金黄色葡萄球菌isdA基因对宿主上皮细胞免疫指标的影响
2014-04-09丛立新
丛立新,李 蕾,于 录
金黄色葡萄球菌(Staphylococcusaureus,简称金葡菌)是引起化脓性感染的重要人畜共患致病菌,能引发表皮感染,甚至心内膜炎、感染性皮炎、奶牛乳房炎及多种感染性疾病。自1960年首次发现耐甲氧西林金葡菌(methicillin-resistantStaphylococcusaureus,MR-SA)以来,其耐药性迅速蔓延。细菌耐药性成为治疗金葡菌感染亟待解决的问题。脊椎动物含有丰富的具有血红素的血红蛋白,在感染过程中,金葡菌分泌溶血素,导致红细胞破裂而释放血红蛋白,优先地夺取和使用亚铁血红素上的铁,能够在体外利用完整的红细胞作为唯一的铁源,有了这些铁,金葡菌可在宿主体内定居并引起发病[1-2]。一些研究表明,铁表面调节因子IsdA(iron surface adjustment factors A)不仅仅可以与宿主血红蛋白和血红素相互作用,还可以粘附许多血清中或细胞外基质蛋白和人类角层细胞包膜蛋白[3]。此外,IsdA能降低菌体细胞的疏水性,使它们能够抵抗人体皮肤脂肪酸和肽的杀菌作用,对金葡菌在人体皮肤上的生存起到关键性作用[4]。本研究通过敲除毒力因子IsdA的编码基因,阻断金葡菌的侵袭,为日后金葡菌感染机制的研究奠定了基础,同时为以后抗金葡菌制剂及疫苗的研究开辟了新的途径。
1 材料与方法
1.1实验材料 人角质化上皮细胞Hacat购自ATCC(American Type Culture Collection),本室常规传代。S.aureusNewman购自中国菌种保藏中心。敲除金葡菌野生株 Newman 的 isdA 基因,通过同源重组获得除 isdA 基因以外相同遗传背景的突变株与野生株,构建NewmanΔIsdA。
1.2主要试剂和实验仪器 RPMI-1640 培养基购自 Gibco 公司;胎牛血清购自 HyClone 公司;胰蛋白酶购自 Sigma 公司; Cell Counting Kit-8(CCK-8)购自日本同仁化学研究所; PrimeScript®RT reagent Kit和SYBR®PremixEx Taq(ROXⅡ)购自宝生物工程有限公司(TaKaRa);DNase I 购自 Fermentas公司; ECL 化学发光试剂盒购自碧云天生物技术研究所;Polyvinyliden edifluoride (PVDF)膜购自 Amersham 公司;鼠抗人 Dectin-1 抗体购自 R&D 公司、人源单抗 LL-37 购自Santa cruz 公司、鼠抗人 TLR2 和 TLR4 抗体购自 eBioscience 公司、鼠源单抗 β-actin和辣根过氧化物酶标记羊抗鼠和羊抗兔IgG购自中杉金桥公司;人IL-8的ELISA检测试剂盒购自 Biolegend 公司;人 IL-6、TNF-α、MCP-1 的 ELISA 检测试剂盒购自 eBioscience 公司;彩色预染蛋白 marker(7-175kDa)购自 NEB 公司。
二氧化碳细胞培养箱(Thermo),PCR扩增仪及梯度扩增仪(Biometra),黑马凝胶成像分析系统(上海天能科技有限公司),垂直板电泳槽、湿法转膜槽及电泳仪(BioRad 公司),水平摇床(鼎国生物公司),酶标仪(芬兰雷勃),倒置显微镜(上海万衡精密仪器有限公司),恒温循环水浴锅(日本EYELA),实验室碎花冰制冰机(日本 SANYO),7500型荧光定量PCR仪(Applied Biosystems 公司),化学发光呈像系统(DNR 公司)等。
1.3实验方法
1.3.1人Hacat上皮细胞的复苏及培养 取出液氮罐中冻存的人Hacat上皮细胞,立刻置于37 °C的恒温水浴锅中,迅速摇动,当细胞完全解冻后放入水平离心机中,1 000 r/min,离心7 min,弃上清,用含有100 U/mL青霉素、100 U/mL链霉素、1%丙酮酸钠、2 mmol/L谷氨酰胺、10%灭活胎牛血清的RPMI1640培养基重悬细胞,转移至25 mL细胞培养瓶中,在37 °C、5% CO2、饱和湿度的二氧化碳培养箱中培养,24 h后经倒置显微镜观察细胞形态,确定生长状态良好后使用预热的PBS清洗两遍,更换预热的细胞培养液,继续培养。采用胰酶和EDTA混合的消化法,将状态良好的细胞进行传代。
1.3.2ELISA法检测Newman及NewmanΔIsdA对上皮细胞趋化因子和细胞因子的影响 将人Hacat上皮细胞按每孔1×105个细胞接种于24孔培养板中。待细胞贴壁后换液,每孔加入含有5×106个细菌(50倍感染)的新鲜培养基300 μL,相同体积的培养基作空白对照,分别作用3 h、12 h和24 h后收集细胞上清,8 000 r/min离心20 min,去除沉淀后,吸出上清分别用人IL-8、IL-6、TNF-α和MCP-1的ELISA试剂盒检测细胞因子的表达水平,按照说明书操作。
1.3.3实时荧光定量PCR分析Newman及NewmanΔIsdA对上皮细胞炎性细胞因子、趋化因子、抗菌肽和相关受体mRNA表达水平的影响 将人Hacat上皮细胞按每瓶2×106个细胞接种于25 cm2培养瓶中。待细胞贴壁后换液,每孔加入含有1×108个细菌(50倍感染)的新鲜培养基5mL,相同体积的培养基作空白对照,分别作用3 h、6 h、12 h和24 h后,提取总RNA进行实时荧光定量PCR分析。按照PrimeScript®RT reagent Kit说明书进行反转录。根据GenBank数据库中发表的人IL-6、IL-8、TNF-a、MCP-1、Dectin-1、LL-37、TLR2、TLR4和β-actin的mRNA基因序列,采用Oligo6生物软件设计Real-time PCR特异引物,并由上海生工合成见表1。
反应程序:95 ℃ 10 min预变性;95 ℃ 15 s,72 ℃ 30 s;退火阶段结束开始收集荧光,40个循环。溶解度曲线程序设置:95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s。各个因子的退火温度50~60℃之间不等(表2)。

表1 基因、引物序列和扩增长度

表2 退火温度
1.3.4Western blot检测 将人Hacat上皮细胞按每瓶2×106个细胞接种于25 cm2培养瓶中。待细胞贴壁后换液,向细胞中分别每瓶加入含有1×108个Newman和NewmanΔIsdA(50倍感染)的新鲜培养基5 mL,设正常培养的Hacat上皮细胞为阴性对照,作用3 h。提取细胞总蛋白质并按照BCA蛋白浓度测定试剂盒说明书测定蛋白质浓度。将提取的蛋白质进行SDS-PAGE、转膜、免疫反应及ECL显色发光等操作进行Western blot检测。
1.4统计学分析 采用GraphPad Prism 5[5]统计软件以及文献[6]中2-△△Ct法进行数据分析进行统计学分析,数据结果为平均值±标准差,P< 0.05时,平均值间差异被记作显著性差异。
2 结 果
2.1金葡菌newman IsdA+/-对人Hacat上皮细胞炎性细胞因子分泌的效果
2.1.1IsdA基因敲除后下调了Hacat上皮细胞分泌促炎性细胞因子TNF-α,IL-6和IL-8的表达水平 如图1所示,与金葡菌Newman组相比,金葡菌缺失IsdA后,细胞的促炎性细胞因子TNF-α,IL-6和IL-8的分泌水平下降;但与空白未刺激组相比,Newman及NewmanΔIsdA均可以促进细胞的促炎性细胞因子TNF-α,IL-6和IL-8(*P<0.05, **P<0.01)的释放分泌;其中IL-6和IL-8的变化较为明显,在3~12 h之间抑制最显著。
2.1.2IsdA基因敲除后抑制了Hacat上皮细胞单核细胞趋化蛋白MCP-1的分泌 收集金葡菌Newman及NewmanΔIsdA处理组的培养上清,ELISA方法检测细胞趋化因子MCP-1的分泌情况。金葡菌缺失IsdA基因后,Hacat上皮细胞单核细胞趋化蛋白MCP-1的分泌明显抑制,而在24 h最为明显。

图1 IsdA基因缺失后对Hacat细胞促炎性细胞因子分泌的影响
2.2金葡菌Newman IsdA+/-对Hacat细胞相关受体及细胞因子mRNA转录水平的影响 采用实时荧光定量 PCR 的方法,检测金葡菌 isdA+/-对 hacat 细胞 TLR2、TLR4、Dectin-1、LL-37 mRNA转录水平的变化和促炎性细胞因子 IL-6、IL-8 和 TNF-α 以及趋化因子 MCP-1 相应的 mRNA 表达量的变化分析了这7个基因在 3 h、6 h、12 h 和 24 h 的表达情况(如图2-4)。
金葡菌IsdA+/-处理Hacat细胞后,与对照组相比,促炎症因子IL-6、IL-8和TNF-α的mRNA表达量在刚加入菌液的3 h达到高峰,随着时间的延长而逐渐降低,而缺失IsdA后,3 h到12 h之间表达水平均明显下降(*P<0.05, **P<0.01),IL-6和TNF-α的mRNA在24h变化无明显差异(P>0.05),IL-8的表达水平却极显著下降(**P<0.01)。金葡菌IsdA+/-均可使细胞的MCP-1 mRNA转录水平上调(**P<0.01),在3 h达到高峰。IsdA缺失后,3 h~12 h之间其表达均明显抑制,并在3 h抑制最明显;在24 h,IsdA缺失前后MCP-1的表达无明显差异(P>0.05)(图2)。 TLR2 和 TLR4 表达量在刚加入菌液的 3h 开始上升,TLR2 在 12h 达到高峰,随着时间的延长而逐渐降低。而在 3h~24h 之间,isdA-组的 TLR2 和 TLR4 的表达水平明显低于 isdA+组(*P<0.05,**P<0.01)(图3)。 Hacat上皮细胞经过金葡菌IsdA+/-作用3~12 h后,与空白对照组相比,明显上调了抗炎性细胞因子Dectin-1和LL-37 mRNA的表达(*P<0.05,**P<0.01)。IsdA缺失后,3h~12h之间其表达水平均明显下调,并在3 h下降最明显;而作用24 h后,却无明显变化(P>0.05)(图4)。Hacat细胞可以自身表达TLR2、TLR4、Dectin-1、LL-37、IL-6、IL-8、TNF-α和MCP-1,且在3 h~24 h之前的表达并无明显差异;进行金葡菌处理后,mRNA的转录水平可以不同程度的升高。而缺失IsdA基因后表达水平均明显下调,个别因子在24 h无明显变化,但仍然显著高于对照组。
2.3Newman及NewmanΔIsdA对上皮细胞Toll样受体、Dectin-1和抗菌肽LL-37蛋白水平变化的影响 金黄色葡萄球菌可促进 Hacat 上皮细胞 TLR2、TLR4、Dectin-1 和 LL-37 蛋白表达上调,TLR2 比 TLR4 上调明显;而缺失 IsdA 基因后,这4个蛋白表达均下降。

图2 IsdA基因敲除对Hacat上皮细胞IL-8I、L-6、TNF-α和MCP-1 mRNA表达的影响
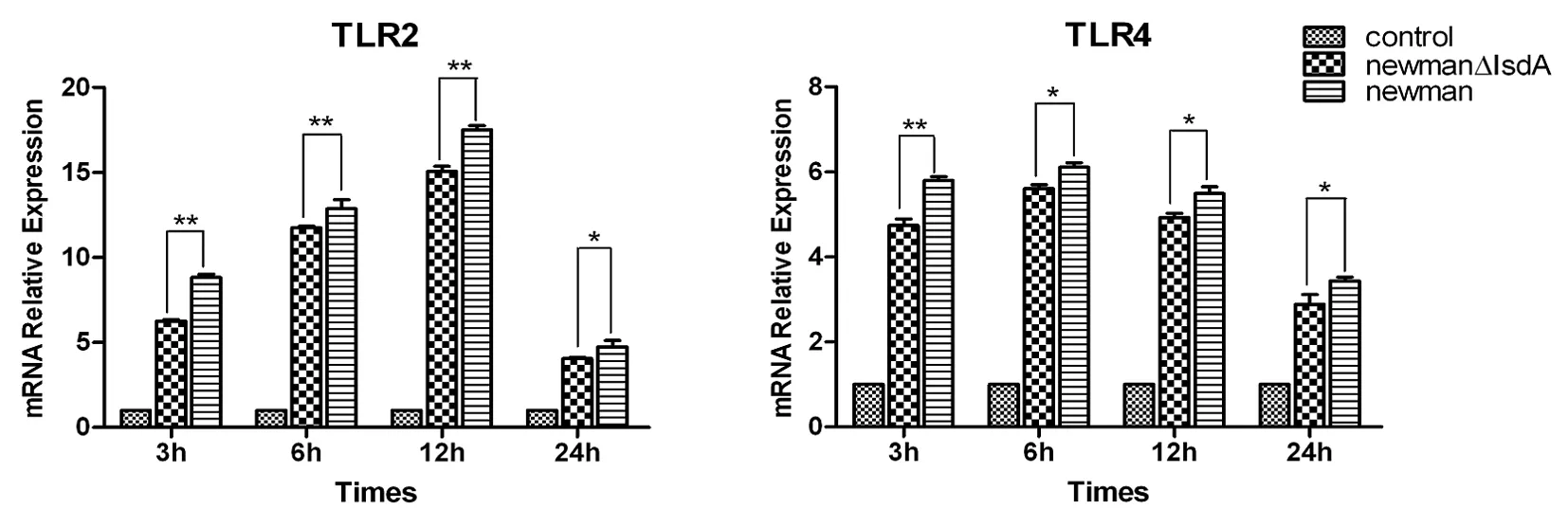
图3 IsdA基因缺失对Hacat上皮细胞表面受体TLR2和TLR4 mRNA表达的影响

图4 IsdA基因缺失对Hacat上皮细胞Dectin-1与LL37 mRNA表达的影响
3 讨 论
金葡菌因表达大量毒力因子而被称为最具致病力的细菌之一。金葡菌可引起包括皮肤感染在内的一系列感染,严重时甚至可以威胁人类的生命。角质化上皮细胞作为金葡菌感染的目标细胞,在金葡菌感染的先天免疫和炎症反应中扮演重要的角色,具有炎性介质合成和释放的能力,是宿主皮肤炎症的积极参与者。人们发现金葡菌表面蛋白IsdA对细菌细胞的表面疏水性的降低可以使它们具有杀菌作用,对人体皮肤上的金葡菌的生存非常必需[7]。因此,本研究选择了人的角质化上皮细胞Hacat做为细胞模型,在体外研究了金葡菌IsdA+/-对上皮细胞活性的影响。
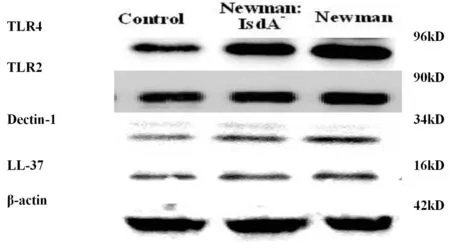
图5IsdA基因缺失对Hacat上皮细胞TLR2、TLR4、Dectin-1与LL-37蛋白表达的影响
Fig.5EffectsofIsdAdeletiononTLR2,TLR4,Dectin-1andLL-37proteinexpressioninHacaTkeratinocytes
哺乳动物先天免疫系统是通过模式识别受体(PRRs)进而检测病原微生物感染。PRRs包括Toll样受体(TLRs)、Nod样受体(NLRs)和RIG样受体(RLRs)三个家族,TLRs是现在研究比较成熟的受体家族,一些TLRs(TLR2和TLR4)在细胞表面上表达,识别病原体相关分子模式(PAMPs),在天然免疫和获得性免疫中均有重要作用。最近有研究证明金葡菌细胞壁上的结合蛋白具有作为宿主细胞表面TLR2原始配体的功能。然而该细胞表面结合蛋白有效识别TLR2的机制仍然不清楚;也有证据表明,TLR2在金葡菌中起着调节宿主反应的重要作用。事实上TLR2缺陷型小鼠对金葡菌感染高度敏感[8]。金葡菌可导致皮肤细胞的TLR2表达显着增加,从而提高了皮肤感染症状发展过程中宿主皮肤对细菌的敏感性。此外,据报道,细菌致脓毒血症儿童血清细胞因子TNF-α,IL-1β,IL-6和IL-8水平明显升高[9]。本实验结果与此相一致:葡萄球菌可以明显上调Hacat上皮细胞的TLR2表达水平,金葡菌增加炎性细胞因子IL-6、IL-8和TNF-α的表达,而TLR4的表达上调并不显著。本研究发现金葡菌作用于上皮细胞后,可显著的上调上皮细胞模式识别受体TLR2、TLR4、Dectin-1和抗菌肽的表达,并促进炎症细胞因子IL-6、IL-8、TNF-α和趋化因子MCP-1的产生。
IsdA是一个多功能的表面蛋白,野生型金葡菌诱导上皮细胞的细胞因子产生,从而导致体内发生炎症反应,而IsdA基因缺失的金葡菌诱导上皮细胞的细胞因子水平降低,因此我们推测IsdA是金葡菌的重要毒力因子之一。在本实验中毒力因子IsdA被敲出后的金葡菌与野生株相比,IsdA基因缺失后的金葡菌降低了Hacat上皮细胞细胞因子IL-8、TNF-α、IL-6和MCP-1的分泌水平和TLR2、TLR4、Ddectin-1和LL-37的蛋白表达水平,但高于空白对照组。证实了IsdA被敲出后的金葡菌的感染力下降,为日后金葡菌病的防治开辟了新的途径。
参考文献:
[1]Torres VJ, Pishchany G, Humayun M, et al. Staphylococcus aureus IsdB is a hemoglobin receptor required for heme iron utilization[J]. J Bacteriol, 2006, 188: 8421-8429. DOI: 10.1128/JB.01335-06
[2]Cheng AG, Kim HK, Burts ML, et al. Genetic requirements forStaphylococcusaureusabscess formation and persistence in host tissues[J]. FASEB J, 2009, 23: 3393-3404. DOI:10.1096/FJ.09-135467
[3]Friedman DB, Stauff DL, Pishchany G, et al.Staphylococcusaureusredirects central metabolism to increase iron availability[J]. PLoS Pathog, 2006, 2(8): e87. DOI:10.1371/Journal.ppat.0020087
[4]Grigg JC, Cooper JD, Cheung J, et al. The Staphylococcus aureus siderophore receptor HtsA undergoes localized conformational changes to enclose staphyloferrin A in an arginine-rich binding pocket[J]. J Biol Chem, 2010, 285: 11162-11171. DOI:10.1074/Jbc.M109.097865
[5]Bai X. Regulation of macrophages and myoblast function byexcretory/secretory products fromTrichinellaspiralisinvitro[D]. Changchun: Jilin University, 2011:37-47. (in Chinese)
白雪. 旋毛虫排泄分泌物调节巨噬细胞及成肌细胞功能的体外研究[D]. 长春: 吉林大学,2011:37-47.
[6]Kenneth J, Thomas D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method[J]. Methods, 2001, 25(4): 402-408. DOI: 10.1006/Meth.2001.1262
[7]Clarke SR, Mohamed R, Foster SJ. TheStaphylococcusaureussurface protein IsdA mediates resistance to innate defenses of human skin[J]. Cell Host Microbe, 2007, 1: 199-212. DOI:10.1016/J.chom.2007.04.005
[8]Kawai T, Akira S. The roles of TLRs, RLRs and NLRs in pathogen recognition[J]. Int Immunol, 2009, 21(4): 317-337. DOI: 10.1093/Intimm/dxp017
[9]Zhao AL. Correlation between serum procalcitonin and inflammatory factors in children with bacterial aepsis[J]. J Pediatr Pharm, 2013, 19(8): 5-7. (in Chinese)
赵安玲. 脓毒血症儿童血清降钙素原与炎症因子的相关性[J]. 儿科药学杂志,2013,19 (8):5-7. DOI:1672-108X( 2013) 08-0005-03.
