌, 胡昌浩, 董树亭, 等. 高油、高淀粉玉米籽粒主要品质成分积累及其生理生化特性[J]. 作物学报, 2002, 28(4): 492-498.
[10] 曹 云, 范晓荣, 贾莉君, 等. 不同水稻品种对NO3-同化差异的比较[J]. 南京农业大学学报, 2005, 28(1): 52-56.
[11] 朱德群, 朱遐龄, 王 雁, 等. 与冬小麦籽粒蛋白质有关的几项生理参数[J]. 作物学报, 1991, 17(2): 135-144.
[12] 张智猛, 万书波, 宁堂原, 等. 不同花生品种氮代谢相关酶活性的研究[J]. 植物营养与肥料学报, 2007, 13(4): 707-713.
[13] 李向东, 王晓云, 张高英, 等. 花生衰老进程的研究[J]. 西北植物学报, 2001, 21(6): 1169-1175.
[14] 王才斌, 孙彦浩, 牛振荣, 等. 高产花生叶片和荚果碳氮化合物代谢规律的研究[J]. 莱阳农学院学报, 1996, 13(2): 99-104.
[15] 姜慧芳, 任小平. 干旱胁迫对花生叶片SOD活性和蛋白质的影响[J]. 作物学报, 2004, 30(2): 169-174.
[16] 李尚霞, 封海胜, 宫清轩, 等. 花生不同类型品种根系生育特性研究[J]. 花生学报, 2005, 34(3): 26-29.
[17] 杜应琼, 廖新荣, 黄志尧, 等. 硼、钼对花生氮代谢的影响[J]. 作物学报, 2001, 27(5): 612-616.
[18] 周录英. 不同肥料运筹对花生生理性状及产量品质的影响[D]. 泰安: 山东农业大学农学院, 2006.
[19] 李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000: 125-128.
[20] 上海植物生理学会. 植物生理学实验手册[M]. 上海: 上海科学技术出版社, 1985: 223-226.
[21] 李 琪. 水分-氮素对玉米苗期生长及氮代谢的影响[D]. 武汉: 华中农业大学资源与环境学院, 2007.
