小陇山林区锐齿栎原始林的径级结构与物种多样性
2014-03-26刘文桢郭小龙张宋智马建伟王军辉
刘文桢,郭小龙,张宋智,马建伟,王军辉
(1 甘肃省小陇山林业实验局 林业科学研究所,甘肃 天水 741022;2 中国林业科学研究院 林业研究所,北京 100091)
森林群落在物种组成和结构上表现出的多样性,是认识群落组织水平以至功能状态的基础,也是生物多样性研究中至关重要的方面[1]。物种多样性反映群落或生境中物种的丰富度、变化程度或均匀度,定量表征群落和生态系统的特征,包括直接或间接地体现群落和生态系统的结构类型、组织水平、发展阶段、稳定程度、生境差异等[2],也可反映不同自然地理条件与群落的相互关系,对深入了解森林植物群落的组成、结构、功能具有重要意义[3-6]。
小陇山林区位于甘肃省东南部,地处秦岭西端,在我国华中、华北、喜马拉雅、蒙新四大自然植被区系的交汇处,拥有我国最大的国有林场群,是西北地区重要的天然林区。由于长期受到人为干扰的影响,目前该林区绝大多数天然林为次生林。从20世纪60年代开始,小陇山林区就开展了“次生林综合培育技术研究”;近10多年来,在森林可持续经营思想的指引下,小陇山林区开展了可持续经营实践,先后引进了“近自然森林经营理念”、“结构化森林经营技术”等现代森林经营方法,探索了小陇山林区次生林可持续经营理论与技术。在“近自然森林经营理念”、“结构化森林经营技术”等现代森林经营理念中,均将“师法自然”作为一条重要的原则,即在森林经营中尽量以“潜在的自然植被”或未经人为干扰的天然林为模板作为森林经营的方向。因此,了解和认识研究区原始林群落的组成、多样性、结构和动态等特征,对运用现代森林经营技术开展森林可持续经营具有重要的意义。以往对小陇山林区森林群落的研究,主要集中在次生林的群落特征方面,特别是对构成该地区的主要森林群落锐齿栎次生林群落的生物量、群落数量特征、群落结构、种间关系、生物多样性等方面的特征研究较多[3-5,7-9],尚未见到有关小陇山林区锐齿栎原始林群落的报道。因此,本研究在小陇山百花林场仙坪营林区设立一个锐齿栎天然林样地,该样地地处深山,地理位置偏僻,自小陇山林业实验局1964年建局以来,没有经营历史记录,林中锐齿栎最大年龄达120年以上,可以认为是锐齿栎原始林。本研究通过对该样地植被群落组成、多样性和结构等的分析,探讨小陇山锐齿栎原始林的植被特征,以期为小陇山林区开展次生林可持续经营提供参考和依据。
1 研究区及样地概况
1.1 研究区概况
研究样地设在小陇山林业实验局百花林场仙坪营林区内,地理坐标106°27′12.9″E、34°14′55.1″N,地势由西北向东南倾斜,山体相对高度不大,坡度一般在26°~45°。最高海拔2 489.2 m,最低海拔1 442.0 m。年平均气温10.9 ℃,极端最高气温38.3 ℃,年均降水量800 mm,年平均蒸发量1 420 mm, 湿润度0.36~0.45,相对湿度69%,无霜期184.8 d,土壤以山地棕壤和山地褐土为主。该区域是未遭到人为干扰的少数区域之一。
小陇山林区海拔2 200 m以下主要分布的是以锐齿栎(Quercusalienavar.acuteserrata)和辽东栎(Quercusliaotungensis)为主的天然林;在栎林带内分布有华山松(Pinusarmandi)、白桦(Betulaplatyphylla)、山杨(Populusdavidiana) 、漆树 (Rhusverniciflus)、冬瓜杨(Populuspurdomii)、千金榆(Carpinuscordata)、甘肃山楂(CrataeguskansuensisBL)、刺楸(Kalopanaxseptemlobus)等乔木树种,灌木有美丽胡枝子(Lespedezathunbergii)、光叶绣线菊(SpiraeajaponicaL. f.var.fortunei)、中华绣线菊(Spiraeachinensis)、胡颓子(Elaeagnuspungens)、华北绣线菊(Spiraeafritschiana)、连翘(Forsytiasuspense)、卫矛(Euonymusalatus)、山豆花(Lespedezatomentosa)等。
1.2 样地概况
样地位于百花林场仙坪营林区91林班的14小班内,海拔1 857 m,坡向东北,平均坡度32°,土壤为山地棕壤,土层厚度52 cm;林分郁闭度为0.9,样地内林木平均胸径24.3 cm,平均树高16.6 m,蓄积量277.59 m3/hm2,树种组成为5栎1鹅耳栎1槭1椴2其他;样地内存在大量的枯立(倒)木,其中枯立木168株,枯倒木37株,最大树木胸径61.3 cm,年龄为120年以上。样地内灌木树种主要有箭竹(Sinarundinarisnitida)、鞘柄菝葜(Smilaxstans)、青荚叶(Helwingiajaponica(Thunb.) Dietr.)、长柄八仙花(HydrangealongipesFranch.)、桦叶荚蒾(ViburnumbetulifoliumBatal.)、绣线梅(NeilliathrysifloraD.Don)、宝兴茶藨子(RibesmoupinenseFranch.)等;草本有秦岭金腰子(ChrysospleniumbiondianumEngl.)、裂叶荨麻(Urticalotabifolia)、酢浆草(OxaliscorniculataLinn.)、羊胡子(Carexbreviculmisvar.fibrillosa)等。
2 研究方法
2.1 样地设置
在研究区内设置面积为1 hm2(100 m×100 m)的样地,样地采用NTS320全站仪测设,在确定了样地的基轴坐标后,用全站仪将整个样地划分为25块小样地(20 m×20 m),角桩用8 cm×10 cm×70 cm的水泥桩标记。采用GPS测定样地地理坐标、海拔,用罗盘仪测定样地坡向、坡度。
2.2 群落组成的调查与统计
调查记录样地内树高大于1.3 m的乔木树种的胸径、冠幅、株数、树高、枝下高,并用全站仪(GTS-602AF)进行定位,树高测量采用超声波测高仪;统计乔木树种的种类,计算各树种的多度、相对多度、相对频度、胸高断面积和重要值,胸径大于3 cm的灌木归入乔木中进行统计,乔木层重要值用式(1)计算;分别在25块小样地的中心设置5 m×5 m的样方,调查灌木和草本的种类,并分析草本和灌木的种类组成。
乔木层重要值=(相对多度+相对频度+相对优势度)/3。
(1)
2.3 林木直径结构分析
对样地内胸径大于3 cm的林木进行统计,根据群落组成,分树种分析建群种和主要伴生树种的径级结构,并运用统计软件Forstat 2.0进行拟合。林分直径分布拟合采用负指数函数:y=ke-ax,式中,e为自然对数函数的底数,x为径级,y为各径级拟合株数[10-18]。
2.4 多样性分析
根据对不同林层植物种类、数量的调查结果,采用Shannon-Wiener指数、优势度和Pielou均匀度指数分析不同层次的多样性特征,计算方法[7-9]如下:
(2)
式中:pi为第i个物种的个体数与所有物种个体总数的比值,S为林分中的物种数。

(3)
Pielou均匀度指数:E=H′/lnS。
(4)
3 结果与分析
3.1 锐齿栎原始林的群落组成
样地内乔木树种共有21科30属53种1 528株,包括裸子植物1科1属1种16株,被子植物20科29属52种1 512株;其中,北温带分布的科有槭树科(Aceraceae)、忍冬科(Caprifoliaceae)、杨柳科(Salicaceae)、山茱萸科(Cornaceae)和松科(Pinaceae),北温带和南温带间断分布的科有桦木科(Betulaceae)和虎耳草科(Saxifragaceae),泛热带分布的科有壳斗科(Fagaceae)、木犀科(Oleaceae)、榆科(Ulmaceae)、樟科(Lauraceae)、天南星科(Araceae)、卫矛科(Celastraceae)和漆树科(Anacardiaceae),热带亚洲和热带美洲间断分布的科有椴树科(Tiliaceae)和省沽油科(Staphyleaceae),热带亚洲(印度-马来西亚)分布的科有清风藤科(Sabiaceae),世界分布的科有蔷薇科(Rosaceae)和豆科(Papilionoideae),东亚分布的科有领春木科(Eupteleaceae)和旌节花科(Stachyuraceae)。北温带分布的属有梣属(Fraxinus)、樱属(Cerasus)、榆属(Ulmus)、忍冬属(Lonicera)、山梅花属(Philadelphus)、山楂属(Crataegus)、省沽油属(Staphylea)、松属(Pinus)、唐棣属(Amelanchier)、铁木属(Ostrya)、盐肤木属(Rhus)、杨属(Populus)和桦木属(Betula),北温带和南温带间断分布的属有槭属(Acer)、梾木属(Swida)和柳属(Salix),地中海、东亚、新西南和墨西哥-智利间断分布的属有椴树属(Tilia)、鹅耳枥属(Carpinus)、花楸属(Sorbus)、荚蒾属(Viburnum)和栎属(Quercus),东亚(热带、亚热带)及热带南美间断分布的属有泡花树属(Meliosma),东亚分布的属有旌节花属(Stachyurus)和领春木属(Euptelea),东亚和北美洲际间断分布的属有香槐属(Cladrastis)、山胡椒属(Lindera)和漆属(Toxicodendron),旧世界温带分布的属有栒子属(Cotoneaster),世界分布的属有卫矛属(Euonymus),中国特有分布的属有双盾属(Dipelta)。由表1可见,样地内胸径≥3 cm的个体总胸高断面积为34.788 8 m2,其中胸高断面积大于1 m2的树种由大到小依次为锐齿栎、鹅耳枥、地锦槭、辽东栎、鄂椴、象腊树、桦椴、小叶鹅耳枥,这些树种胸高断面积之和占样地总胸高断面积的76.61%。由表1可知,重要值最大的是锐齿栎(0.223 8),其他树种在林分中虽有分布,但重要值都较小。锐齿栎种群胸高断面积占林分总胸高断面积的46.26%,为该群落的建群树种;此外,该群落中除具有当地的常见树种外,还出现了许多小陇山林区的稀有种,如膀胱果、领春木等,这2个种在群落中的相对频度、胸高断面积都较大,在人为干扰较大的地区已很少见到,说明该群落所受人为干扰较小,进一步表明该林分具有原始状态的特征。
小样方调查结果表明,群落中共有灌木33个属46个种,平均高度1.2 m,平均盖度27.9%,主要种有箭竹(Sinarundinarisnitida)、鞘柄菝葜(Smilaxstans)、青荚叶(Helwingiajaponica(Thunb.) Dietr.)、长柄八仙花(HydrangealongipesFranch.)、桦叶荚蒾(ViburnumbetulifoliumBatal.)、绣线梅(NeilliathrysifloraD.Don)、宝兴茶藨子(RibesmoupinenseFranch.)、细梗淡红荚蒾(Viburnumerubescensvar.gracilipes)、绢毛绣线菊(SpiraeasericeaTurcz.)、尖叶栒子(CotoneasteracutifoliusTurcz.)、绿叶胡枝子(LespedezabuergeriMiq.)、阔叶荚蒾(ViburnumlobophyllumGraebn.)、糖茶藨子(Ribeshimalense)、尖叶绣线菊(Spiraeajaponicavar.acuminata)、米面翁(Buckleyahenryi)、南川绣线菊(Spiraearosthornii)、盘叶忍冬(LoniceratragophyllaHemsl.)等,其中箭竹为中国特有种。草本共46个属58个种,平均高度0.31 m,平均盖度8.1%,主要种有秦岭金腰子(ChrysospleniumbiondianumEngl.)、裂叶荨麻(Urticalotabifolia)、酢浆草(OxaliscorniculataLinn.)、羊胡子(Carexbreviculmis var.fibrillosa)、活血丹(Glechomalongituba)、白花碎米荠(Cardamineleucantha(Tausch) O.E.Schulz)、钝萼铁线莲(ClematispeteraeHand-Mzt.)等。

表1 小陇山林区锐齿栎原始林样地乔木树种的物种组成
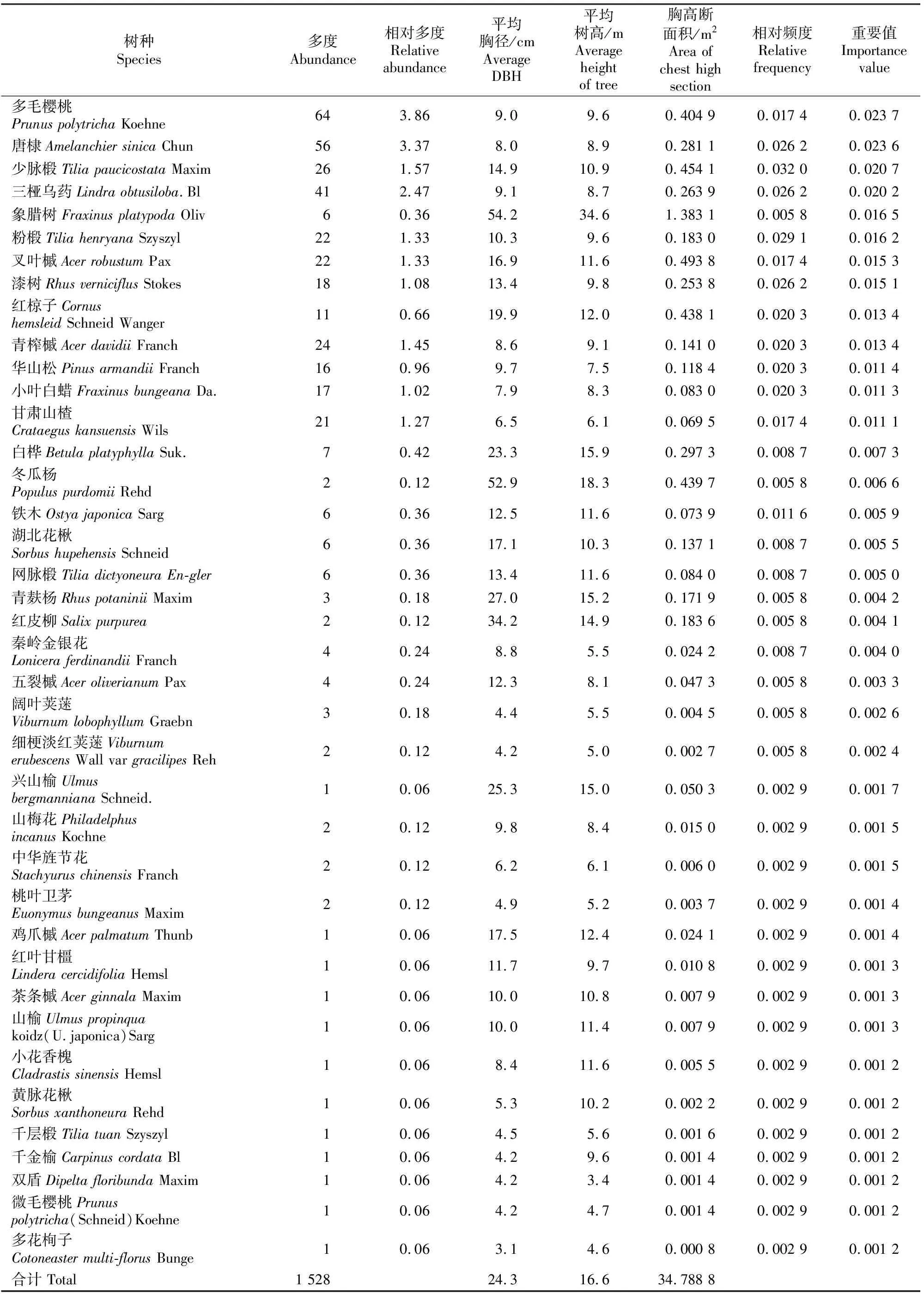
续表1 Continued table 1
3.2 锐齿栎原始林主要树种及林分的直径结构分析
林分直径分布是最重要、最基本的林分结构。由图1可以看出,小陇山锐齿栎原始林林分径级结构呈倒J型分布,林分中胸径大于30 cm的大径级木占总株数的9.3%,这部分林木主要集中在30~36 cm径级。林分直径分布拟合方程为:y=739.22e-0.094 1x(R2=0.969 2),拟合模型的q值(相邻径级株数之比)为1.353,介于1.3~1.7,呈现出典型的天然异龄林的径级分布特征。该群落中,中小径级林木占多数,胸径≤8 cm径级和胸径为12 cm的林木比例分别为52.23%和13.61%,占林分中总株数的65.84%;随着径级的增大,林木株数开始急剧减少,当径级达到36 cm时,林木株数减少幅度渐趋平缓。另外,径级在16 cm后呈现出小幅波浪式变动的趋势,出现这种现象可能是由于轻微的自然干扰或是群落内各树种种间、种内竞争的结果,但其并未对林分直径分布的整体趋势造成影响,林分直径分布仍呈倒J型。
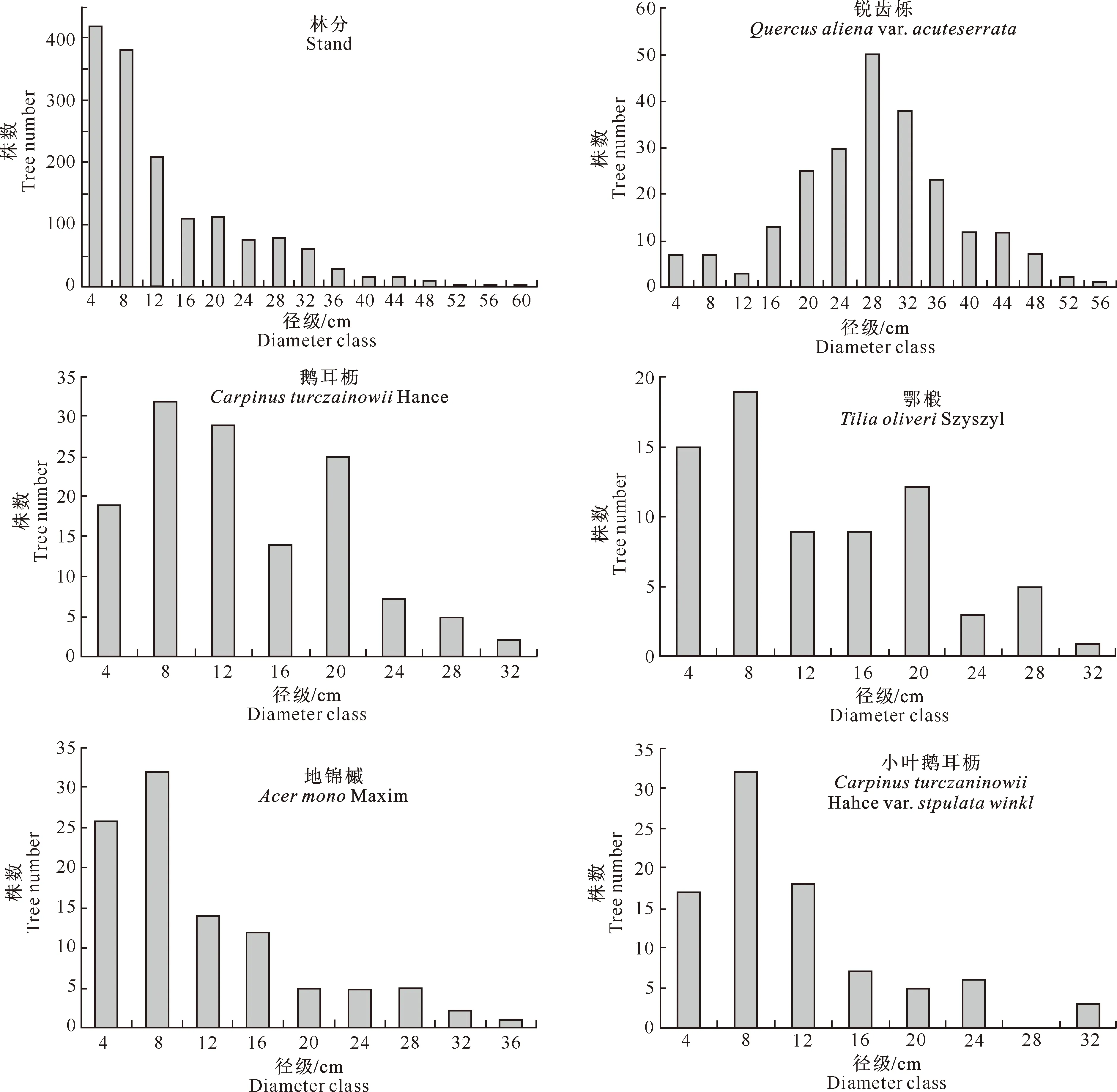
图1 小陇山锐齿栎原始林主要乔木树种和林分的径级分布
对群落中5个主要树种的径级结构进行分析,结果见图1。图1显示,群落中主要树种锐齿栎的平均胸径为29.8 cm,其径级结构为以平均胸径为峰值的单峰山状曲线,这与人工林的径级结构相似,为典型的正态分布,说明锐齿栎为该群落中最稳定的建群种。主要伴生树种鹅耳枥、小叶鹅耳枥、鄂椴3个树种的径级结构分布呈现多峰山状曲线,并且有“个体缺失”现象;而地锦槭则呈现倒J型分布,株数随着径级的增加而减少,属于进展型种群。
3.3 锐齿栎原始林群落的物种多样性
根据样地每木调查数据和小样方调查数据计算锐齿栎原始林不同林层的物种多样性,结果见表2。

表2 小陇山林区锐齿栎原始林群落乔、灌、草层的物种多样性
物种多样性是生物多样性在物种水平上的表现形式,是反映群落组织水平,继而通过结构与功能的关系间接反映群落功能特征的重要指标,维持物种多样性已成为森林可持续经营的一个主要内容和重要目标。由表2可以看出,样地内乔木层共有乔木树种53种1 528株,Shannon-Wiener指数为3.130,说明锐齿栎原始林乔木层的树种组成丰富,多样性较高。
优势度指数值越大,表明生态优势度越大,即优势树种的集中性越大,物种多样性越小;Pielou均匀度指数反映的是各物种个体数分配的均匀程度,其值越大说明物种分配的均匀程度越高。由表2可知,研究样地乔木层优势度指数与Pielou均匀度指数分别为0.063和0.955,说明锐齿栎原始林乔木层优势树种的集中性较小,物种分配较为均匀,是群落长期进展演替的结果。群落中灌木层和草本层的物种数分别为46种和58种,从数量上来看,物种组成也较为丰富,但灌木层和草本层的Shannon-Wiener指数分别为1.395和1.899,相对于乔木层而言,物种多样性较低;优势度和Pielou均匀度指数也表明,灌木层优势树种的集中性较大,树种分配相对集中,而草本层的物种优势度和均匀性介于乔木层与灌木层之间。
形成锐齿栎原始林群落乔灌草不同林层物种多样性和物种分配特征的原因,可能是由于群落经过长期进展演替趋向于稳定,乔木层占据了上层空间,林下得不到充分的光照,从而导致一些喜光的灌木植被和草本植被发展困难,这些植被只能在林下有限的空隙内生长,因此,灌木层和草本层的物种多样性较小、优势度较大而物种分配的均匀度较差,这还需进一步研究。
4 结论与讨论
小陇山林区是我国西北重要的天然林区,以锐齿栎为主的栎类天然林是其主要的林分类型,但由于历史上长期不合理的经营利用,现存栎类天然林多为次生林,因此如何合理地经营次生林是小陇山林区实现森林可持续经营的关键。对次生林而言,如果任其自由发展,有可能顺向演替为原来的状态,这是植被自然趋同的结果,也是次生林经营的基础,但这一过程相当漫长[19]。如果以长期未受干扰的锐齿栎天然林的群落特征为模板,采用恰当的森林经营措施对次生林进行人为干扰,加速其顺向演替的进程,则可能会尽快使次生林恢复到合理状态。
本研究对小陇山一块长期未受人为干扰的锐齿栎天然林的群落结构和多样性进行了分析,结果表明,这块长期未受人为干扰的锐齿栎原始林的物种组成较为丰富,以地带性植被为主,锐齿栎为建群种并伴生稀有种,群落内有大量的枯立木和枯倒木;群落径级呈典型的倒J型分布,锐齿栎种群的径级分布则呈正态分布,而其他物种则多呈不规则的山状曲线。群落中乔木层树种多样性较高,树种分配均匀且优势树种集中性低,草本层和灌木层多样性低,物种分配不均匀且优势树种集中性高。赵中华等[20-22]对小陇山林区锐齿栎次生林及不同经营措施下锐齿栎次生林的物种多样性、空间结构特征、直径分布特征等进行了系统的研究,认为长期封育后锐齿栎次生林乔、灌、草各层的物种组成十分丰富,其直径分布趋向于异龄林的径级分布特征,但其相邻径级的株数分布不合理;采用大强度单株择伐的锐齿栎次生林各层次的物种组成相对较少,多样性较低,其径级结构以小径木所占的比重较大,但各径级间变幅平缓,有明显人为干扰的痕迹。与长期封育和人为干扰的锐齿栎次生林相比,本研究中的锐齿栎原始林各层次的物种数都比较丰富,直径结构明显,呈现复层、异龄、混交天然林的特征,说明该林分是长期自然演替的结果,可以代表该林区锐齿栎天然林的顶级群落特征。天然林是森林生物与环境相互依存、相互作用、长期协同进化的产物,是众多生物栖息繁衍的场所,是由丰富多彩的生物和环境组成的动态复合体。小陇山林区现有林大多是经过长期人为干扰后形成的次生林,以锐齿栎为主的松栎混交林、阔叶混交林是林区的主要林分类型。在对锐齿栎次生林进行经营时,可根据生态学原理,从优化树种组成角度出发,以长期未受干扰的天然林的结构特征为模板,通过补植、人工促进天然更新、调整林分各树种的配置等方式,使林分结构更接近于自然状态的林分结构特征,从而提升林分的整体功能。
[参考文献]
[1] 马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究:Ⅱ.丰富度、均匀度和物种多样性指数 [J].生态学报,1995,15(3):268-275.
Ma K P,Huang J H,Yu S L,et al.Plant community diversity in Dongling mountain,Beijing,China:Ⅱ.Species richness,evenness and species diversities [J].Acta Ecologica Sinica,1995,15(3):268-275.(in Chinese)
[2] 张丽霞,张 峰,上官铁梁.芦芽山植物群落的多样性研究 [J].生物多样性,2000,8(4):361-369.
Zhang L X,Zhang F,Shangguan T L.Vegetation diversity of Luya Mountains [J].Chinese Biodiversity,2000,8(4):361-369.(in Chinese)
[3] 吴 彦,刘 庆,何 海,等.亚高山针叶林人工恢复过程中物种多样性变化 [J].应用生态学报,2004,15(8):1301-1306.
Wu Y,Liu Q,He H,et al.Dynamics of species diversity in artificial restoration process of subalpine coniferous forest [J].Chinese Journal of Applied Ecology,2004,15(8):1301-1306.(in Chinese)
[4] Kvalseth T O.Note on biological diversity,evenness,and homogenety measures [J].Oikos,1991,62:123-127.
[5] McNaughton S J.Stability and biodiversity of ecological communities [J].Nature,1978,274:251-253.
[6] Odum E P.The strategy of ecosystem development [J].Science,1969,164:262-270.
[7] Margalef R.Information theory in ecology [J].General Syst,1957(3):37-71.
[8] 王树森,余新晓,班嘉蔚,等.华北土石山区天然森林植被演替中群落结构和物种多样性变化的研究 [J].水土保持研究,2006,13(6):48-50.
Wang S S,Yu X X,Ban J W,et al.Community structure and species diversity changes of forest vegetation with succession in rocky mountain area of north of China [J].Research of Soil and Water Conservation,2006,13(6):48-50.(in Chinese)
[9] 唐志尧,方精云,张 玲.秦岭太白山木本植物物种多样性的梯度格局及环境解释 [J].生物多样性,2004,12(1):115-122.
Tang Z Y,Fang J Y,Zhang L.Patterns of woody plant species diversity along environmental gradients on Mt.Taibai,Qinling Mountains [J].Chinese Biodiversity,2004,12(1):115-122.(in Chinese)
[10] 贾秀红,郑小贤.长白山过伐林区云冷杉针阔混交林空间结构分析 [J].华中农业大学学报,2006,25(4):436-440.
Jia X H,Zheng X X.Analysis on the spatial structure of the spruce-fir mixed stands of coniferous and broad-leaved trees in the over-logged forest [J].Journal of Huazhong Agricultural University,2006,25(4):436-440.(in Chinese)
[11] 安慧君,惠刚盈,郑小贤,等.不同发育阶段阔叶红松林空间结构的初步研究 [J].内蒙古大学学报,2005,36(6):714-718.
An H J,Hui G Y,Zheng X X,et al.Study on the spatial structure of broad-leaved korean pine forest in the different growth stage [J].Acta Scientiarum Naturalium Universitatis Neimongol,2005,36(6):714-718.(in Chinese)
[12] Hui G Y,Gadow K.Das winkelmass-theoretische Überlegun- gen zum optimalen standardwinkel [J].Allgemeine Forst und Jagdzeitung,2002,173(9):173-176.
[13] 郑丽凤,周新年,江希钿,等.松阔混交林林分空间结构分析 [J].热带亚热带植物学报,2006,14(4):275-280.
Zheng L F,Zhou X N,Jiang X D,et al.Analysis of the stand spatial structure ofPinusmassoniana-broadleaved mixed forest [J].Journal of Tropical and Subtropical Botany,2006,14(4):275-280.(in Chinese)
[14] Gadow K.Zur bestandesbeschreibung in der foresteinrichtung [J].Forst und Holz,1993,48(21):602-606.
[15] 惠刚盈,克劳斯·冯佳多.森林空间结构量化分析方法 [M].北京:中国科学技术出版社,2003.
Hui G Y,Klaus von G.Quantitative analysis of forest spatial structure [M].Beijing:China Science and Technology Press,2003.(in Chinese)
[16] 孙儒泳.普通生态学 [M].北京:高等教育出版社,1993.
Sun R Y.General ecology [M].Beijing:Higher Education Press,1993.(in Chinese)
[17] Gadow K,Füldner K.Zur methodik der bestandesbeschreibung [R].Klieken:Vortrag Anlaesslich der Jahrestagung der A G Forsteinrichtung,1992.
[18] 惠刚盈,Klaus von G,胡艳波,等.结构化森林经营 [M].北京:中国林业出版社,2007.
Hui G Y,Klaus von G,Hu Y B,et al.Tructure-based forest management [M].Beijing:China Forestry Press,2007.(in Chinese)
[19] 朱教君.次生林经营基础研究进展 [J].应用生态学报,2002,13(12):1689-1694.
Zhu J J.A review on fundamental studies of secondary forest management [J].Chinese Journal of Applied Ecology,2002,13(12):1689-1694.(in Chinese)
[20] 赵中华,惠刚盈,袁士云,等.小陇山锐齿栎天然林空间结构特征 [J].林业科学,2009,45(3):1-6.
Zhao Z H,Hui G Y,Yuan S Y,et al.Spatial structure characteristic ofQuercusalienavar.acuteserratanatural forest in Xiaolongshan [J].Scientia Silvae Sinicae,2009,45(3):1-6.(in Chinese)
[21] 赵中华,白登忠,惠刚盈,等.小陇山不同经营措施下次生锐齿栎天然林物种多样性研究 [J].林业科学研究,2013(3):326-331.
Zhao Z H,Bai D Z,Hui G Y,et al.Species diversity ofQuercusalienavar.acuteserratanatural forest with different management measures at Xiaolongshan [J].Forest Research,2013(3):326-331.(in Chinese)
[22] 赵中华,袁士云,惠刚盈,等.甘肃小陇山5种不同灌木林改造模式对比分析 [J].林业科学研究,2008(2):262-267.
Zhao Z H,Yuan S Y,Hui G Y,et al.Comparative analysis of five shrub forest transformation models on Xiaolongshan in Gansu [J].Forest Research,2008(2):262-267.(in Chinese)
