天目山柳杉叶水势日变化及其与空气温湿度和PAR的相关性
2014-04-09赵明水蒋文伟
秦 登, 刘 鹤, 赵明水, 蒋文伟,①
(1. 浙江农林大学园林学院, 浙江 临安 311300; 2. 浙江天目山国家级自然保护区管理局, 浙江 临安 311311)
植物的生理活动需要一定的水分环境,水势是植物水分状态的基本度量单位,也是目前最常用的水分生理指标[1]。叶水势表示植物水分运动的能量水平、反映植物组织水分状况,是衡量植物抗旱性的重要生理指标[2];叶水势越低则表明植物的吸水能力越强。Selles等[3]和Jones等[4]的研究结果表明:植物叶水势是干旱胁迫环境下最敏感的度量指标;田丽等[5]建立了不同供水条件下树木叶水势与气象因子的定量关系;魏晓霞等[6]则认为:随树龄增加树木叶水势的主要影响因子有所变化。众多的研究结果均表明植物叶水势的日变化与环境因子关系密切[7-10]。
柳杉(CryptomeriafortuneiHooibrenk ex Otto et Dietr.)系杉科(Taxodiaceae)柳杉属(CryptomeriaD. Don)常绿乔木,为中国特有种,在浙江天目山国家级自然保护区植物区系中占有重要地位。杨广远等[11]的研究结果表明: 天目山柳杉春秋季树干液流与空气温度、空气相对湿度、光合有效辐射和叶面湿度有显著相关性;夏爱梅等[12]认为:柳杉幼苗很难在天目山海拔1 000 m以上的区域自然扎根生长,柳杉古树的枯亡对天目山高海拔景观林也有重要影响。
为了解导致天目山柳杉古树枯亡及其幼苗难以生存的原因,作者对不同样点柳杉叶水势进行观测,分析不同海拔区域柳杉叶水势与空气温度、空气相对湿度和光合有效辐射的相关性,以期为天目山柳杉古树的保护及其种群抚育管理提供科学依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区域位于浙江省临安市天目山国家级自然保护区,共设3个样点:山顶老殿,地理坐标为北纬30°20′32″、东经119°26′02″,海拔1 067 m,土壤含水量20.31%;山腰五里亭,地理坐标为北纬30°20′08″、东经119°26′09″,海拔698 m,土壤含水量19.16%;山麓太子庵,地理坐标为北纬30°19′28″、东经119°26′24″,海拔419 m,土壤含水量18.25%。
研究区属中亚热带北缘向北亚热带的过渡地带,气候温和、四季分明、雨水充沛,森林气候复杂多变。年均气温8.8 ℃~14.8 ℃,年太阳辐射3 270~4 460 MJ·m-2, 年降水量1 390~1 870 mm; 最冷月平均 气温-2.6 ℃~3.4 ℃, 最热月平均气温19.9 ℃~ 28.1 ℃;无霜期209~235 d,年雨日159.2~183.1 d,年雾日64.1~255.3 d。成土母岩主要为流纹质凝灰岩;海拔600 m以下为红壤,海拔600 m以上为黄壤。
1.2 方法
1.2.1 样株选择 在上述3个样点附近的柳杉群落中分别选择3株生长健壮、树龄120~160 a且株型高大的柳杉作为样株。
1.2.2 叶水势测定 在柳杉生长旺盛期选择3个晴朗观测日(2011年8月8日至10日),选取样株向阳面不同冠层的典型针叶测定叶水势,树冠高度(测定叶片距离地面的高度)分别为1.3、4和8 m,叶片为1年生新枝上部1/3~1/2段的完整新叶。采用Psypro露点水势仪(美国WESCOR公司)观测叶水势,观测时间为9:00至17:00,每隔2 h观测1次,每次测定重复6次,结果取平均值。
1.2.3 气候因子测定 分别在样株的东、南、西、北4个方向,用Kestrel 4500手持式风速风向仪(美国KESTREL公司)测定各样点空气温度和空气相对湿度,用3415FQF光量子与照度计(美国SPECTRUM公司)测定光合有效辐射(PAR),测定时间与叶水势同步,结果取平均值。
1.3 数据处理
应用EXCEL 2003数据分析软件进行数据处理,用Sigma Plot 10软件制图,并用SPSS 11.5 统计分析软件进行相关性分析。
2 结果和分析
2.1 柳杉叶水势的日变化分析
天目山不同海拔样点柳杉不同冠层叶水势的日变化曲线见图1。由图1可知:同一样点不同高度树冠的叶水势日变化曲线均呈典型的“V”型,其原因在于早晨太阳辐射逐渐增强、空气温度随之升高且空气相对湿度降低,植物蒸腾作用加强,导致叶水势逐渐降低,至中午前后叶水势降至全天最低值。其中,海拔1 067 m处柳杉树冠下层(高1.3 m)叶水势最低值为-0.79 MPa,出现在13:00;树冠中层(高4 m)叶水势最低值为-0.89 MPa,树冠上层(高8 m)叶水势最低值为-0.98 MPa,均出现在11:00。海拔698 m处树冠下层叶水势最低值为-0.78 MPa,出现在15:00;树冠中层叶水势最低值为-0.92 MPa,出现在11:00;树冠上层叶水势最低值为-0.95 MPa,出现在13:00。海拔419 m处树冠下层、中层和上层叶水势最低值分别为-1.16、-1.22和-1.19 MPa,均出现在11:00。随时间推移,太阳辐射减弱、空气温度下降,植物蒸腾作用减弱,导致叶水势又逐渐上升。
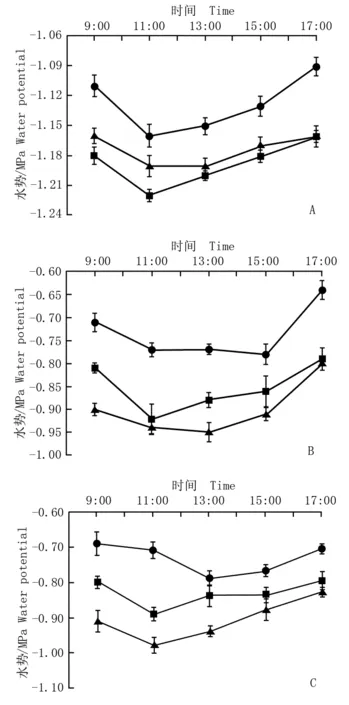
─●─: 冠层离地面高1.3 m Height of crown layer off the ground of 1.3 m;
各样点柳杉叶水势日均值从树冠下层往上层逐渐降低,海拔1 067 m处树冠下层、中层和上层的叶水势日均值分别为-0.73、-0.83和-0.91 MPa;海拔698 m处树冠下层、中层和上层的叶水势日均值分别为-0.73、-0.85和-0.90 MPa;海拔419 m处树冠下层、中层和上层的叶水势日均值分别为-1.13、-1.19和-1.17 MPa,表明随树冠增高柳杉对水分的吸取能力依次增强。
不同样地间柳杉叶水势差异明显。海拔419 m处叶水势明显低于海拔1 067和698 m处相同冠层的叶水势,而后2个样点同一冠层叶水势较为接近;海拔419 m处叶水势为-1.11~-1.22 MPa,而海拔1 067和698 m处叶水势为-0.78~-0.98 MPa。多重比较结果表明:海拔419 m处柳杉叶水势与海拔698和1 067 m处柳杉同一冠层的叶水势差异极显著(P<0.01),而后2个样点同一冠层叶水势的差异未达极显著水平(P>0.01)。
2.2 空气温度、空气相对湿度和PAR的日变化分析
天目山不同海拔样点空气温度、空气相对湿度和光合有效辐射(PAR)的日变化曲线见图2。由图2可见:在8月份,天目山不同海拔样点的空气温度日变化曲线均呈典型“单峰”型,海拔1 067、698和419 m处全天最高气温分别为26.4 ℃、29.3 ℃和32.4 ℃,均出现在13:00左右;全天最低气温分别为24.0 ℃、25.8 ℃和28.1 ℃,均出现在17:00左右。
同一样点PAR也呈典型的“单峰”型。在上午9:00,海拔1 067、698和419 m处的PAR分别为228.00、286.76和329.78 μmol·m-2·s-1;随时间推移,太阳辐射逐渐增强,各样点的PAR在13:00左右达到最高值,海拔1 067、698和419 m处的PAR分别为340.08、384.19和420.35 μmol·m-2·s-1;此后PAR逐渐减弱,至17:00左右达到最低值,海拔 1 067、698和419 m处的PAR分别为173.10、188.32和189.95 μmol·m-2·s-1。
全天空气相对湿度的变化趋势与空气温度和PAR的变化趋势明显不同,呈非典型的“U”型曲线。海拔1 067 和698 m处的空气相对湿度在9:00的初始观测值分别为79.8%和79.3%;此后随时间推移逐渐下降,于13:00降至最低值70.2%和64.5%;随后又逐渐上升,至17:00分别达到80.1%和80.7%。海拔419 m处空气相对湿度在9:00的初始观测值为71.1%,此后逐渐下降并在11:00降到最低值62.2%,然后随时间推移逐渐上升,至17:00达到77.3%。
由以上分析结果可知:天目山不同海拔柳杉样地的空气温度和PAR均随海拔升高而降低,而空气相对湿度则随海拔升高而增大。
2.3 柳杉叶水势与空气温度、空气相对湿度和PAR的相关性分析
对天目山不同海拔样点柳杉叶水势(不同冠层的叶水势平均值)日平均值与空气温度、空气相对湿度和光合有效辐射(PAR)进行相关性分析,相关系数见表1。由表1可知:不同海拔样地柳杉叶水势与空气相对湿度均呈正相关、与空气温度和PAR均呈负相关。其中,不同海拔样点柳杉叶水势与空气相对湿度的正相关性均达到显著水平(P<0.05); 但海拔 1 067 m样点柳杉叶水势与PAR呈显著负相关(P<0.05),与空气温度的相关性不显著(P>0.05);海拔419 m样点柳杉叶水势与空气温度呈显著负相关(P<0.05),与PAR的相关性未达显著水平(P>0.05);而海拔698 m样点柳杉叶水势与空气温度和PAR的相关性均未达显著水平(P>0.05)。
以叶水势为因变量(Y),以空气温度(Ta)、空气相对湿度(RH)和PAR为自变量(X)进行逐步回归分析,结果表明:在海拔1 067、698和419 m处柳杉叶水势的逐步回归方程分别为Y=-1.337+0.007XRH、Y=-1.265+0.006XRH和Y=-0.818-0.012XTa,表明在海拔1 067和698 m处空气相对湿度是影响柳杉叶水势日变化的最主要环境因子,而在海拔419 m处空气温度是影响其变化的最主要环境因子。
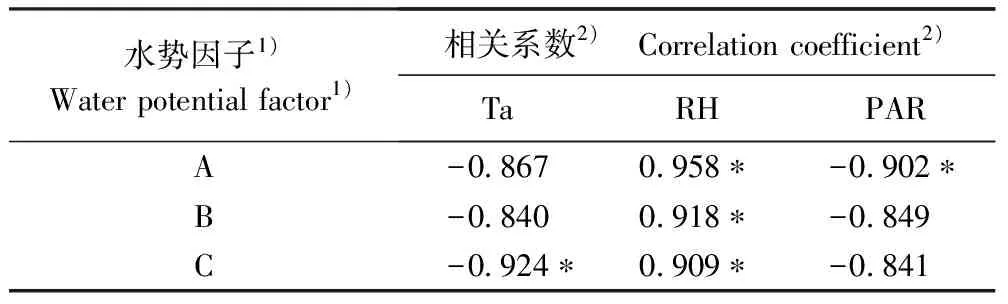
表1 天目山不同海拔样点柳杉叶水势与空气温度(Ta)、空气相对湿度(RH)和光合有效辐射(PAR)的相关系数
3 讨论和结论
罗维成等[13]认为:清晨的植物水势可以反映植物水分的恢复状况,并据此判断植物水分亏缺的程度。从上述研究结果可见:天目山不同海拔样点的柳杉叶水势日变化曲线均呈明显的“V”型,均在上午处于较高水平,说明夜晚柳杉根系吸水良好,从而使植株白天消耗的水分得到补充。
水势的高低表明植物吸水能力的强弱,而水势日变化幅度的大小也可指示植物对外界环境的敏感性强弱[14]。天目山海拔419 m处柳杉叶水势明显低于海拔1 067和698 m处,且差异极显著,说明低海拔处柳杉的吸水能力较强,根系供水能够满足植株生长代谢的需求。与海拔419 m处的柳杉叶水势相比,海拔1 067和698 m处柳杉叶水势的变化幅度更大,说明在天目山高海拔区域生长的柳杉对外界环境的敏感性更强,这也可能是天目山高海拔区域柳杉枯亡、柳杉景观林衰退的原因之一。
叶水势体现了植物水分运动的能量水平,是植物体内水分变化的直接表现,反映了植物在生长过程中各种生理活动受环境水分条件的制约程度[15]。环境因子对植物叶水势的日变化过程有重要影响。植物叶水势在全天呈现先下降后上升的趋势,这种规律与空气温度、空气相对湿度和太阳辐射日变化有密切关系。研究结果表明:天目山不同海拔样点柳杉叶水势的日变化均与空气相对湿度呈显著正相关,与空气温度和光合有效辐射则呈负相关。逐步回归分析结果显示:在海拔419 m处影响柳杉叶水势日变化的主导因子是空气温度,而在海拔1 067 和698 m处主导因子是空气相对湿度。表明随海拔升高,影响柳杉叶水势变化的主导因子有所改变。
参考文献:
[1] 庄 丽, 陈亚宁, 李卫红, 等. 渗透胁迫条件下植物茎叶水势的变化——以塔里木河下游胡杨为例[J]. 中国沙漠, 2006, 26(6): 1002-1008.
[2] 刘海涛, 贾志清, 朱雅娟, 等. 高寒沙地不同林龄乌柳的水分生理特性及叶性状[J]. 应用生态学报, 2012, 23(9): 2370-2376.
[3] SELLES G, BERGER A. Physiological indicators of plant water status as criteria for irrigation scheduling[J]. Acta Horticulturae, 1990, 278: 87-100.
[4] JONES H G, LAKSO A N, SYVERTSEN J P. Physiological control of water status in temperate and subtropical fruit trees[J]. Horti-cultural Reviews, 1985, 7: 301-304.
[5] 田 丽, 王进鑫, 庞云龙. 不同供水条件下气象因素对侧柏和刺槐叶水势的影响[J]. 西北林学院学报, 2008, 23(3): 25-28.
[6] 魏晓霞, 呼和牧仁, 周 梅, 等. 不同年龄华北落叶松叶水势及其影响因素的研究[J]. 干旱区资源与环境, 2010, 24(7): 144-147.
[7] 张 鸣, 张仁陟, 蔡立群. 不同耕作措施下春小麦和豌豆叶水势变化及其与环境因子的关系[J]. 应用生态学报, 2008, 19(7): 1467-1474.
[8] 韩 磊, 贺康宁, 芦新建, 等. 青海高寒半干旱区蒙古莸叶水势变化及其与环境因素的关系[J]. 水土保持通报, 2008, 28(6): 1-5.
[9] 袁国富, 庄 伟, 罗 毅. 冬小麦叶片气孔导度模型水分响应函数的参数化[J]. 植物生态学报, 2012, 36(5): 463-470.
[10] 刘 静, 贺康宁, 韩 磊. 黄土高原西部高寒区引进树种银水牛果叶水势研究[J]. 水土保持研究, 2009, 16(5): 194-198.
[11] 杨广远, 张 涛, 戴 锋, 等. 天目山柳杉树干液流春秋季特征及影响因素分析[J]. 西北林学院学报, 2013, 28(2): 8-14.
[12] 夏爱梅, 达良俊, 朱虹霞, 等. 天目山柳杉群落结构及其更新类型[J]. 浙江林学院学报, 2004, 21(1): 44-50.
[13] 罗维成, 曾凡江, 刘 波, 等. 疏叶骆驼刺母株与子株间的水分整合[J]. 植物生态学报, 2013, 37(2): 164-172.
[14] 杨吉华, 张永涛, 王贵霞, 等. 栾树、黄连木、黄栌水分生理生态特性的研究[J]. 水土保持学报, 2002, 16(4): 152-154, 158.
[15] 王华田, 马履一, 徐军亮. 油松人工林SPAC水势梯度时空变化规律及其对边材液流传输的影响[J]. 植物生态学报, 2004, 28(5): 637-643.
