水生植物荇菜和菹草分解对物种混合的响应研究
2014-03-29巩崇贤
巩崇贤 王 东
(华中师范大学生命科学学院, 武汉430079)
水生植物荇菜和菹草分解对物种混合的响应研究
巩崇贤 王 东
(华中师范大学生命科学学院, 武汉430079)
为探讨水生植物混合的分解效应, 研究了浮叶植物荇菜(Nymphoides peltatum)、沉水植物菹草(Potamogeton crispus)及两物种混合的分解速率和养分动态。结果显示: (1)两单物种的分解速率与初始N含量呈显著正相关关系(P<0.05, r=0.862), 荇菜和菹草分解90d后的干重剩余率分别为24.74%和44.91%。物种混合干重剩余率在分解初期阶段的实测值比期望值高6.63% (P<0.05), 表明物种混合对分解速率具有拮抗效应, 但在随后的分解时间里无显著的混合效应, 分解90d后干重剩余率为30.39%; (2)在分解初期的N、P释放阶段, 物种混合的N、P剩余率实测值比其期望值分别高14.36%和12.88% (P<0.05), 表明物种混合对初期N、P元素释放具有拮抗效应, 在随后的分解过程中对N元素无显著的混合效应, 但分解后期P剩余率实测值比期望值低4.26% (P<0.05), 表现为协同效应; (3)物种混合N、P动态在分解初期呈一个快速释放的过程,但在随后的分解阶段N元素释放或积累, P元素持续释放, 最终N、P均表现为净释放, 与两单物种分解的N、P动态的规律基本一致。另外, 总酚在物种混合分解初期释放迅速, 随后释放缓慢。研究结果表明, 荇菜和菹草混合分解存在非加和效应, 即单物种的分解速率和营养动态变化不能用来预测两物种混合的分解速率和营养动态变化。物种混合在分解的不同阶段其分解效应不同, 这说明混合效应具出一定的时间依赖性。此外, 混合效应与浮叶植物和沉水植物其初始质量特征有较密切的关系。
水生植物; 混合; 分解速率; N、P动态; 非加和效应
水生植物是构成湖泊生物多样性的主体之一,植物残体的腐烂分解影响湖泊的元素循环和水体的营养平衡[1—3]。在淡水湖泊中, 水生植物以单优种群或由多种水生植物组成群落的情况普遍存在, 不同物种的残体经常在湖泊沿岸带混合堆积, 其有机物质在分解后归还水体[1,2]。前人对混合分解的研究表明, 物种混合对分解的影响可表现为加和效应(Additive effect), 即混合对分解速率和养分的释放速率没有显著影响; 或表现为非加和效应(Nonadditive effect), 包括协同效应(Synergistic effect)即提高分解速率和养分的释放速率和拮抗效应(Antagonistic effect)即降低分解速率和养分的释放速率[4,5]。已有的相关研究关于陆生植物的较多, 对水生植物腐烂分解的研究相对薄弱[6]。目前对水生植物多涉及单一物种的分解研究[2,3,7,8—10], 有关物种混合分解的研究报道很少。
荇菜(Nymphoides peltatum)和菹草(Potamogeton crispus)是长江中下游湖泊常见的水生植物, 植物组织的 N和P含量不同[11]。荇菜为根生浮叶植物, 菹草为沉水植物, 在扰动环境下它们可产生大量的茎叶残体并混合堆积于湖边沿岸带。为揭示荇菜和菹草混合的分解效应, 本文运用分解袋法研究了荇菜、菹草及其混合的分解速率及N、P释放动态, 以期为探讨水生植物的分解规律提供资料, 也为认识水生植物的整个生态过程和湖泊生态系统的管理提供参考。
1 材料与方法
1.1 实验材料
荇菜、菹草的新鲜茎叶于2012年5月16日取自华中师范大学南湖校区水生植物实验基地(N30°30′, E 114°21′), 材料用池水洗净带回实验室, 自然风干后剪成5 cm左右的小段, 在60℃下烘干至恒重, 样品放入干燥器并于10d后用于实验。
1.2 实验方法
将烘干处理后的荇菜、菹草和等量混合的荇菜和菹草三种样品各10 g分别装入尼龙网袋(大小20 cm× 15 cm, 网孔1 mm×1 mm)。采用经过筛处理(以去除杂物和根系)、洗干净的细沙[TC (0.67±0.20) mg/g、TN (0.07±0.05) mg/g、TP (0.12±0.01) mg/g, 根据底质干重获得]作为分解实验的基质, 沙厚约 1.5 cm,待稳定10d后作为植物腐烂分解实验反应器。将分解袋置于塑料盆(54 cm × 27 cm × 6 cm, 长×宽×高)中, 保持分解袋处于湿润状态, 一个处理每个样品6袋(按6次取样计)并置于1个塑料盆中, 3组重复。实验共计9个塑料盆、54袋样品。取部分剩余材料用于实验材料初始质量特征参数的测定。在实验中,塑料盆在生物园室内实验区有遮雨处随机放置, 保持盆中无杂物和分解袋湿润。
1.3 样品采集与分析
于2012年5月26日将分解袋放置在塑料盆中,在分解袋放置后的第10、第20、第30、第40、第60和第90天分别取样, 实验周期为90d。每种材料每次在每组处理中分别取1袋, 共3袋。带回实验室后用纯净水冲去沙子等杂物。60℃烘干至恒重, 称重后,磨碎过0.25 mm筛, 用于样品中TC、TN、TP、总酚含量及干重测定。采用K2Cr2O7氧化-FeSO4滴定法测定TC, 材料经 H2SO4-H2O2消化, 分别采用靛酚蓝比色法和钼锑抗比色法测定TN和TP[12]; 采用范氏分析方法(Analysis of Van Soest)测定木质素、纤维素、半纤维素含量[13]; 采用福林酚法测定总酚含量[14]。
1.4 统计分析
用指数分解模型描述分解材料干重变化: Wt=W0×e–kt(t为分解时间; k为分解常数; W0为初始干重; Wt为经 t天分解后的剩余干重), 用非线性回归分析计算分解速率[15]。为了直观地反映分解材料的损失情况, 采用剩余率表示分解过程中干重和养分含量的变化。干重剩余率和养分剩余率分别为分解后材料干重和养分剩余量占初始量的百分率。
根据Salamanca等[16]采用的方法计算混合材料的期望干重剩余率和养分剩余率: 期望干重剩余率=[M1/(M1+M2)]×R1+[M2/(M1+M2)]×R2; 期望养分剩余率=[N1/(N1+N2)]×R1+[N2/(N1+N2)]×R2。式中, 以1、 2表示混合物中的两种组分, M和N分别表示各组分在初始混合物中的干重和养分含量, R表示各组分单一物种的干重剩余率和养分剩余率。对混合材料混合分解时是否存在混合效应进行判断: 若实测值与期望值之间差异显著(P<0.05), 则表示混合材料各组分之间存在非加和效应, 实测值大于期望值,则混合效应是负的, 即拮抗效应, 实测值小于期望值, 则混合效应是正的, 即协同效应。若实测值与期望值之间差异不显著(P>0.05), 则表示混合材料各组分之间存在加和效应, 即无明显的相互作用。
使用统计软件 SPSS 17.0 进行数据分析, 采用单因素方差分析(One-way analysis of variance)检验差异性水平, 若差异显著, 则采用 Duncan法(Duncan’s multiple comparison test)对具显著性差异的处理间进行多重比较检验。对3种材料的干重剩余率、C、N、P、总酚剩余率、C/N、C/P的比较先进行方差齐性检验, 如方差不齐则进行对数转换(lg-transformed)。实测值与期望值的比较, 采用配对样本 t检验其差异性水平。对于两单种之间质量特征、分解速率的比较采用独立样本 t检验其差异性水平。对荇菜和菹草的分解速率与其初始N含量的相关性进行Pearson相关分析。
2 结果
2.1 荇菜和菹草的初始质量特征
荇菜和菹草在 C、N、P、C/P、木质素、纤维素、总酚含量上存在显著差异(P<0.05)(表 1)。与菹草相比, 荇菜具有较高的 C、N、C/P、木质素、总酚含量和较低的 P和纤维素含量; 荇菜的 C/N、半纤维素含量与菹草的差异不显著。
2.2 荇菜、菹草及其混合分解的干重动态变化
荇菜和菹草干重动态变化均表现为先快后慢(图 1), 其分解速率存在显著差异(P<0.01), 分解速率分别为 0.032/d和 0.017/d, 实验结束时干重剩余率分别为24.74%和 44.91%。荇菜 20d左右可分解50%的干重。菹草 40d左右可分解 50%的干重。Pearson相关分析表明, 两单物种的分解速率与初始N含量呈显著正相关关系(P<0.05, r=0.862)。荇菜和菹草混合的分解速率为0.023/d, 其分解速率介于两物种单独分解速率之间(表2)。1个月内可分解50%的干重。实验结束时荇菜和菹草混合的干重剩余率为30.39% (表3)。物种混合对干重剩余率在不同分解阶段有不同的分解效应, 在分解的第10天, 实测值比期望值高 6.63%(P<0.05), 表现出拮抗作用, 随后的时间里实测值与期望值无明显差异(P>0.05),表现为加和效应(表3)。

表1 荇菜和菹草的初始质量特征Tab. 1 Initial quality characteristics of N. peltatum and P. crispus materials
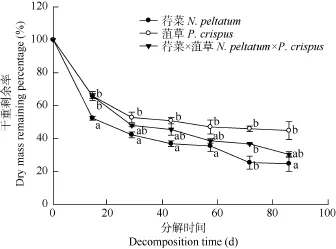
图1 荇菜、菹草及其混合在分解过程中的干重动态变化Fig.1 Mass loss dynamics of N. peltatum and P. crispus and their mixture in decomposition
2.3 荇菜、菹草及其混合分解的N、P动态变化
荇菜、菹草的N剩余率在最初10d下降明显, 分别为 62.30%和 64.68%。在随后的时间里, 荇菜 N剩余率呈继续下降趋势, 到90d N剩余率略有增加,但仍低于初始值; 菹草N剩余率呈不规律的变化。在分解90d后荇菜和菹草的N剩余率分别为36.33%和53.09%。荇菜、菹草的P剩余率在最初10d下降明显, 分别为 21.13%和 36.59%。在随后的时间里,荇菜、菹草P剩余率在总体上呈平缓下降趋势(荇菜在第90天和菹草在第60天P剩余率略有增加)。在分解90d后荇菜和菹草的P剩余率分别为11.84%和26.93%。总体上, 荇菜N、P的释放速率大于菹草N、P的释放速率, 最终均表现为净释放(图 2)。

表2 荇菜、菹草及其混合的分解速率比较Tab. 2 Comparison of the decomposition rates among and between N. peltatum and P. crispus and their mixture
荇菜和菹草混合在整个分解过程中, N、P剩余率变化均先表现出明显的快速下降, 随后呈继续平缓下降趋势(除N剩余率在第60天略有上升外), 在分解90d后N、P剩余率分别为43.60%和15. 88% (图3)。在分解的第10天, 物种混合N、P剩余率实测值比其期望值分别高 14.36%和 12.88%, 且差异显著(P<0.05), 表现出拮抗效应, 物种混合在分解初期抑制了N、P元素的释放。在随后的时间里, 除在分解的第90天P剩余率实测值比期望值低4.26%,且差异极显著(P<0.01)外, N、P剩余率实测值与期望值均无显著差异, 说明物种混合对N元素的释放无显著影响, 对 P的拮抗效应逐渐减弱并在最终表现出协同效应。此外, 物种混合N、P动态在分解初期呈一个快速释放的过程, 但在随后的分解阶段N元素表现出释放或积累, P元素持续释放, 最终 N、P均表现为净释放。总酚在物种混合分解初期迅速释放, 随后缓慢释放, 与物种混合的N、P释放规律表现出一致的趋势。
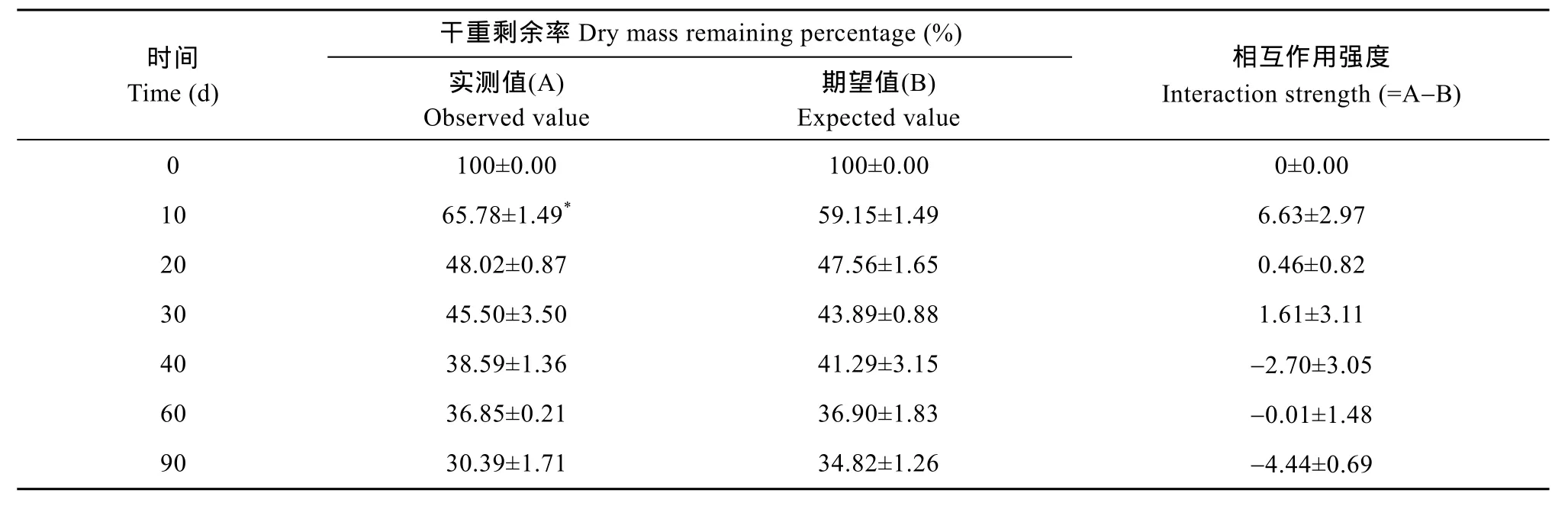
表3 荇菜和菹草混合分解的干重剩余率(%)(实测值和期望值)Tab. 3 Observed and expected values of the dry mass remaining percentage (%) of the mixed N. peltatum and P. crispus
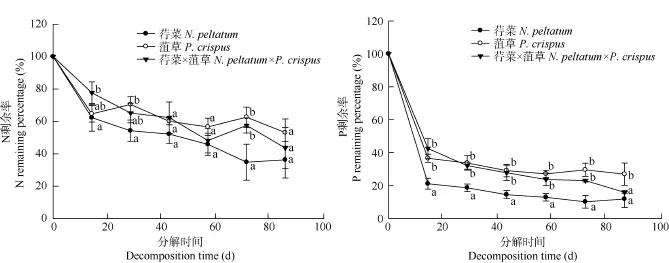
图2 荇菜、菹草及其混合在分解过程中的N、P剩余率(%)变化Fig. 2 Changes of nitrogen and phosphorus remaining percentage (%) of N. peltatum, P. crispus, and its mixture during the experiment
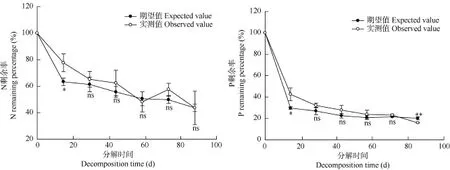
图3 荇菜和菹草混合分解的N、P的实测剩余率(%)和期望剩余率(%)Fig. 3 The values of observed and expected nitrogen and phosphorus remaining percentage (%) of the mixed N. peltatum and P. crispus materials
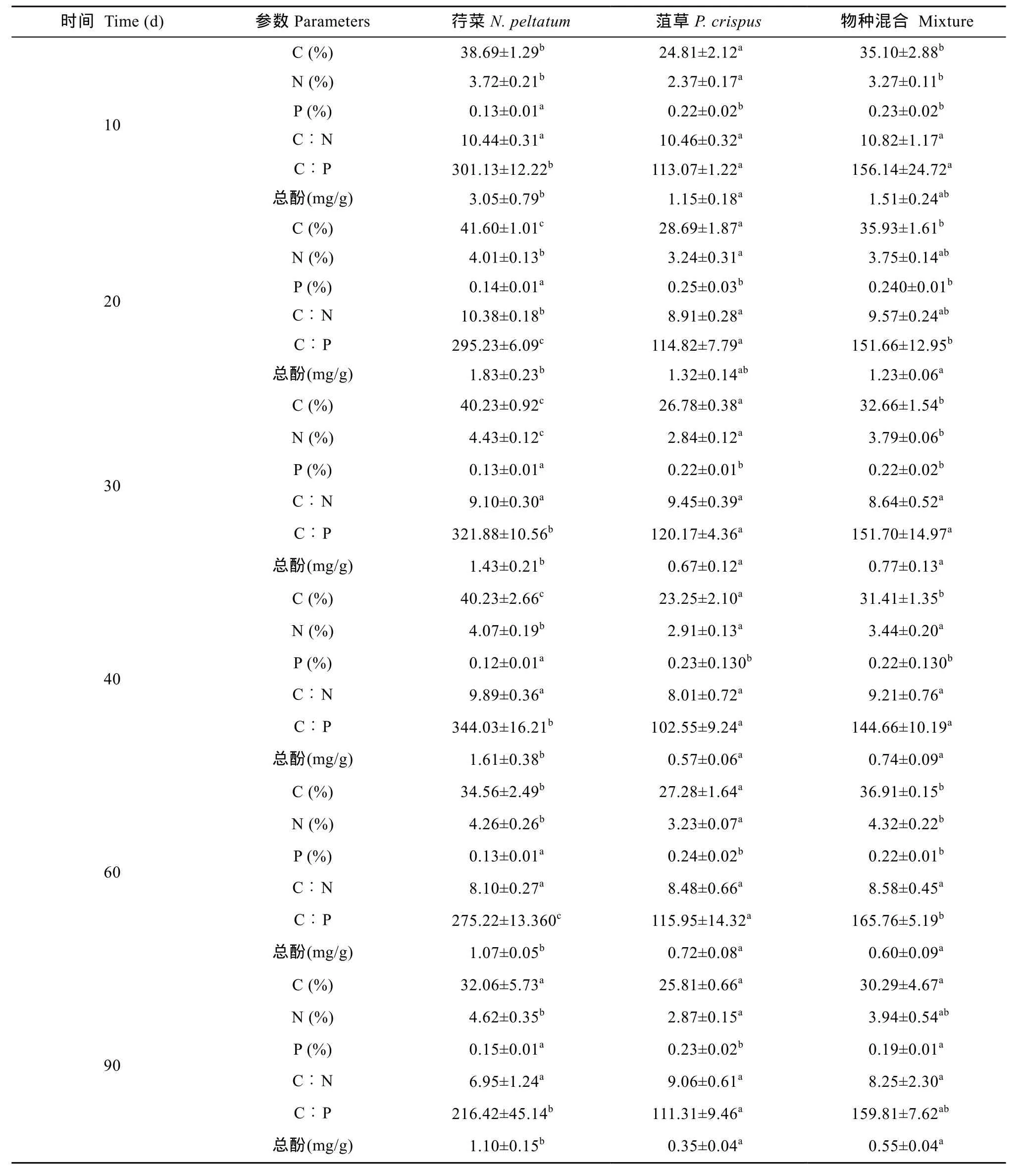
表4 荇菜和菹草及其混合在分解过程中的质量特征变化Tab. 4 Changes of quality characteristics of N. peltatum, P. crispus and their mixture during the decomposition
3 讨论
3.1 质量特征对荇菜和菹草分解的影响
植物组织的质量特征是影响分解快慢的重要生物因素[3,17]。其中, N含量对初始阶段的分解有较大的影响, 初始N含量较高其分解较快。荇菜的初始N含量显著高于菹草, 荇菜的分解速率显著高于菹草, 这表明荇菜和菹草的分解快慢也受其初始N含量所控制。另外, 具有较高P含量和较低C/P比的材料其分解也相对较快[18], 菹草较荇菜具有较高的P含量和较低的 C/P, 但菹草的分解速率小于荇菜,这可能与初始N含量较P含量和C/P对分解速率的影响更显著有关[17]。此外, 有研究表明分解速率与材料初始纤维素、半纤维素、木质素含量、酚类化合物的含量呈负相关[1,7,19]。在本研究中, 荇菜较菹草有较高的总酚和木质素含量, 但荇菜仍表现出较高的分解速率, 荇菜的干重剩余率和N、P剩余率也均小于菹草, 这可能与荇菜在不同的分解阶段各种物质的化学组成变化对微生物活动的影响有关。一些酚类物质可能会改变酶的结构使酶失活, 或与营养蛋白结合后阻碍微生物对N元素的利用, 从而降低分解速率等[20,21]。荇菜的初始纤维素含量显著低于菹草, 荇菜的分解速率显著高于菹草, 这表明荇菜和菹草的分解快慢也与其初始纤维素含量有关。有研究发现淋溶作用可导致水生植物组织中的可溶性成分在初期分解阶段迅速释放, 一般在 24h干重损失率可达初始重量的25%[1]或50%[22—24]。本研究也发现在分解前10d荇菜和菹草腐烂分解均有较高的干重损失率和N、P含量的快速下降过程, 这在一定程度上表明淋溶使荇菜和菹草丧失了大部分养分。有研究表明, 植物在腐烂分解过程中其N剩余率的变化与微生物固定N的趋势有密切关系[25], 微生物对N的固定将导致分解材料N含量的升高。分解材料中N含量越低微生物固定N的趋势越强[26],反之亦然。在本实验中, 荇菜N剩余率呈持续下降趋势, 仅在分期后期(第90天时)略有增加。这说明由于荇菜初始N含量较高, 其自身N源能够满足微生物活动的需求, 以至于在分解后的很长一段时间内微生物没有发生对外源N元素的固定, 仅在分解后期外源N素被微生物所固定, 使植物残余物中的氮素浓度有所上升。另外据Brady和Weil[27]的研究发现, 当植物残余物的C/N比超过30, N元素的固定就已经开始, 相反, 如果小于 30, N元素将被矿化。荇菜初始C/N比小于30, 在分解第10、第20、第30、第40和第60天时, 残余物中的C/N比也小于30, 这说明荇菜N的释放动态与Brady和Weil[27]的研究结论是一致的。与荇菜N剩余率变化趋势比较, 菹草N剩余率表现出不规律的变化, 如在第20、第 60天略有增加, 而在其他时间呈下降趋势。这与菹草初始N含量较荇菜低, 但初始C/N较荇菜高, 在分解的不同阶段微生物对外源N元素产生固定有关。
与分解过程N动态相比较, 荇菜和菹草的P剩余量在分解过程中总体上均呈平缓下降趋势, 且一直表现为释放。尽管荇菜的初始P含量较菹草低, 但相对较低的初始P含量并不导致微生物对P固定。有研究发现, 当初始 C/P<100时, P元素发生矿化,相反, 如果大于100, P元素将被固定[28,29]。荇菜和菹草初始 C/P比分别为 110.32±1.90和 82.03±0.59,但本研究中荇菜的初始C/P比似乎并不能解释荇菜分解过程中的P元素的释放动态。对荇菜和菹草来说, 微生物在分解过程中的营养需求没有受到 P供给的限制, 或者说没有发生微生物对P元素的固定。
3.2 物种混合对分解速率和养分释放的影响
不同物种混合对分解的响应可表现为非加和效应或加和效应[4,5]。据Gartner和Cardon[4]统计, 在物种混合分解中出现非加和效应的比例约占 70%,其中协同效应占 50%, 拮抗效应占 20%, 而出现加和效应的比例占30%。同时, 76%的物种混合在分解过程中表现出非加和的养分动态。在本研究中, 荇菜和菹草混合仅在分解初期对分解速率和N释放具有显著的拮抗效应, 在随后的分解阶段对分解速率和N释放均不产生混合效应; 但分解过程中却在一定程度上都促进了 P释放或积累。有研究表明, 当不同质量的物种混合时, 由于营养物质(如 N)通过被动扩散和微生物的主动运输等从高质量材料中向低质量材料发生转移, 改善了质量较差材料的营养状况, 从而促进混合物的分解速率[30]; 但也有一些研究发现酚类物质可以控制可溶性有机氮和无机氮的释放比例[31], 酚类物质的释放会抑制微生物丰度和活性从而降低混合物种的分解速率[32]。在本研究中, 荇菜和菹草的初始N和总酚含量存在显著差异,但N含量的差异对分解初期的混合效应作用似乎并不明显, 而总酚释放动态则与混合后的分解速率和N释放有相同的趋势。具有一面叶气生的荇菜比完全沉水生长的菹草具有更高的总酚含量, 在混合分解初期总酚也出现一个快速释放过程, 之后出现缓慢释放趋势。此外, Bonanomi等[5]在研究地中海物种混合分解时发现, 相对于对 N动态的影响而言,混合效应对分解速率的影响更具有分解时间上的依赖性, 非加和效应的产生更多发生在分解初期阶段,并认为物种质量特征对物种混合是否产生混合效应起到了决定性的作用。也有研究发现物种混合并不影响分解速率, 但显著加快N元素的释放[33]。在本结果中, 在分解初期荇菜和菹草混合抑制了干重的损失, 但在随后的分解阶段分解没有发现显著的混合效应, 混合效应表现出明显的时间依赖性。N释放动态在混合分解初期之后表现出加和性, 而 P元素的释放动态在分解初期、后期均表现出显著的非加和性, 这说明P元素对混合的响应比N元素更为敏感。
水生植物腐烂分解对湖泊生态系统将产生影响,由于受环境扰动如风浪、人类活动的(捕捞鱼虾和打捞水草等)影响, 植物的茎叶残体经常混合堆积在湖泊沿岸带。尽管水生植物在人工湿地或极端环境的水体中也存在单种群落的情况, 但淡水湖泊中由多种水生植物组成群落的情况普遍存在, 水生植物残体的混合分解是必然的。本研究初步表明, 荇菜和菹草混合的分解效应在干重损失率和营养元素的释放动态方面不同于两单种的分解规律。在分解的初始阶段物种混合对分解速率具有拮抗效应, 但在随后的分解时间里无显著的混合效应; 同时物种混合对N、P元素释放具有拮抗效应, 在随后的分解过程中对N元素无显著的混合效应, 但在分解后期对 P释放具有协同效应。由于在野外自然状态下, 环境因子的影响作用复杂, 另外也不涉及环境容量对分解过程的影响, 以及实验过程中材料的减少对容量影响产生的作用。因此, 很有必要加强野外条件下的相关研究。
[1] Webster J R, Benfield E F. Vascular plant breakdown in freshwater ecosystems [J]. Annual Review of Ecology and Systematics, 1986, 17: 567—594
[2] Battle J M, Mihuc T B. Decomposition dynamics of aquatic macrophytes in the lower Atchafalaya, a large floodplain river [J]. Hydrobiologia, 2000, 418(1): 123—136
[3] Shilla D, Asaeda T, Fujino T, et al. Decomposition of dominant submerged macrophytes: implications for nutrient release in Myall Lake, NSW, Australia [J]. Wetlands Ecology and Management, 2006, 14(5): 427—433
[4] Gartner T B, Cardon Z G. Decomposition dynamics in mixed-species leaf litter [J]. Oikos, 2004, 104(2): 230—246
[5] Bonanomi G, Incerti G, Antignani V, et al. Decomposition and nutrient dynamics in mixed litter of Mediterranean species [J]. Plant Soil, 2010, 331(1-2): 481—496
[6] Gustafsson C, Boström C. Biodiversity influences ecosystem functioning in aquatic angiosperm communities [J]. Oikos, 2011, 120(7): 1037—1046
[7] Godshalk G L, Wetzel R G. Decomposition of aquatic Angiosperms. . Ⅲ Zostera marina L. and a conceptual model of decomposition [J]. Aquatic Botany, 1978, 5(4): 329—354
[8] Li W C, Chen K N, Wu Q L, et al. Experimental studies on decomposition process of aquatic plant material from East Taihu Lake [J]. Journal of Lakes Sciences, 2001, 13(3): 331—336 [李文朝, 陈开宁, 吴庆龙, 等. 东太湖水生植物生物质腐烂分解实验. 湖泊科学, 2001, 13(3): 331—336]
[9] Xie Y H, Yu D, Ren B. Effects of nitrogen and phosphorus availability on the decomposition of aquatic plants [J]. Aquatic Botany, 2004, 80(1): 29—37
[10] Li E H, Liu G H, Li W, et al. Decomposition rate and nitrogen and phosphorus dynamics of three kinds of hydrophyte in Lake Honghu [J]. China Environmental Science, 2006, 26(6): 667—671 [厉恩华, 刘贵华, 李伟, 等.洪湖三种水生植物的分解速率及NP动态. 中国环境科学, 2006, 26(6): 667—671]
[11] Wu A P, Wu S K, Ni L Y. Study of Macrophytes nitrogen and phosphorus contents of the shallow lakes in the middle reaches of Changjiang River [J] Acta Hydrobiologica Sinica, 2005, 29(4): 406—412 [吴爱平, 吴世凯, 倪乐意. 长江中游浅水湖泊水生植物氮磷含量与水柱营养的关系. 水生生物学报, 2005, 29(4): 406—412]
[12] Dong M, Wang Y F, Kong F Z, et al. Survey, Observation and Analysis of Terrestrial Biocommunities [M]. Beijing: China Standards Press. 1996, 152—155 [董鸣, 王义凤, 孔繁志, 等. 陆生生物群落调查观测与分析. 北京: 中国标准出版社. 1996, 152—155]
[13] Van Soest P J. Use of detergents in analysis of fibrous feeds: a rapid method for the determination of fiber and lignin [J]. Journal of the Association of Official Analytical Chemists, 1963, 46(5): 829—835
[14] Box J D. Investigation of the Folin-Ciocalteau phenol reagent for the determination of polyphenolic substances in natural waters [J]. Water Research, 1983, 17(5): 511—525
[15] Olson J S. Energy storage and the balance of producers and decomposers in ecological systems [J]. Ecology, 1963, 44(2): 322-331
[16] Salamanca E F, Kaneko N, Katagiri S. Effects of leaf litter mixtures on the decomposition of Quercus serrata and Pinus densiflora using field and laboratory microcosm methods [J]. Ecological Engineering, 1998, 10(1): 53—73
[17] Aerts R, Caluwe H D. Nutritional and plant-mediated controls on leaf litter decomposition of Carex species [J]. Ecology, 1997, 78(1): 244—260
[18] Berg B, Staaf H. Decomposition rate and chemical changes of Scots pine needle litter. II. Influence of chemical composition [J]. Ecological Bulletins, 1980, 32: 373—390
[19] Freeman C, Ostle N J, Fenner N, et al. A regulatory role for phenol oxidase during decomposition in peatlands [J]. Soil Biology and Biochemistry, 2004, 36(10): 1663—1667
[20] Harrison A F. The inhibitory effect of oak leaf litter tannins on the growth of fungi, in relation to litter decomposition [J]. Soil Biology and Biochemistry, 1971, 3(3): 167—172
[21] Hättenschwiler S, Vitousek P M. The role of polyphenols in terrestrial ecosystem nutrient cycling [J]. Trends in Ecology and Evolution, 2000, 15(6): 238—243
[22] Puriveth P. Decomposition of emergent macrophytes in a Wisconsin marsh [J]. Hydrobiologia, 1980, 72(3): 732—739
[23] Polunin N V C. The decomposition of emerged macrophytes in freshwater [J]. Advances in Ecological Research, 1984, 14(1): 115—166
[24] Vähätalo A V, Søndergaard M. Carbon transfer from detrital leaves of eelgrass (Zostera marina) to bacteria [J]. Aquatic Botany, 2002, 73(3): 265—273
[25] Gessner M O. Mass loss, fungal colonization and nutrient dynamics of (Phragmites australis) leaves during senescence and early aerial decay [J]. Aquatic Botany, 2001, 69(2): 325—339
[26] Baker T T, Lockaby B G, Conner W H, et al. Leaf little decomposition and nutrient dynamics in four southern forested floodplain communities [J]. Soil Science Society of America Journal, 2001, 65(4): 1334—1347
[27] Brady N C, Weil R R. The Nature and Properties of Soils [M]. Vol. 11. Prentice-Hall, Englewood Cliffs, NJ. 1996, 740
[28] Enwezor W O. The mineralization of nitrogen and phosphorus in organic materials of varying C: N and C: P ratios [J]. Plant and Soil, 1976, 44(1): 237—240
[29] Rejmánková E, Houdková K. Wetland plant decomposition under different nutrient conditions: what is more important, litter quality or site quality [J]? Biogeochemistry, 2006, 80(3): 245—262
[30] Wardle D A, Bonner K I, Nicholson K S. Biodiversity and plant litter: experimental evidence which does not support the view that enhanced species richness improves ecosystem function [J]. Oikos, 1997, 79(2): 247—258
[31] Northup R R, Yu Z S, Dahigren R A, et al. Polyphenol control of nitrogen release from pine litter [J]. Nature, 1995, 377(6564): 227—229
[32] Schimel J P, Cates R G, Ruess R. The role of balsam poplar secondary chemicals in controlling soil nutrient dynamics through succession in the Alaskan taiga [J]. Biogeochemistry, 1998, 42(1): 221—34
[33] Blair J M, Parmelee R W, Beare M H. Decay rates, nitrogen fluxes and decomposer communities of single-and mixedspecies foliar litter [J]. Ecology, 1990, 71(5): 1976—1985
EFFECT OF MIXED FLOATING AND SUBMERGED MACROPHYTES ON DECOMPOSITION RATE AND NUTRIENT DYNAMICS
GONG Chong-Xian and WANG Dong
(School of Life Sciences, Central China Normal University, Wuhan 430079, China)
Decomposition of aquatic macrophytes can considerably influence nutrient cycling and energy flow in aquatic ecosystems, and may therefore alter aquatic ecosystem structure and functioning. Most studies of decomposition processes have focused on single species of aquatic macrophytes; however, most aquatic ecosystems consist of a mixture of aquatic plant species and fragments they produce which intermingle during decomposition. To explore the effect of species mixtures of aquatic macrophytes with different life forms, decomposition rate and nutrient dynamics were quantified in mixed-species litterbags (containing Nymphoides peltatum, a floating-leaved plant, and Potamogeton crispus, a submerged plant) and in litterbags containing fragments of a single species in a laboratory experiment. There were 10 g of materials used for each species in the litter bags, and for the mixture experiment, also 10 g of the dried macrophyte fragments were used at a mixtures rate of 5:5, Nymphoides:Potamogeton, w/w basis). The decomposition rates, nitrogen and phosphorus content of the remaining materials were determined after 10, 20, 30, 40, 60, and 90 days. Data from single-species litterbags were used to generate expected decomposition rates, nitrogen and phosphorus dynamics for mixed-species litterbags experiments. The result showed that the decomposition rates of N. peltatum (0.032/d) was pronounced higher than that of P. crispus (0.017/d), 24.74% and 44.91% dry mass remaining after 90 days, respectively. The decomposition rates of both N. peltatum and P. crispus were significantly and positively correlated with initial N content (P < 0.05, r = 0.862). The decomposition rate of the mixture was 0.023/d which was intermediate between N. peltatum and P. crispus. The observed remaining mass of the mixture at the early stages of decomposition in ten days was 6.63% (P < 0.05) higher than the expected, indicating the occurrence of negative, non-addtivie effects of mixed species early on. In contrast, there was no significant mixing effect after ten days in subsequent samplings. After 90 days, the remaining dry mass of the mixture was 30.39%. The N and P contents of both N. peltatum and P. crispus released rapidly at the early stages and then slowed down. The remaining percentage of N and P of N. peltatum were lower than that of P. crispus. During the early stages of decomposition of mixed material in ten days, the observed N and P remaining were 14.36% and 12.88% (P < 0.05) higher than the expected, indicating the occurrence of antagonistic effects on N and P release in the mixture. However, there were no significant antagonistic mixing effects in subsequent times for N. After 90 days, the observed P remaining was 4.26% (P < 0.05) lower than expected, indicating a synergistic effect on P release occurred. The remaining percentage of N and P were 43.60% and 15.88%, respectively. Nutrients and polyphenol concentrations in the mixture decreased rapidly at the early stages and then decreased slowly through the end of the study, in a manner similar to that of the single species. Our results indicated that there were negative, non-additive effects on decomposition rate, N and P releases when two species were mixed together at the early stages, while there was a synergistic effect on P release in the final stage of the decomposition. This suggests that neither decomposition nor nutrient release patterns can be assessed on basis of single species dynamics. In addition, there was a significant time-independent non-additive effect of species interactions. We further suggest that different aquatic macrophytes of contrasting life forms such as floating-leaved plants and submerged plants may differ in initial chemical quality and may exhibit major determinants for decomposition of mixed aquatic macrophytes.
Aquatic macrophyte; Mixed-species; Decomposition rate; Nitrogen and phosphorus dynamics; Non-additive effect
Q948.8
A
1000-3207(2014)06-1098-09
10.7541/2014.161
2014-01-14;
2014-04-23
国家自然科学基金项目(31270378); 中央高校基本科研业务费专项(CCNU12A02006)资助
巩崇贤(1987—), 男, 山东枣庄人; 硕士研究生; 研究方向为湿地生态学。E-mail: 1933759109@qq.com
王东, E-mail: dongwang.cn@gmail.com
