南岭大东山长柄双花木群落物种组成与种群结构特征分析
2014-04-09缪绅裕曾庆昌陈志明刘嘉俊李晓杰黄金玲
缪绅裕, 曾庆昌, 陈志明, 刘嘉俊, 李晓杰, 黄金玲
(1. 广州大学: a. 生命科学学院, b. 建筑与城市规划学院, 广东 广州 510006;2. 广东南岭国家级自然保护区大东山管理处, 广东 连州 513400)
金缕梅科(Hamamelidaceae)双花木属(DisanthusMaxim.)种类长柄双花木(Disanthuscercidifoliusvar.longipesChang)为中国特有植物,于1999年被列为国家Ⅱ级重点保护野生植物,散生分布于江西、湖南、浙江和广东北部的少数地区[1-3],广东南岭大东山潘家洞地段是迄今为止发现的长柄双花木种群自然分布的最南界限。目前,对江西吉安井冈山、玉山三清山、宜春官山和南丰军峰山,湖南道县空树岩和千家洞、常宁阳山和宜章莽山,浙江龙泉和开化等地分布的长柄双花木群落的相关研究报道较多[1-5]; 而对广东南岭大东山长柄双花木群落植物区系和资源现状以及种群特征仅有少量研究报道[6-7]。目前,针对长柄双花木的研究主要涉及群落组成和物种多样性、种群结构和动态、种子休眠和萌发、繁育体系、扦插繁殖和遗传多样性等方面[2-16]。
肖宜安等[1]认为:影响井冈山长柄双花木生长的潜在主导因子之一是海拔高度;缪绅裕等[7]的研究结果也表明:广东南岭大东山长柄双花木种群的个体密度、乔木层个体平均高度和地径均受海拔高度的影响;但沈如江等[3]认为:海拔高度对不同区域(江西三清山和官山以及湖南千家洞)分布的长柄双花木群落的影响并不明显。因此,海拔高度对长柄双花木生长及分布的生态作用可能因分布地不同以及其自身形态的不同而存在一定的差异。
作者以广东南岭大东山不同海拔的长柄双花木自然种群为研究对象,对其群落物种组成及种群年龄结构、静态生命表、生存曲线和生殖力特征进行比较分析,以期为南岭大东山长柄双花木资源的保护和可持续发展提供科学依据。
1 研究区概况和研究方法
1.1 研究区概况
广东省境内的长柄双花木自然种群仅分布于南岭大东山潘家洞山体,地理坐标为北纬24°55′11″、东经112°39′33″,海拔约700~1 300 m;相对集中分布面积约2.5 hm2,初步统计长柄双花木有1万株以上;该区域毗邻南岭国家级自然保护区大东山管理处。大东山地处南岭山地南侧、北接湖南宜章莽山,属典型的中亚热带季风气候;年均气温15.5 ℃,最冷月1月均温8.9 ℃,最热月7月均温28.5 ℃,≥10 ℃年均有效积温6 236.5 ℃; 年降雨量1 637.6 mm, 集中在 3月份至8月份;盛吹东南风。大部分为中山山地,最低海拔650 m,山峰海拔多在1 000 m以上,最高海拔1 597 m。主要山体由中生代花岗岩侵入体构成,海拔800~1 150 m的山地土壤为黄壤,海拔1 150 m以上的山顶为山地灌丛草甸土[17]。
1.2 研究方法
在踏查该区域长柄双花木资源量的基础上,于2012年7月(距2008年初南岭地区遭受严重冰冻灾害约4.5 a)选择3个不同海拔的群落作为样带,其中,小洞水库边坡地海拔1 010 m、大洞水库边坡地海拔1 170 m、鸡子场海拔1 260 m,样带选择主要根据自然种群集中分布情况设定(无法按海拔高差相等设定样带);每个样带面积10 m×100 m,各自划分为10个面积10 m×10 m的小样方[7]。
长柄双花木基部多分枝,为丛生灌木或小乔木,故其构件以株为单位,用游标卡尺测定个体的基径和高度;DBH≥5.0 cm的长柄双花木个体计入乔木层;对乔木层中DBH≥5.0 cm的所有种类的个体进行每木调查(记录胸径和树高等),根据重要值计算公式[18]29计算乔木层各树种的重要值。
长柄双花木为国家重点保护野生植物,因而不能采用伐木或钻木法测定其个体年龄,只能根据长柄双花木的基径推测其年龄。假设长柄双花木每年的生长速率均等,从基径1.0 mm开始(所测基径最小为1.08 mm),基径1.5 mm为1个龄级(相当于1 a),最大基径101.72 mm,据此计算出大东山长柄双花木个体最大年龄约68 a,接近井冈山长柄双花木种群的最大年龄(65 a)[8],并由此绘制出种群径级结构图(替代年龄结构图)。 “以空间代替时间”编制大东山长柄双花木种群静态生命表[8];为便于比较,编制静态生命表时把3个种群数据汇总计算,并以5 a为1个龄级,具体计算公式和方法参见文献[9]。
2 结果和分析
2.1 群落乔木层的种类组成
南岭大东山3个样带长柄双花木群落乔木层主要种类(DBH≥5.0 cm)的重要值见表1。由表1可见:在海拔较低(1 010 m)的小洞样带长柄双花木的优势明显大于海拔更高的大洞(1 170 m)和鸡子场(1 260 m)样带;而且小洞样带乔木层仅有11个种,种类数相对较少,与长柄双花木的优势较大有关;其中,部分地段长柄双花木生长过于密集,导致林下几乎没有草本植物。

表1 南岭大东山不同样带长柄双花木群落乔木层种类的重要值
黄山松(PinustaiwanensisHayata)与马尾松(P.massonianaLamb.)在不同样带中有替代分布现象,其中,海拔较低的小洞样带中未见黄山松而仅有马尾松分布;海拔较高的大洞样带中既分布有马尾松也分布有黄山松,而3个样带中海拔最高的鸡子场样带仅见黄山松而无马尾松分布。3个样带的乔木层共出现26个种类,其中的共有种除长柄双花木外,仅有多花杜鹃(RhododendroncavalerieiLévl.)和两广杨桐(AdinandraglischrolomaHand.-Mazz.)2种;而小洞和大洞2个样带的共有种有8种;大洞和鸡子场2个样带的共用种有6种,小洞和鸡子场2个样带的共有种仅3种。从重要值看,小洞和大洞样带中重要值位居前2位的种类均为长柄双花木和马尾松,鸡子场样带中重要值位居前2位的种类是黄山松和长柄双花木;3个样带总重要值最大的是长柄双花木,而黄山松和马尾松的总重要值也较大。
2.2 种群龄级结构分析
在小洞、大洞和鸡子场3个样带中长柄双花木个体数分别为806、504和464株(合计1 774株),随海拔上升,种群个体数和密度呈下降趋势;根据长柄双花木每木基径的测定结果绘制的小洞、大洞和鸡子场3个样带长柄双花木的龄级结构见图1。3个样带长柄双花木种群的龄级结构基本一致,由此推测整个山体长柄双花木种群应在同一时期产生。海拔较低的小洞样带(与大洞样带海拔相差160 m)种群规模(个体数量)更大,尤其是基径31.0 mm(相当于株龄20 a)以下的幼体(长柄双花木在生长20 a后才进入生殖期[9])所占比例最大;海拔较高的大洞样带和鸡子场样带长柄双花木种群的径级结构则更为一致,可能与这2个样带的海拔高度差(相差90 m)较小有关。由此可见:海拔高度对长柄双花木种群的发育有一定影响。

―●―: 小洞样带(海拔1 010 m) Xiaodong sampling transect with altitude 1 010 m;
整个南岭大东山的长柄双花木种群以基径8.6~25.0 mm的个体(相当于株龄6~16 a)为主,均处于生殖前期,而基径8.5 mm以下(相当于株龄5 a以内)的幼苗数量偏低,这一现象可能与2008年初南岭地区遭受的冰冻灾害有较密切的关系;其他龄级的个体数量呈现波动但整体随株龄增大而下降的趋势。从图1可见:南岭大东山长柄双花木种群属于基本稳定型,但因目前株龄5 a以内的幼苗数量明显偏少,预测50~60 a后该种群将进入衰退状态。种群中基径77.6 mm(相当于株龄52 a)及以上的个体所占比例很低,并且从该龄级后出现某些龄级个体数为0的现象,说明该种群在经历了约50 a的生长期后将进入死亡高峰阶段。
2.3 种群静态生命表
生命表是同龄群个体在各特定年龄阶段的生存和死亡概率的总结,不仅可反映种群从出生到死亡的数量动态,还可用于预测种群未来发展的趋势[18]39。根据野外实测数据和公式编制的南岭大东山长柄双花木种群的静态生命表见表2。龄级5~15 a的长柄双花木个体死亡率相对较高,5~10 a和10~15 a龄级的个体死亡率分别为20.30%和20.50%;15~20 a龄级的个体死亡率也达到13.90%,之后随龄级增加个体死亡率均在10.00%以下,总体上呈现随龄级增加而逐渐下降的趋势,这可能与南岭大东山长柄双花木种群在2008年遭受冰冻灾害有关。此外,由表2还可见:不同龄级的长柄双花木个体平均生命期望值(ex)为4.49~0.50,基本呈现出随个体年龄增长而下降的趋势。种群生命期望值总体偏低,可能与南岭大东山长柄双花木种群的生境条件(例如光照等环境因素)有关。
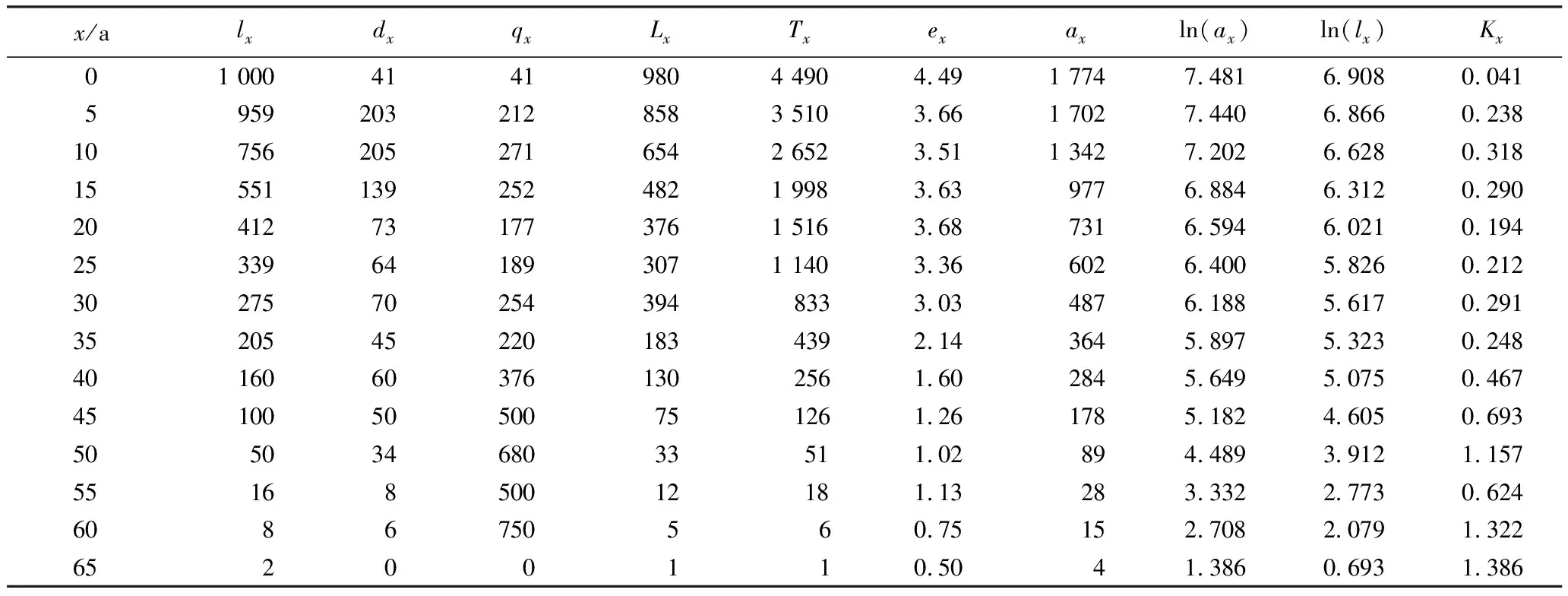
表2 南岭大东山长柄双花木种群的静态生命表1)
2.4 种群存活曲线
为更直观了解南岭大东山长柄双花木种群的死亡过程,以表2中lx的对数值ln(lx)为纵坐标、长柄双花木的年龄间隔为横坐标[18]50,绘制南岭大东山长柄双花木种群存活曲线,结果见图2。南岭大东山长柄双花木种群的存活曲线为较典型的Ⅰ型(凸型),即种群在接近生理寿命之前死亡率相对较低(特别是25~40 a年龄段),直到生命后期(约50 a后)死亡率增加较快,这与静态生命表(表2)的分析结果一致。
2.5 种群生殖力分析
就植物种群而言,特定年龄产生种子的数目为生殖力,而反映产生种子的数量动态过程称为生殖力表或生育力表[18]43。南岭大东山长柄双花木种群的生殖力表见表3。根据表3中的数据,计算出该种群的净增殖率R0=0.504,说明长柄双花木每一世代将以0.504倍的数量增加;而R0<1,则表明该种群难以自我更新。内禀增长率rm=-0.018,rm值较低预示着种群在自然条件下可能出现低死亡率。周限增长率λ=0.982,表明该种群规模将以0.982倍的速度下降。世代平均周期T=38.351 a,也即上一代到下一代所需的平均年限较长,对于种群更新较为不利。前述这些参数均表明:南岭大东山长柄双花木种群在当前的环境状况下(经历2008年初南岭地区冰冻灾害后)自我更新不良,种群将呈现负增长的趋势,但其负增长的速率相对较缓,属于缓慢的负增长型种群。
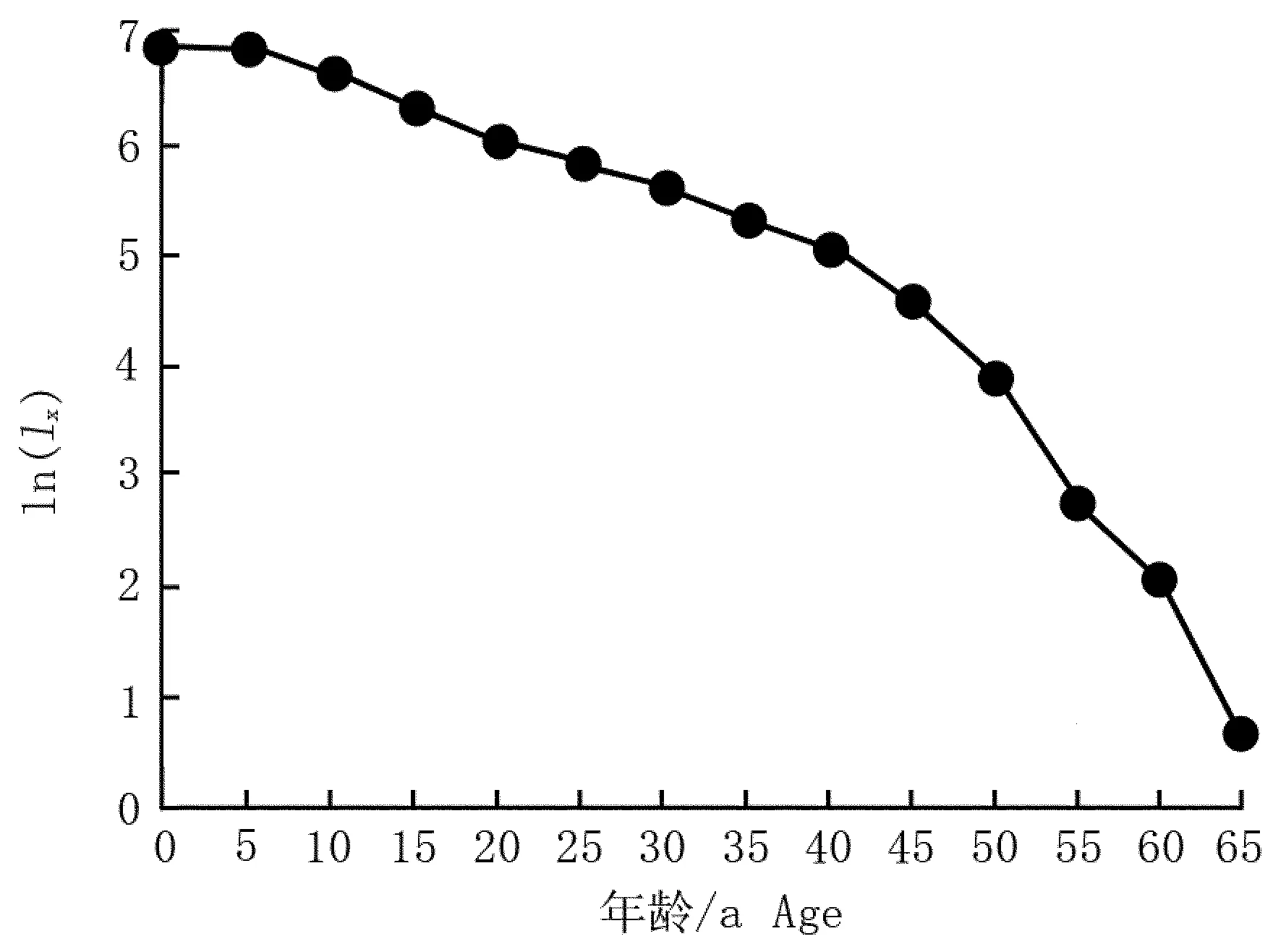
图2 南岭大东山长柄双花木种群生存曲线
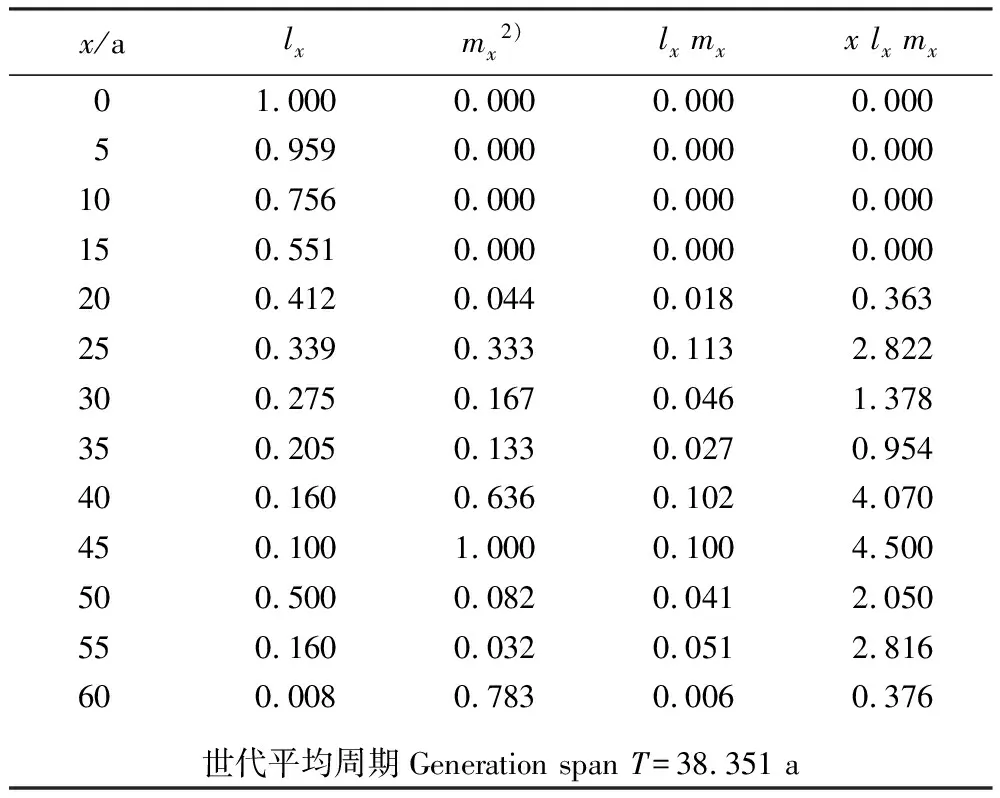
表3 南岭大东山长柄双花木种群生殖力表1)
3 讨论和结论
3.1 种群特征综合分析
在南岭大东山长柄双花木群落中,不同植物种类出现在海拔高度不同的样带中,或同一植物在不同样带中的重要值不同,说明海拔高度不同的长柄双花木群落的物种组成有一定差异。3个样带的长柄双花木种群的龄级结构也有一定区别,表明海拔高度对群落中不同植物的生存和发展有一定的影响。
由于长柄双花木为国家Ⅱ级重点保护野生植物,因而本研究未采用断木法测定其真实年龄而是根据基径推测其年龄,与种群的实际龄级存在一定的偏差。基于“长柄双花木每年生长速率相同”的假设,南岭大东山长柄双花木种群为成熟种群(各龄级个体均有分布[18]10)。通常情况下,常根据树高(H)和胸径(DBH)划分木本植物的径级结构,划分标准为:Ⅰ级幼苗(H≤33 cm)、Ⅱ级幼苗(H>33 cm,DBH<2.5 cm)、Ⅲ级小树(DBH 2.5~7.4 cm)、Ⅳ级中树(DBH 7.5~22.4 cm)、Ⅴ级大树(DBH≥22.5 cm)[18]12,但这种划分法显然不适用于作为灌木或小乔木的长柄双花木。本研究中,长柄双花木的实测胸径小于9 cm,但是若按“H≤33 cm”的标准划分Ⅰ级幼苗,则小洞样带有14株(基径0.429~0.920 cm)、大洞样带有25株(基径0.108~0.793 cm)、鸡子场样带有10株(基径0.216~0.760 cm),可见南岭大东山长柄双花木种群的幼苗数量与其他龄级相比仍处于较低水平。
在大东山长柄双花木群落中,基径在8.5 mm以内(对应株龄5 a)的幼苗或幼树个体数量很少,这主要是因为该种群在2008年遭受冰冻灾害的影响,导致该种群从原先的增长趋势转变为目前的稳定型,也说明该种群的基径结构与龄级结构有较好的对应性。若非遭受冰冻灾害,该种群似乎是典型的增长型种群。根据图1推测:若无特殊气候事件影响,该长柄双花木种群将会在2058年前后进入典型的衰退期,因此加强对该种群的保护将有可能延缓种群衰退或使种群恢复至受灾前的增长状态。
大东山潘家洞是长柄双花木自然分布的南界,因而,该种群在生态学研究中具有独特的地位。从图1结果看:该种群规模较为稳定,与江西井冈山的长柄双花木衰退种群(呈典型的倒金字塔型[7])不同,其原因可能是不同分布地长柄双花木种群的发生时期及气候变化有异,或人为破坏状况各不相同。对井冈山长柄双花木种群的调查是在10 a前,当时该种群已出现衰退趋势,故推测井冈山长柄双花木种群的发育早于大东山长柄双花木种群。井冈山长柄双花木种群1~5 a龄级的个体死亡率较高[9],而大东山长柄双花木种群5~15 a龄级的个体死亡率相对较高(见表2)、1~5 a龄级的个体死亡率较低,可能因为2008年的冰冻灾害导致部分个体死亡,而本次调查的均为幸存植株,个体死亡率不会太高。从种群的生存曲线看,大东山长柄双花木种群的存活曲线为较典型的凸型,预计在种群生存50 a后死亡率增加较快,这一结果与“井冈山长柄双花木种群50 a龄级时出现又一个死亡高峰[9]”一致。以相同龄级个体平均生产的子代数目(mx)进行比较,长柄双花木的大东山种群和井冈山种群的各增长率指标和世代平均周期均较接近,且大东山种群的所有指数均较低,两地的长柄双花木种群都属于缓慢的负增长型种群。
3.2 种群的保护策略
大东山潘家洞区域与广东南岭国家级自然保护区大东山管理处的实验区毗邻,长柄双花木在该山体海拔700 m以上有零星分布,在海拔1 000~1 300 m范围内基本成片生长,总数超过1万株。由于该种群的发现时间晚于保护区的成立时间(1994年),因而并未纳入保护区范围。该区域目前交通较为闭塞,仅有1条简易的上山林区公路,人为干扰程度相对较低,保护管理上也相对容易。为有效保护这一资源,应尽早把这一区域划入保护区范围。
因建设小洞水库和大洞水库,20世纪70年代潘家洞区域的山体遭受过较严重破坏,因此导致森林郁闭度降低,无形中增加了长柄双花木的生存空间。因长柄双花木具有喜光特性(在郁闭度较高的阔叶林中数量锐减[9]),故目前在小洞水库旁和大洞水库旁的山坡生长良好,种群规模基本处于稳定状态,若非遭受冰冻灾害,其自然更新能力正常。
生境破碎化和人为干扰(村民缺乏保护意识的“樵柴”砍伐),以及自然灾害(冰灾等)和自身繁殖力低等因素是长柄双花木种群濒危状况加剧的主要原因。因此,应采取综合措施对长柄双花木自然种群进行保护,具体措施包括:1)将其纳入保护区范围,加强自然种群原生地的就地保护;2)加大宣传教育力度,强化当地民众的保护意识;3)加强相关学科研究,建立定位观测站对该自然种群进行长期监测;4)通过采种育苗及扦插繁殖对长柄双花木进行迁地保护。
参考文献:
[1] 肖宜安, 何 平, 邓洪平, 等. 井冈山长柄双花木种群形态分化的数量分析[J]. 武汉植物学研究, 2002, 20(5): 365-370.
[2] 李根有, 陈征海, 邱瑶德, 等. 浙江省长柄双花木数量分布与林学特性[J]. 浙江林学院学报, 2002, 19(1): 20-23.
[3] 沈如江, 林石狮, 凡 强, 等. 江西省三清山长柄双花木优势群落研究[J]. 武汉植物学研究, 2009, 27(5): 501-508.
[4] 李矿明, 汤晓珍. 江西官山长柄双花木灌丛的群落特征与多样性[J]. 南京林业大学学报: 自然科学版, 2003, 27(3): 73-75.
[5] 饶 军, 李荣同. 军峰山的长柄双花木[J]. 江西林业科技, 1997(4): 7-8, 14.
[6] 谢国文, 谭巨清, 曾宇鹏, 等. 国家重点保护物种长柄双花木南岭群落植物区系与资源[J]. 广东教育学院学报, 2010, 30(5): 79-87.
[7] 缪绅裕, 陈志明, 陶文琴, 等. 大东山不同海拔长柄双花木种群形态特征比较[J]. 广东农业科学, 2013, 40(12): 167-170.
[8] 肖宜安, 肖 南, 胡文海, 等. 濒危植物长柄双花木自然种群年龄结构及其生态对策[J]. 广西植物, 2007, 27(6): 850-854.
[9] 肖宜安, 何 平, 李晓红, 等. 濒危植物长柄双花木自然种群数量动态[J]. 植物生态学报, 2004, 28(2): 252-257.
[10] 史晓华, 徐本美, 黎念林, 等. 长柄双花木种子休眠与萌发的初步研究[J]. 种子, 2002(6): 5-7.
[11] 肖宜安, 曾建军, 李晓红, 等. 濒危植物长柄双花木自然种群结实的花粉和资源限制[J]. 生态学报, 2006, 26(2): 496-502.
[12] 肖宜安, 何 平, 李晓红. 濒危植物长柄双花木的花部综合特征与繁育系统[J]. 植物生态学报, 2004, 28(3): 333-340.
[13] XIAO Y A, NEOG B J, XIAO Y H, et al. Pollination biology ofDisanthuscercidifoliusvar.longipes, an endemic and endangered plant in China[J]. Biologia, 2009, 64(4): 731-736.
[14] 黄绍辉, 方炎明, 谭雪红, 等. 不同浓度NAA对长柄双花木扦插繁殖的影响[J]. 植物资源与环境学报, 2007, 16(4): 74-75.
[15] 肖宜安, 何 平, 邓洪平, 等. 井冈山长柄双花木种群遗传多样性与遗传分化[J]. 西南师范大学学报: 自然科学版, 2003, 28(3): 444-449.
[16] 谢国文, 赵俊杰, 李荣华, 等. 濒危植物长柄双花木ISSR-PCR反应体系的优化[J]. 广州大学学报: 自然科学版, 2010, 9(2): 45-50.
[17] 唐绍清, 张宏达, 唐志信, 等. 粤北大东山种子植物区系研究[J]. 广西植物, 1997, 17(2): 127-132.
[18] 王伯荪, 李鸣光, 彭少麟. 植物种群学[M]. 广州: 广东高等教育出版社, 1995.
