贝氏高原鳅脑组织学观察
2014-03-29王志坚
黄 静 王志坚
(西南大学生命科学学院, 淡水鱼类资源与生殖发育教育部重点实验室, 水产科学重庆市市级重点实验室, 重庆 400715)
贝氏高原鳅脑组织学观察
黄 静 王志坚
(西南大学生命科学学院, 淡水鱼类资源与生殖发育教育部重点实验室, 水产科学重庆市市级重点实验室, 重庆 400715)
鱼类体内各种器官分工合作维持着整个身体的正常生理活动, 神经系统与体液相互协作调控着鱼类的生长、发育、繁殖等一系列生命活动。对鱼类神经系统的研究是鱼类学研究中的一个重要方面, 有助于阐明鱼类的生态习性和对鱼类各种生命活动的理解。到目前为止, 对于淡水真骨鱼类脑的研究主要集中在鲤科鱼类, 对爬鳅科的研究尚未见报道。
贝氏高原鳅 Triplophysa bleekeri, 隶属于鲤形目Cypriniformes, 爬鳅科Balitoridae, 高原鳅属Triplophysa,是一种小型底栖鱼类, 生活于开阔河流及山溪石滩浅水处。多名学者分别对其繁殖生物学、血液学、消化系统、泌尿系统、鳃、肾脏、肝脏、垂体的组织学结构等进行了研究[1—9], 笔者已对脑的大体解剖结构进行了观察[10]。本文进一步对贝氏高原鳅脑进行组织学观察, 旨在完善对该鱼的研究, 丰富淡水真骨鱼类中枢神经系统组织学内容, 充实比较神经生物学的研究资料。
1 材料与方法
1.1 实验鱼
实验鱼采自巫溪大宁河, 于循环水养殖系统中暂养,不间断充气。选取健康、性成熟个体用于实验。
1.2 实验方法
HE染色 用于观察组织基本结构。在MS 222麻醉后, 取脑用Bouin’s液固定24h, 酒精梯度脱水, 二甲苯透明, Lecia EG1150型石蜡包埋机包埋, Reichert-Jung820旋转切片机连续冠状和矢状切片, 切片厚 6—10 μm, HE染色, NIKON 80i显微镜观察、照相, Image pro Plus 6.0软件测量。
Nissl染色 尼氏体为神经细胞所特有, 取材必须新鲜, 因机体死亡后尼氏体会很快溶解、分散, 不易着色。Bouin’s液或10%中性福尔马林液固定, 酒精梯度脱水, 二甲苯透明, 石蜡包埋, 切片厚 6—10 μm, 常规脱蜡、梯度酒精复水, 蒸馏水洗1—2min, 用1%的硫堇水溶液于60℃温箱内浸染 30min, 蒸馏水洗 1—2min, 无水酒精脱水8—10min, 二甲苯透明 8—10min, 中性树胶封片, NIKON 80i显微镜观察、照相, Image pro Plus 6.0软件测量, spss16.0软件对数据进行统计分析。神经细胞的胞浆中有大量的核糖核酸, 主要以尼氏体形式存在, 因而染成蓝色;细胞核中的染色质少, 故染色浅, 但核仁染色较深。神经胶质细胞的核中染色质多, 着色较深, 但胞浆不着色。
2 结果
贝氏高原鳅整个脑呈中间稍宽, 两端尖细的梭形,分为端脑、间脑、中脑、小脑和延脑5部分。
2.1 端脑
端脑由嗅叶和大脑组成。嗅叶分化为嗅球和嗅束。
嗅球表面为上皮组织。嗅球周缘富含神经纤维, 由外向内分为神经纤维外层、小球层、神经纤维内层(图版Ι-1)。嗅神经末梢、血管和僧帽状细胞构成小球层。僧帽状细胞胞体特别大, 呈球形、椭圆形或梭形, 轴突明显, 细胞长径(10.69±0.24) μm, 短径(7.07±0.27) μm(图版Ι-2)。神经纤维外层染色较深, 且不均匀。神经纤维内层染色浅而均匀, 其间有散在的圆形细胞。嗅球中央区域分布的神经细胞多而密集, 为中等大小细胞, 细胞直径(4.87±0.42) μm,细胞核占胞体大部分, 胞质少, 细胞多呈圆形、椭圆形,该层为小细胞层(图版 Ι-1), 在此层也可见少量多角形细胞和较大的梭形细胞。
嗅球中的纤维由周缘及内边各处的细胞延展, 构成嗅束。每一嗅束呈中空的圆筒状, 这一空心叫做嗅茎腔,此腔由嗅球后部起首(图版Ι-3)。
大脑由背面的大脑皮和腹面的纹状体组成(图版Ι-4)。背面的大脑皮是一薄膜, 薄而透明, 为上皮组织, 无神经组织。大脑皮与纹状体侧腹面相连, 之间的空隙即为公共脑室, 两基叶之间的空隙也是公共脑室的一部分。大脑前与嗅茎相连, 公共脑室向前通过嗅茎的中心孔到嗅球的小腔。每一纹状体表面皆有小沟, 将纹状体分成3个小叶:在大脑中部的小叶名中轴叶, 在大脑的侧部者名侧叶,在大脑的后侧部者为楔叶。在侧叶与楔叶之间的小沟颇深,较显著, 名席氏沟(Y状沟), 其余脑沟不明显, 但可观察到各脑沟将大脑分成的不同功能区, 即中央嗅区、背嗅区、侧嗅区、梨状区。神经细胞主要集中分布于各区的外周, 形成对应的神经核。
左右纹状体中下部富含神经纤维, 神经细胞分布稀疏, 细胞体积大, 细胞核明显。在纹状体内侧, 即公共脑室中间两侧, 从前至后细胞排列甚密集, 为连前核所在,其表面分布有室管膜细胞。连前核细胞分成两群, 背侧群区域较大, 细胞较小, 成索分布, 数量较多, 呈圆形或近椭圆形, 胞质少, 细胞直径(5.65±0.36) μm; 腹侧群区域小, 细胞较大, 数量少, 呈圆形或梨形、多角形, 胞质较丰富, 尼氏体明显, 细胞直径(7.22±0.25) μm。
界沟与席氏沟之间的背嗅区有一细胞排列较密的神经核, 为背嗅核(图版 Ι-5)。背嗅核细胞密度较连前核小,细胞较小[直径: (5.54±0.09) μm], 一些体积较大的细胞分布于外围, 细胞多呈圆形或近椭圆形。席氏沟与内嗅沟之间的侧嗅区分布有侧嗅核(图版 Ι-5), 侧嗅核细胞密度较背嗅核小, 细胞大小[直径: (7.33±0.44) μm]较背嗅核大,但比纹状体中部神经细胞[直径: (10.07±0.54) μm]小, 细胞以圆形为主。在大脑半球后端, 两个核团之间的界限变得模糊。在侧嗅核后部、端脑的尾部具有排列疏松的梨状核, 该核细胞小, 呈梨状或锥状, 也可作为侧嗅核的一部分。在其腹侧, 近内嗅沟处为细胞排列甚密的带核, 带核区小, 细胞圆形, 细胞直径(5.42±0.26) μm。
大脑后段腹侧出现前连合横行的纤维, 左右纹状体通过前连合相连。大脑的后部, 前连合的上方, 中间室的两侧有大小细胞明显分成两群的视前核(图版 Ι-6)。视前核大细胞为背侧群, 细胞直径(7.40±0.37) μm, 细胞形态呈梭形、多角形。视前核小细胞位于大细胞的腹侧和背内侧, 细胞小, 细胞直径(4.91±0.31) μm, 呈圆形或梭形, 前与连前核相接, 向后延伸。视前核的外侧, 端脑的基部有一小区, 细胞密度大, 为脚内核, 细胞小, 细胞直径(4.15±0.10) μm, 多呈圆形。
2.2 间脑
间脑分为上丘脑、丘脑和下丘脑三部分。上丘脑包括背囊、松果体、松果缰及缰连和, 背囊和松果体非神经组织。松果体为内分泌腺, 较小, 近圆形, 由背囊中的室管膜细胞分化而成, 其组织与背囊其余组织不同, 染色较深。松果缰在上丘脑中央空隙的侧壁呈半圆突, 左右各一,形成缰核(图版 Ι-7)。两缰核的纤维形成缰连和。松果缰细胞较小[直径: (5.26±0.25) μm], 呈颗粒状, 纤维把细胞分作数群。丘脑的本体在上丘脑的腹面, 侧壁增厚成视丘,位于视盖腹面, 细胞群分布于中央。间脑腹面的下丘脑在丘脑腹面, 前方是视交叉, 中央是呈长圆形、左右各一的下叶, 两下叶中间是脑下垂体, 垂体的基部是漏斗, 漏斗的后方是单个血管囊。血管囊背侧是左右成对的半月形沟,为视前隐窝, 其周围细胞呈颗粒状。间脑内腔为第3脑室,下叶内腔为下叶腔, 是第3脑室侧面的延伸部分, 并深入漏斗, 向后通过中脑水管, 达于第4脑室。第3脑室顶部有脉络膜丛。
2.3 中脑
中脑的视盖神经纤维和细胞层交替排列, 分为 6层(图版Ι-9、10), 由内向外依次为: ①围脑室层, 由围脑室神经细胞和视盖深层纤维所组成。可进一步分为室管膜与视盖深层纤维层及结层。室管膜与视盖深层纤维层, 室管膜细胞分布疏松, 呈圆形或椭圆形, 染色深, 纤维束粗大;结层, 分布神经幼芽细胞, 细胞排列密集, 胞质极少, 胞核呈圆形, 细胞直径(5.47±0.12) μm, 染色较深; ②中央纤维层, 由传出神经纤维组成, 纤维环绕视盖的深层, 起联络作用, 较薄, 结构致密, 其神经细胞胞体位于围脑室层。此层零星分布有水平细胞; ③中央细胞层, 含有多极神经细胞、梭状神经细胞和梨形神经细胞等多种类型的神经细胞, 很薄。该层的神经纤维联系视盖, 形成一种臂的构造, 包括了位于背部的前臂和腹侧部的后臂。前臂较小,在视盖的背中部, 含有视神经的前枝纤维, 纵枕和丘脑的纤维也入于其中; 后臂较大, 在视盖的腹侧部, 含有视神经后枝的纤维及中脑和丘脑的纤维; ④表面纤维层, 此层最厚, 为接受传入联系的神经细胞和神经纤维, 含有视神经的前枝纤维, 稀疏分布有水平细胞、梭状细胞和较大的锥体细胞; ⑤视神经层, 细胞分布稀疏, 多呈圆形,有来自中央和两侧视束的视神经纤维, 纤维排列稀松; ⑥边缘层, 较薄, 纤维排列致密。
视盖与被盖之间的腔为视叶腔(图版 Ι-8)。左右两视盖相接处的腹面有纵枕(图版 Ι-11), 为视盖的一部分, 呈梯形, 细胞呈颗粒状, 细胞直径(3.57±0.10) μm。纵枕的中部略分离, 随着视盖向两侧突出, 其亦分成左右两部, 并逐渐减小。纵枕背面有联系左右视盖的纤维束, 下方是后连合, 是中脑与间脑共有的连合, 其中大部分纤维属于中脑。后连合是中脑背部前端与间脑的分界, 中脑腹部与间脑的分界之处在间脑下叶基部。在被盖各一侧与视盖相接处有一突起, 名半圆枕, 其中分布的神经核为瓣侧核。半圆枕之间、中线两侧各有一突起, 为中隆起, 其中有扁豆核。中隆起在中脑前部较显著, 往后不明显。瓣侧核的侧面为峡核。峡核的腹面为中脑深核, 与侧纵束侧面彼此贴近。中脑腹部前端接近丘脑, 近中线处分布有 III脑神经(动眼神经)核。III脑神经由此核起首, 纤维向腹面延伸。在中脑与丘脑下叶之间伸出脑外, 形成III神经的根。III核尾部又分成IV核, 细胞较小, IV神经(滑车神经)根自中脑腹面发出。中脑腹部有网状核。中脑水管位于被盖正中, 前与视叶腔相通, 后接第4脑室。
2.4 小脑
横切面上小脑由外至内分为分子层、蒲氏细胞层和颗粒层(图版 Ι-12)。分子层细胞较小, 呈圆形, 细胞直径(5.09±0.18) μm, 分布稀疏, 染色浅, 纤维多; 蒲氏细胞层围绕颗粒层, 细胞比分子层及颗粒层中的细胞都大, 细胞直径(12.41±0.25) μm, 分布较稀疏, 形成一薄层, 还常分散于分子层中, 极少数分布于颗粒层中, 细胞数量不多; 颗粒层细胞多而密, 呈颗粒状, 细胞直径(2.96± 0.09) μm。颗粒层从两侧突破分子层向下扩展入侧叶。侧叶颗粒状细胞排列稍稀疏, 名颗粒隆起(图版 Ι-14)。小脑各层的厚度在不同位置有所差异, 而蒲氏细胞层总是最薄的。小脑瓣组织结构分层与小脑一致, 颗粒层向两侧扩展成倒“γ”形。小脑腹部因分子层内凹形成小脑腹脊, 覆于延脑之上(图版Ι-13)。
2.5 延脑
纵切面可观察到延脑实际上就是被盖向后的延续。延脑中间是面叶, 较发达, 为一独突, 前端略分离, 呈两半,向后融合为一体, 周缘有细胞密集分布, 中间成团分布,一些细胞散布其间, 细胞大小不一, 胞体呈锥形、梭形、圆形或椭圆形, 锥形细胞最大。两侧为迷叶, 细胞集中分布于周缘, 细胞大小、形态与面叶相似(图版 Ι-15)。第 4脑室有脉络膜分布, 其由富含血管的软膜和室管膜相贴突入脑室形成, 细胞呈立方形或短柱状, 核大, 呈圆形,核径(3.42±0.08) μm。
延脑的腹部由前至后分布有运动细胞, 组成了 V、VI、VII、VIII核及颖核(IX、X运动神经纤维的来源)。延脑前部正中线两侧各有一个巨大的神经细胞, 为莫氏细胞, 细胞直径(37.62±0.14) μm。后部几个比莫氏细胞稍小的大细胞为苗勒氏细胞, 细胞直径(30.31±0.25) μm。
3 讨论
贝氏高原鳅嗅叶分化为嗅球和嗅束, 属“鲤型”嗅叶[11],表面为上皮组织, 周缘富含神经纤维, 由外向内分为神经纤维外层、小球层、神经纤维内层, 中央为小细胞层,各层细胞无明确界限, 该结果与报道的齐口裂腹鱼(Schizothorax prenanti)、鲤鱼(Cyprinus carpio)的嗅球结构相似[12, 13]。
大脑神经核团的分布与大多数真骨鱼类相似, 主要划分成中央嗅区、背嗅区、侧嗅区及梨状区[12,14,15]。大脑连前核依细胞大小及疏密不同, 分为了背侧群和腹侧群, 这与秉志[13]描述鲤鱼的连前核细胞分为数群不同。贝氏高原鳅的连前核背侧群区域较大, 细胞较小, 成索分布, 数量较多, 呈圆形或近椭圆形, 胞质少, 腹侧群区域小, 细胞较大, 数量少, 呈圆形或梨形、多角形, 胞质较丰富, 尼氏体明显, 该特征与潘康成等[12]描述的齐口裂腹鱼连前核背侧群区域较小、细胞较小、细胞数量相对较少, 常呈椭圆形或梨形、三角形, 胞质少; 腹侧群区域大,细胞大、圆、数量多, 在分群、细胞大小及胞质上相符, 数量及形态特征描述相反。视前核明显分成两群, 大细胞群位于背侧, 小细胞群位于大细胞群的腹侧和背内侧, 这与潘康成等[12]报道的齐口裂腹鱼视前核大小细胞群的分布相同, 与秉志[13]报道的鲤鱼视前核大细胞被小细胞三面包围及王典群[16]、李仲辉等[17]、张耀光[18]分别研究的一些鲤科鱼类、南方鲇(Silurus meridionalis)的视前核细胞颗粒状呈带状致密排列不同。
中脑视盖在横切面上, 呈神经纤维和细胞层交替排列的层状结构, 由内向外分为围脑室层、中央纤维层、中央细胞层、表面纤维层、视神经层及边缘层6层。分层情况与陈蓉等[19]报道的莫桑比克罗非鱼(Tilapia mossambica)、张耀光[18]报道的南方鲇、秉志[13]报道的鲤鱼在分层上稍有不同, 如张耀光、秉志将视盖分为了8层, 由内至外分别是室管膜与视盖深层纤维层、结层、视盖连和层、视盖臂纤维层、视神经前枝纤维层、席氏连和层、外结层、视神经层, 但从各层描述上视盖的分层是相似的, 只是不同的学者分层的原则不尽相同。
小脑由外至内亦分为分子层、蒲氏细胞层和颗粒层。所报道的真骨鱼类小脑的分层均较一致[17,20,21]。蒲氏细胞是小脑的特征细胞, 细胞大, 分布稀疏, 细胞数量不多。在哺乳动物中, 小脑在眼的运动中起着重要作用, 鱼类中也不例外, 尤其是蒲氏细胞, 其直接与眼球运动相关的神经细胞相联系[22]。
延脑面叶呈一独突, 但并不是所有的真骨鱼类的面叶都是单一的。鲢鱼(Hypophthalmichthys molitrix)、鳙鱼(Aristichthys nobilis)、条(Hemiculter leucisculus)、草鱼(Ctenopharyngodon idellus)等上层鱼的延脑未分化出面叶和迷叶, 面叶和迷叶呈细胞团, 并且很小; 鲢鱼、鳙鱼、条的面叶呈两团, 对称排列在第 4脑室两侧[16]。Evans[23]认为真骨鱼类面叶不是1个, 是2个, 1个面叶是由2个面叶愈合的结果。本论文也观察到贝氏高原鳅的面叶在前端略分离, 向后才融合为一体, 因而作者支持Evans的观点。
鱼类不同种间脑的形态存在明显的不同, 因为其适应不同的生态习性, 相应地发展了脑的不同部位。因而从脑各结构的形态及发达程度我们就可以初步推断出各种鱼的生态习性。贝氏高原鳅为鲤形目鱼类, 其脑组织学结构与所报道的其他鲤形目鱼类是相似的, 但不同鱼类适应不同的生境, 具有不同的生活习性及进化地位, 因而又呈现出不同。在生活习性相同的鱼类中, 脑的形态结构越相似, 亲源关系越近[16]。贝氏高原鳅生活于开阔河流或山溪石滩浅水处, 在摄食行为中, 主要以触须、体表的味蕾感知食物, 面叶较发达, 与视觉功能相关的中脑无生活于上层水体的鱼类发达[10]。从目前关于真骨鱼类脑组织学结构的研究对比看出, 如细胞的形状、排列分层等是较一致的。但学者们对真骨鱼类脑各结构细胞的大小、数量、类型、生化特征等研究相对较少, 目前很难归纳出不同进化地位及生态习性的鱼类在组织学方面有哪些显著差异, 这些方面有待开展深入研究。
[1] He X F, He J S, Yan T M. Reproductive characteristic of Triplophysa bleekeri in mabian river [J]. Journal of Southwest China Normal University (Natural Science Edition), 1999, 24(1): 69—73 [何学福, 贺吉胜, 严太明. 马边河贝氏高原鳅繁殖特性研究. 西南师范大学学报(自然科学版), 1999, 24(1): 69—73]
[2] Wang Z J. The reproductive biology and hematology of Triplophysa bleekeri (Teleostei: Cypriniformes) from daning river [D]. Thesis for Doctor of Science. Southwest University, Chongqing. 2011 [王志坚. 大宁河贝氏高原鳅繁殖生物学及血液学研究. 博士学位论文, 西南大学. 2011]
[3] Xiong H L, Yao Y H, Wang Z J. Study on the structure of the digestive tract in Triplophysa bleekeri [J]. Journal of Southwest China Normal University (Natural Science Edition), 2012, 37(1): 113—120 [熊洪林, 姚艳红, 王志坚. 贝氏高原鳅消化道的结构. 西南师范大学学报(自然科学版), 2012, 37(1): 113—120]
[4] Xiong H L, Yao Y H, Wang Z J. Histomorphology of the digestive system during post-embryonic development of Triplophysa bleekeri [J]. Chinese Journal of Zoology, 2013, 48(2): 144—152 [熊洪林, 姚艳红, 王志坚. 贝氏高原鳅消化系统胚后发育的形态及组织结构. 动物学杂志, 2013, 48(2): 144—152]
[5] Wen L L, Yao Y H, Wang Z J. The urinary system morphology in Triplophysa bleekeri [J]. Chinese Journal of Zoology, 2007, 42(6): 108—114 [温龙岚, 姚艳红, 王志坚. 贝氏高原鳅泌尿系统显微和超微结构. 动物学杂志, 2007, 42(6): 108—114]
[6] Ren S, Yan Z B, Ma X H, et al. Observation of gill structure of Triplophysa bleekeri [J]. Guizhou Agricultural Sciences, 2011, 39(6): 128—130 [任爽, 晏正碧, 马秀慧, 等. 贝氏高原鳅鳃结构的观察. 贵州农业科学, 2011, 39(6): 128—130]
[7] Wen L L, Yao Y H, Wang Z J. A study of the development of the kidney of Triplophysa bleekeri [J]. Journal of Southwest China Normal University (Natural Science Edition), 2009, 34(3): 179—183 [温龙岚, 姚艳红, 王志坚. 贝氏高原鳅肾脏发育研究. 西南师范大学学报(自然科学版), 2009, 34(3): 179—183]
[8] Xiong H L, Yao Y H, Wang Z J. Structure of the liver of Triplophysa bleekeri [J]. Journal of Southwest University (Natural Science Edition), 2010, 32(8): 87—91 [熊洪林, 姚艳红, 王志坚. 贝氏高原鳅肝脏的结构. 西南大学学报, 2010, 32(8): 87—91]
[9] Liu X H, He X, Geng X C, et al. Triplophysa bleekeri pituitary microstructure [J]. Sichuan Journal of Zoology, 2009, 28(6): 810—812, 815, cover 2 [刘小红, 何翔, 耿相昌, 等.贝氏高原鳅脑垂体显微结构观察. 四川动物, 2009, 28(6): 810—812, 815, 封2]
[10] Huang J, Peng Z G, Wang Z J. Comparison of the gross anatomy of the brains between Triplophysa bleekeri and Triplophysa rosa [J]. Journal of Southwest China Normal University (Natural Science Edition), 2013, 38(3): 94—100 [黄静, 彭作刚, 王志坚. 贝氏高原鳅与玫瑰高原鳅脑大体解剖结构的比较. 西南师范大学学报(自然科学版), 2013, 38(3): 94—100]
[11] Kottelat M. Freshwater Fishes of Northern Vietnam [M]. A preliminary check-list of the fishes known or expected to occur in Northern Vietnam with comments on systematics and nomenclature. The World Bank, Washington, 2001
[12] Pan K C, Fang J. Study on the telecephlon morphology and histology of Schizothorax prenat (Tchang) [J]. Journal of Sichuan Agricultural University, 2002, 20(2): 145—147 [潘康成, 方静. 齐口裂腹鱼端脑形态和组织学研究. 四川农业大学学报, 2002, 20(2): 145—147]
[13] Bing Z. Tissue of Cyprinus carpio [M]. Beijing: Science Press. 1983 [秉志. 鲤鱼组织. 北京: 科学出版社. 1983]
[14] Pushchina E V, Varaksin A A, Romanov N S. Neuronal organization of the telencephalon in the Chum Salmon Onchorhynchus keta [J]. Russian Journal of Marine Biology, 2003, 29(6): 362—367
[15] Huang X Z, Yang X P, Wang Q. Histological study of the telencephalon of mandarin fish Sinipeca [J]. Journal of Huazhong Agricultural University, 1996, 15(2): 169—171 [黄祥柱, 杨秀平, 王琪. 鳜鱼端脑组织学研究. 华中农业大学学报, 1996, 15(2): 169—171]
[16] Wang D Q. The observation on the morphology and organization of the brain of eight freshwater cyprinids [J]. Journal of Fisheries of China, 1986, 10(1): 95—106 [王典群. 八种鲤科鱼类脑的形态构造观察. 水产学报, 1986, 10(1): 95—106]
[17] Li Z H, Lu J Q, Wang Y, et al. Study on brain morphology of cyprindae in Henan province [J]. Sichuan Journal of Zoology, 1997, 16(3): 109—112 [李仲辉, 路纪琪, 王玉, 等. 河南鲤科鱼类脑的形态学研究. 四川动物, 1997, 16(3): 109—112]
[18] Zhang Y G. Observation on the morphology and organization of the brain and cranial nerves of Silurus Meridioanlis [J]. Journal of Southwest China Normal University (Natural Science Edition), 1992, 17(1): 102—106 [张耀光. 南方鲇脑和脑神经的观察. 西南师范大学学报(自然科学版), 1992, 17(1): 102—106]
[19] Chen R, Rao Y Z, Yang S J, et al. The histological study of the optic tectum in Tilapia mossambica [J]. Journal of Zhanjiang Normal College, 2006, 27(6): 58—61 [陈蓉, 饶颖竹,杨素娇, 等. 莫桑比克罗非鱼中脑视顶盖组织学结构观察.湛江师范学院学报, 2006, 27(6): 58—61]
[20] Fang Z Q, Chen L Y, Chen G Z. Observation on the morphology and histology of the brain in Tanichthys albonubes [J]. Chinese Journal of Zoology, 2006, 41(2): 24—28 [方展强, 陈丽玉, 陈国柱. 唐鱼脑的组织形态学观察. 动物学杂志, 2006, 41(2): 24—28]
[21] Wan A, Hua T M, Kong X J, et al. Morphology observation on the loach brain [J]. Journal of Biology, 2009, 26(6): 15—18 [万安, 华田苗, 孔学军, 等. 泥鳅脑的形态构造观察. 生物学杂志, 2009, 26(6): 15—18]
[22] Northmore D P M, Williams B, Vanegas H. The teleostean torus longitudinalis: responses related to eye movements, visuotopic mapping, and functional relations with the optic tectum [J]. Journal of Comparative Physiology, 1983, 150(1): 39—50
[23] Evans H M. The brain of Gadus, with special reference to the medulla oblongata and its variations according to the feeding habits of different Gadida-1 [J]. Proceedings of the Royal Society of London Series B, 1935, 117: 367—399
OBSERVATION ON THE HISTOLOGY OF THE BRAIN OF TRIPLOPHYSA BLEEKERI
HUANG Jing and WANG Zhi-Jian
(Key Laboratory of Freshwater Fish Reproduction and Development (Ministry of Education); Key Laboratory of Aquatic Science of Chongqing; Southwest University School of Life Science; Chongqing 400715, China)
贝氏高原鳅; 脑; 组织学
Triplophysa bleekeri; Brain; Histology
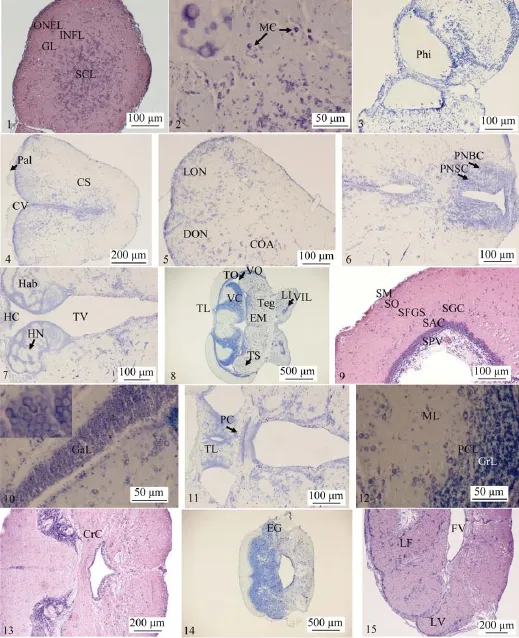
图版Ⅰ 贝氏高原鳅脑组织学结构PlateⅠ Histological structure of T. bleekeri brain
Q174
A
1000-3207(2014)03-0576-06
10.7541/2014.81
2013-04-16;
2014-01-12
西南大学博士基金(SWU111056); 重庆市科委重点实验室专项经费资助
黄静(1986—), 女, 重庆北碚人; 助理实验师, 硕士; 主要从事资源动物学研究。E-mail: a86728@163.com
王志坚(1969—), 男, 四川南充人; 教授, 博士; 主要从事鱼类资源与渔业生物学研究。E-mail: wangzj1969@126.com
