陕西豌豆种质资源形态性状遗传多样性分析
2014-03-26刘萌娟李鸣雷郭小华王静雅宋卫宁
刘萌娟,李鸣雷,郭小华,王静雅,宋卫宁
(1 西北农林科技大学 a 农学院,b 水土保持研究所, 陕西 杨凌 712100;2 连云港市国欣种业有限公司, 江苏 连云港 222000)
豌豆(PisumsativumLinn)是豆科(Leguminosae)豌豆属(PisumL.)植物中的一个栽培种,染色体数为2n=14,为1年或越年生攀缘性作物。豌豆起源于地中海沿岸地区及非洲北部、亚洲西部等地,栽培驯化的历史在6 000年以上,豌豆在中国栽培有2 000多年[1], 在豆类作物中,豌豆的种植面积和产量仅次于大豆、花生和普通菜豆[2]。据联合国粮农组织统计,2012年全世界前20个生产干豌豆的国家,总产量781.3万t,总产值11.4亿美元;前20个生产青豌豆的国家,总产量1 778.7万t[2],总产值达到58.9亿美元,其中中国大陆青豌豆的总产量和产值分别是1 150万t和38.1亿美元[3],均占到全世界20个青豌豆生产国产量和产值总和的64%。我国从1978年开始有计划有组织地进行豌豆种质资源的研究,到1990年底,经初步鉴定编目的豌豆资源有2 616份,其中陕西省搜集和保存的豌豆种质资源358份,占国内保存豌豆种质资源的 13.7%,这些资源以陕西地方品种为主,少数是来自澳大利亚、法国、英国、日本等国家的种质[4-6]。
关于作物品种资源遗传多样性的研究,目前主要采用分子标记和表型性状的研究方法。在豌豆分子标记遗传多样性研究方面,Choudhury等[7]以来源于印度的24个豌豆品种为材料,利用60个RAPD引物区分了高秆和矮秆豌豆品种。Yadav等[8]利用11对RAPD引物,结合种子特征和地理来源对15份豌豆材料(印度14份、英国1份)进行了遗传多样性研究,结果表明15份豌豆间的相似系数为0.263~0.793,说明15份豌豆材料间具有较高的遗传多样性。Baranger等[9]利用同工酶、贮藏蛋白、RAPD、ISSR、SSR和STS标记,对来自西欧的148份豌豆栽培资源和育成品种进行遗传多样性分析,发现按用途区分的栽培豌豆类型间存在明显差别,相同系谱来源的育成品种聚为一类,并在聚类基础上筛选出由43份参试材料组成的核心种质,其含有所有参试材料96%的等位变异。Burstin等[10]根据43对SSR标记引物,对12个豌豆品系进行分析,检测到31个等位变异。宗绪晓等[11]对来自我国19个省区的1 221份豌豆地方品种,通过SSR标记进行遗传多样性研究,将中国豌豆地方品种分化为3个基因库,其中基因库Ⅰ主要由春播区的内蒙古、陕西资源构成,基因库Ⅱ主要由秋播区最北端的河南资源构成,基因库Ⅲ主要由除上述省份之外的其他省区市的资源构成,中国豌豆地方品种资源群体间的遗传距离与其来源地的生态环境相关。作为揭示植物不同层次上的遗传变异的方法,分子标记和表型性状各有优缺点,是不可互相代替的。农艺性状具有简单、方便、直观的特点,是研究作物遗传多样性的一种重要手段。李玲等[12]对642份国内豌豆种质资源的26个形态性状进行鉴定,结果表明,国内豌豆种质资源具有丰富的形态多样性,平均多样性指数为0.895;基于形态性状,将642份豌豆种质划分为14大组群,聚类结果显示豌豆品种间的遗传距离与地理距离有一定相关。贺晨帮等[13]针对国内外不同地理来源的624份豌豆资源,研究了其20个性状的平均值、变异系数、遗传多样性指数,结果表明,国内外不同地理来源豌豆资源群体间的遗传变异大,参试资源由国内和国外两大基因库组成,证明基于形态性状得到的遗传多样性分析结果同样可靠。
陕西地域南北长,东西窄,南北长约880 km,东西宽160~490 km;陕西地势总体呈南北高、中部低。南北区域的地形、地势、气候等因素差异很大,形成了丰富多样的豌豆品种资源。除了位于渭北和陕北的春播豌豆以外,在关中平原和陕南的秦岭、巴山山区还存在着大量的秋播豌豆资源。然而目前有关陕西豌豆资源的遗传多样性研究尚未见报道。本试验以陕西保存豌豆资源中的100份种质(含省内及来自省外的资源)为材料,分析其生育期、株高、单株荚数、单荚粒数、百粒质量、单株产量、花色、粒形和粒色9个形态性状的分布特点,研究陕西豌豆品种资源的遗传多样性及地理分布特点,以期为豌豆种质资源的保护、研究和利用提供理论依据。
1 材料与方法
1.1 试验材料
参考《中国食用豆类品种资源目录》,从陕西保存的豌豆资源中,根据生育期、花色、粒形、粒色性状不同选取100份种质,其中陕北、关中以及陕西省外各20份,陕南40份;按陕北(N01~N20)、关中(M01~M20)、陕南(S01~S40)、省外(F01~F20)分别编号,详见表1。

续表1 Continued table 1
1.2 试验设计
本研究是结合国家中期库种质资源保种项目进行的。全部繁种的豌豆品种资源于2003-2005年种植于西北农林科技大学南校区试验地。田间种植采用四行区顺序排列,行长2 m,行距0.6 m,每小区4行,小区面积4.8 m2。播期为10月中旬,收获期在翌年的6月上中旬,田间管理同大田。
1.3 性状调查
田间调查出苗期、花色、成熟期,计算生育期;收获后随机取连续10株考种,统计株高、单株产量、单荚粒数、单株粒数、百粒质量,取2年的平均值,并记载种子的粒形和粒色性状。
1.4 数据处理
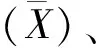
遗传多样性指数(Shannon-Weaver index,H′)的计算公式如下:
H′=-∑PilnPi。
式中:Pi为某性状第i级别内材料份数占总份数的比例。
对花色、粒色、粒形3个质量性状进行赋值(花色:白=1,红(包括粉红)=2,紫=3;粒色:白=1,粉红=2,浅绿=3,绿=4,灰绿=5,褐=6,麻=7;粒形:圆粒=1,皱粒=2),计算各级性状的相对频率及该性状的遗传多样性指数(H′)。
将上述3个质量性状赋值转化为数量性状后,与6个数量性状一起构成数据阵。对数据进行标准化处理,用欧氏距离平方(Squared euclidean distance)计算成对品种间的遗传距离(D)。
D=∑(xi-yi)2。
式中:x、y代表任意成对的品种,i代表不同的变量(性状)值。
同时计算不同生态区内部以及生态区之间品种的遗传距离,对遗传距离进行方差分析和多重比较。
用Ward法、Spss统计软件对供试的100份豌豆种质进行聚类,建立树状聚类图。对标准化后的数据阵进行主成分分析,取前3个主成分,根据每个品种在前3个主成分上的取值,利用Spss软件绘制品种的三维散点图。
2 结果与分析
2.1 豌豆形态性状在不同地区的分布
2.1.1 花 色 豌豆的花色有白、紫和红3种类型,其中红为稀有类型。从图1可以看出,陕北、关中、陕南3个地区紫花豌豆所占的比例大于白花豌豆,红花豌豆所占的比例最小。陕西豌豆品种以紫花为主,省外豌豆品种则以白花为主。红花豌豆在各地区的比例都比较低,关中和陕南各有1份,省外材料中有3份,陕北则没有红花豌豆。
2.1.2 粒 色 豌豆的粒色有白、粉红、浅绿、灰绿、绿、褐、麻7种类型。从图2可以看出,陕西豌豆具有全部7种粒色类型,省外豌豆具有6种粒色类型(没有灰绿粒色)。陕南具有全部7种粒色类型,关中和陕北地区都只有4种粒色类型,说明陕南豌豆籽粒形态丰富。不论陕西还是省外豌豆主要以白和麻两色为主,陕北和关中麻豌豆多于白豌豆,陕南的麻豌豆略低于白豌豆,而省外是白豌豆明显多于麻豌豆,这可能与有目的地选取省外优质资源有关。麻豌豆多作为粮用或饲料用作物,较能适应干旱和瘠薄的土壤条件,关中虽然水肥条件好,但是豌豆作为一种小杂粮作物,常被种植于瘠薄和干旱的土地;白豌豆以及绿豌豆(包括浅绿和灰绿豌豆)、粉红豌豆多作菜用,要求种植在水肥条件好的地块,所以在陕南的比例相对较高。豌豆粒色的分布规律反映了品种随环境变化的特点以及人为选择对品种类型的影响。
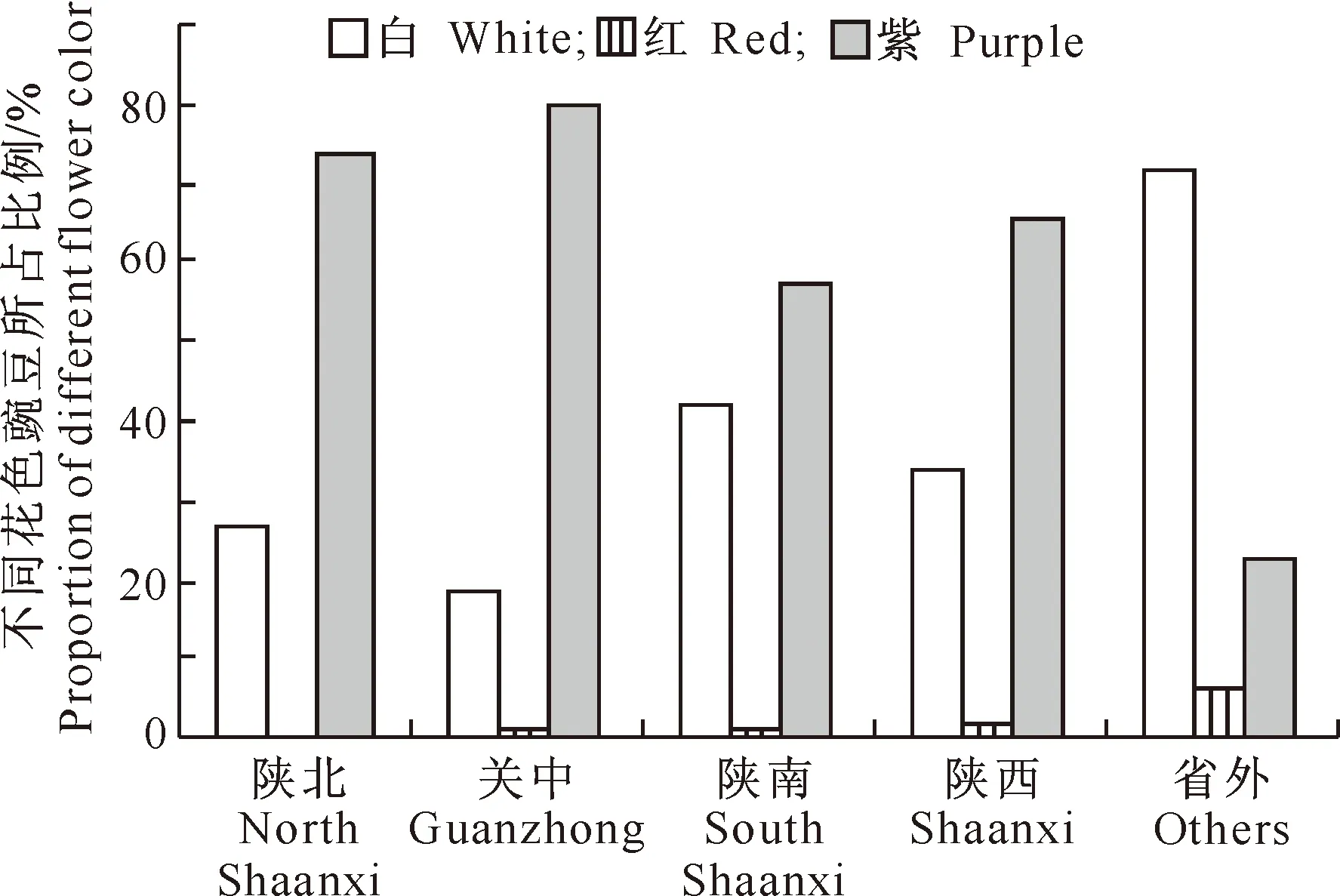
图1 不同花色豌豆在地区间的分布
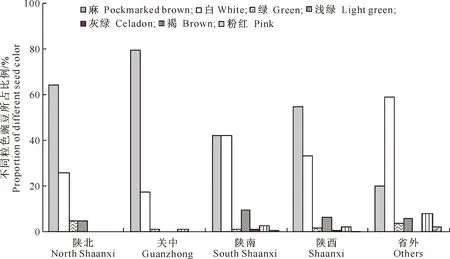
图2 不同粒色豌豆在地区间的分布
2.1.3 粒 形 豌豆的粒形分为圆粒和皱粒2种。从图3可以看出,在陕西和省外,圆粒豌豆所占比例分别为89.67%和75.51%。在陕西省内,从陕北、关中到陕南,圆粒豌豆的比例依次下降,皱粒豌豆的比例依次上升,同时省外皱粒豌豆的比例高于省内皱粒豌豆比例最高的陕南地区。皱粒品种是由于种子内可溶性糖含量高,在成熟期失水收缩形成的,口感甜脆,食用品质好,是一种鲜食菜用的类型。陕西豌豆的粒形分布特点反映了从北到南粮用或饲料用豌豆品种减少、菜用品种增加的趋势。较之陕西豌豆,省外皱粒豌豆比例较大反映了省外豌豆菜用品种较多的特点,同时也可能与早期资源研究中有目的地选取省外优质资源有关。

图3 不同粒形豌豆在地区间的分布
2.1.4 不同生态区豌豆数量性状的比较 由表2可以看出,不同生态区豌豆品种数量性状间的变异系数相差很大。生育期和株高2个性状的变异系数较小,多在15%以下,并且省内的变异系数小于省外。单株荚数、单荚粒数、百粒质量和单株产量4个性状的变异系数较大,多数在30%以上,省内品种的变异系数大于省外品种。不同性状间的变异系数相差很大,表明豌豆品种不同性状的分化程度有很大差异。单株荚数、单荚粒数、百粒质量和单株产量4个性状变异幅度较大,易于通过选择达到育种目标;而生育期和株高的变异幅度较小,通过选择达到育种目标的可能性较小。与省内豌豆种质相比,省外资源表现为生育期缩短,株高降低,单株荚数、百粒质量增加以及单株产量升高,说明省外豌豆资源的农艺性状优于省内种质。
2.2 不同生态区豌豆性状的遗传多样性指数
由表3 可以看出,豌豆不同性状的遗传多样性指数有很大差异, 6个数量性状的遗传多样性高于3个质量性状。生育期和单荚粒数的遗传多样性指数较低,其余4个数量性状的遗传多样性指数较高,但是不同地区之间有差异。质量性状中粒色的遗传多样性指数明显高于粒形和花色,并且在不同地区间差异较大。陕西豌豆9个性状的遗传多样性指数在0.422(粒形)~2.039(株高),省外豌豆的遗传多样性在0.557(粒形)~2.101(株高)。

表2 不同生态区豌豆数量性状的比较
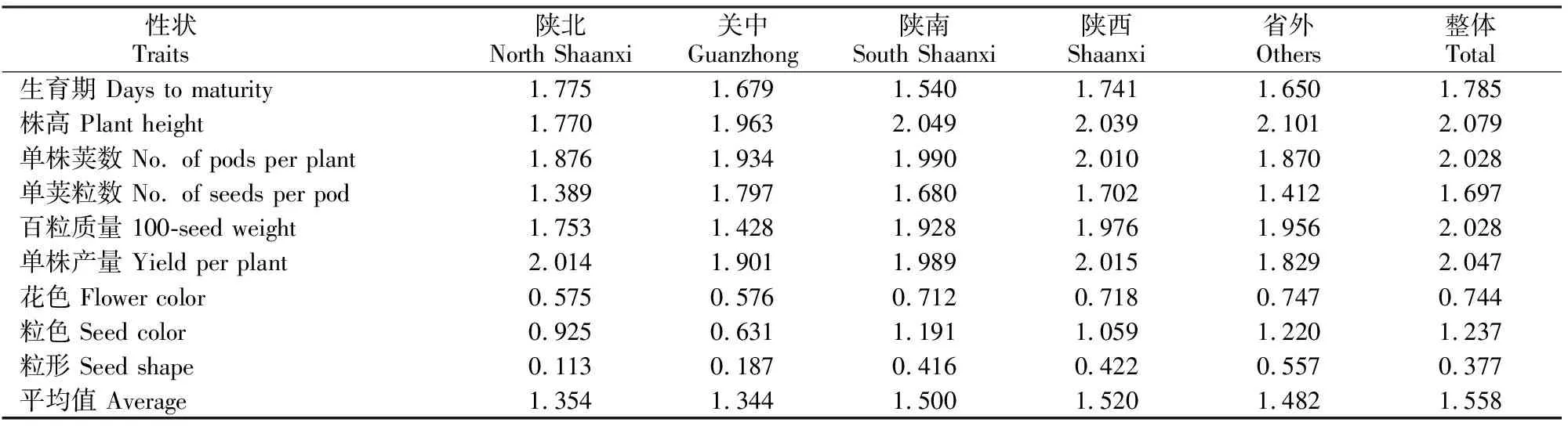
表3 不同生态区豌豆性状的遗传多样性指数
除单荚粒数和粒形以外,100份种质整体的平均遗传多样性指数高于各地区的品种群体,株高、单株荚数、百粒质量、单株产量、粒色的遗传多样性指数较高;陕西品种的单株荚数、百粒质量、单株产量、花色、粒形的平均遗传多样性指数高于省内不同地区。说明随着群体中个体数量的增多,群体的遗传多样性相应地也会增加,但是如果性状的不同级别在群体中分布不均匀,反而会降低群体的遗传多样性。如,粒形的遗传多样性指数整体明显低于陕南、陕西以及省外,但由于关中和陕北豌豆品种以圆粒种质为主,最终导致粒形整体分布不均匀。
陕西豌豆的平均遗传多样性指数高于省外群体。陕西豌豆除株高以外的生育期、单株荚数、单荚粒数、百粒质量和单株产量5个数量性状的平均遗传多样性指数均高于省外群体,而花色、粒色和粒形3个质量性状的遗传多样性指数却低于省外群体。省内不同地区间的平均遗传多样性指数以陕南为高,其次是陕北、关中;9个性状中有6个性状的遗传多样性指数以陕南最高,其次是陕北2个(生育期、单株产量),关中1个(单荚粒数)。与省外群体相比,陕南群体仅3个性状(单株荚数、单荚粒数、单株产量)的遗传多样性指数高于省外,但其平均遗传多样性指数略高于省外群体,显示陕南群体遗传多样性水平较高。
2.3 不同生态区豌豆的遗传距离
100份豌豆种质间的遗传距离为0.07~67.19,平均遗传距离为18.00,标准差为10.86。由表4可以看出,不同地区豌豆品种间遗传距离不同,省外品种群体的平均遗传距离最大,为20.27,省内品种依次分别为陕南(17.98)、关中(17.34)、陕北(11.97)。关中与陕南群体平均遗传距离差异不显著,关中与陕北群体及陕南与陕北群体的平均遗传距离差异极显著;省外与陕南群体的平均遗传距离达到差异显著,省外群体与关中、陕北群体的平均遗传距离差异极显著。省外群体与陕西3个地区之间的平均遗传距离都大于陕西3个地区之间的平均遗传距离;而陕北与陕西其他2个地区品种群体的遗传距离则小于其他3个地区群体间的平均遗传距离;关中和陕南群体与其他3个地区品种群体的遗传距离则较为接近且居中。
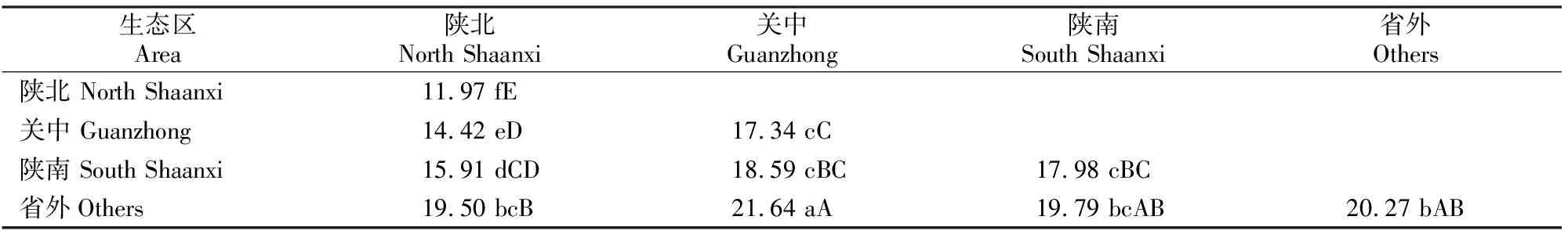
表4 不同生态区内及区间品种的平均遗传距离
2.4 不同生态区豌豆种质的聚类分析和三维散点图
由图4可见,依据遗传距离将100份参试豌豆材料分成2类,第Ⅰ类由51份材料组成,包括17份省外品种、23份陕南品种、6份关中品种和5份陕北品种;第Ⅱ类由其余的49份材料组成,包括3份省外品种、17份陕南品种、14份关中品种和15份陕北品种。进一步分析发现,第Ⅰ类可以分成3个亚组,其中第1亚组由34份材料组成,包含11份省外品种、13份陕南品种和关中、陕北品种各5份;第2亚组由5份材料组成,分别为省外品种3份和关中、陕南品种各1份;第3亚组由12份材料组成,即省外品种3份和陕南品种9份。第Ⅱ类可以进一步划分为2个亚组,第1亚组含有17份材料,其中省外品种1份、陕南品种4份、关中品种8份、陕北品种4份;第2亚组含有32份材料,其中省外品种2份、陕南品种13份、关中品种6份、陕北品种11份。
由此可见,陕南、省外品种主要聚在第Ⅰ类,分别占到各自群体的57.5%和85%;关中、陕北品种主要聚在第Ⅱ类,分别占各自群体的70%和75%,说明聚类分析反映了品种的地理来源。陕南和省外的品种散布在5个亚组中,而关中、陕北的品种只分布到其中的部分亚组中,说明前者的分布更为广泛。与省外品种相比,陕南品种分布更加均衡,主要出现在Ⅰ-1、Ⅰ-3和Ⅱ-2亚组中,而省外品种主要出现在Ⅰ-1亚组中。与陕北品种相比,关中品种分布较为广泛,出现在聚类形成的4个亚组中,并且以Ⅰ-1、Ⅱ-1和Ⅱ-2亚组为主;而陕北品种只出现在聚类形成的3个亚组中,以Ⅱ-2类为最多。从聚类分析结果来看,陕南品种的多样性最高,其次分别为省外、关中和陕北,这与遗传多样性指数反映的顺序基本一致,同时聚类分析也显现出了一定的品种地理分布特征。

图4 不同生态区豌豆资源的聚类结果
依据主成分分析结果绘制的散点图(图5)显示了品种间的三维空间关系,形象地解释了品种间的遗传距离以及聚类分析的结果。第Ⅰ主成分解释了总变异的27.45%,第Ⅱ主成分解释了总变异的22.21%,第Ⅲ主成分解释了总变异的15.96%,三者之和达到了65.61%。图5结果表明,省外品种最为分散,其次是陕南品种,而关中尤其是陕北的品种最为接近,反映了不同地区内部品种间的遗传距离关系;省外品种与陕南品种距离较近,陕北品种与关中品种距离较近;不同地区的品种没有形成相对独立的区域,而是相互穿插,但是同一地区的品种还是首先集中在一起,具有较为明显的分布区域,与聚类分析的结果相吻合。
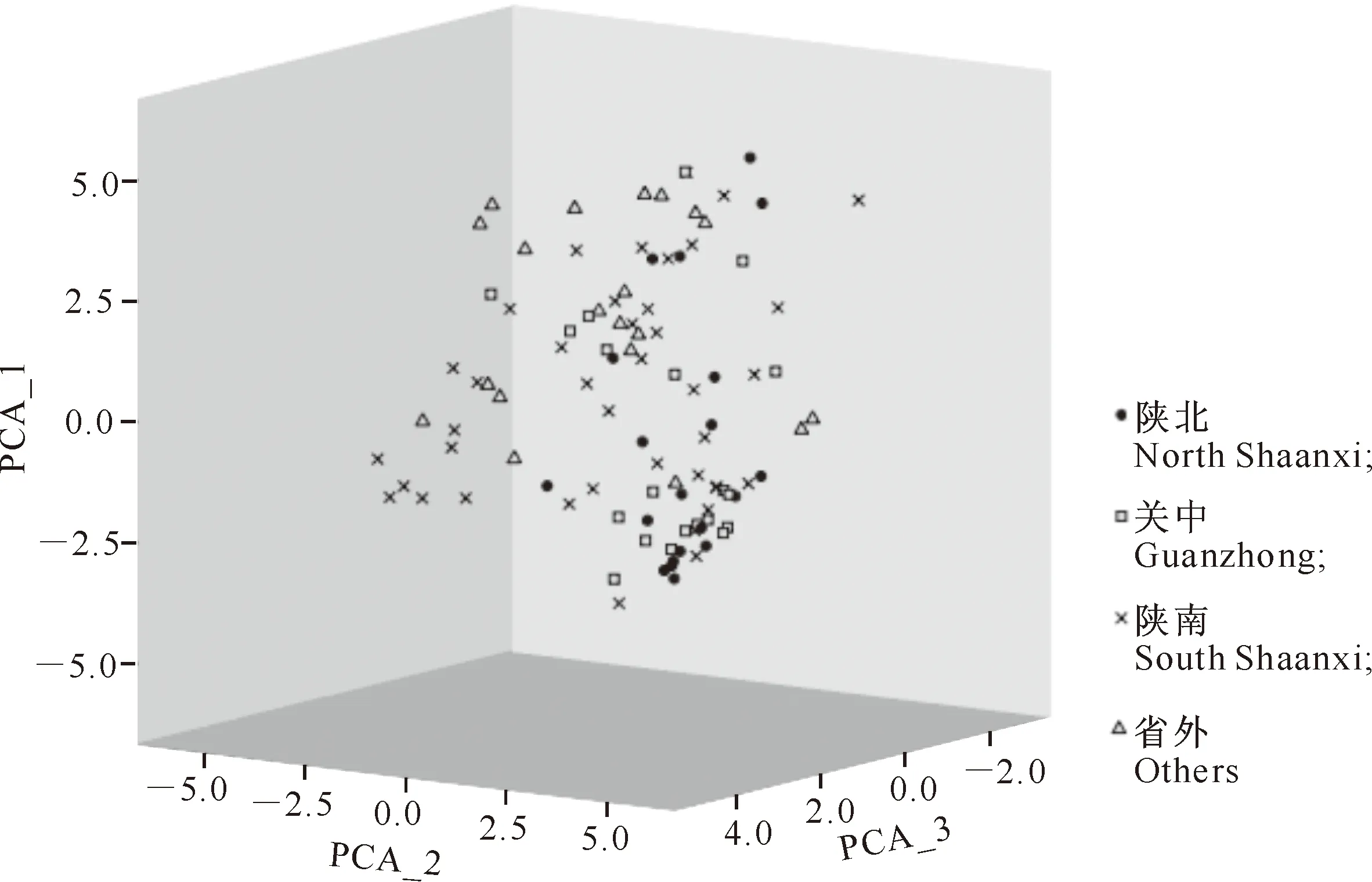
图5 不同生态区豌豆品种分布的三维散点图
3 讨 论
3.1 补充省外豌豆资源的意义
陕西省外豌豆资源中白花、白粒豌豆以及皱粒豌豆的比例明显高于省内,说明省外豌豆的籽粒性状优于省内豌豆;同时,省外豌豆资源的株高降低,生育期缩短,单株荚数、百粒质量增加,单株产量升高;省外豌豆9个性状的平均遗传多样性指数略低于全省和陕南,明显高于关中和陕北,说明由于省外资源的加入使整体资源的平均遗传多样性指数增大。省外群体的平均遗传距离大于省内群体,而省外群体与省内任一地区之间的平均遗传距离都大于陕西3个地区之间的平均遗传距离。以上分析表明,省外豌豆资源的农艺性状优于省内资源,多样性丰富;同时省外豌豆群体的遗传差异大,并且与陕西豌豆群体的遗传差异也大,说明省外豌豆是陕西省内豌豆种质资源的重要补充,应该在种质资源筛选和育种中充分利用。
3.2 陕西豌豆的生态适应性和遗传多样性
省内豌豆在单株荚数、单荚粒数和百粒质量3个性状上表现出一定的增减规律,从陕北到陕南单株荚数、单荚粒数减少,百粒质量增加,这与陕西从北到南的气候生态特点是相适应的,陕北地区干旱少雨、灌溉条件差;关中地区降雨基本能满足农业生产的要求,地势平坦,灌溉方便,水分因素不再是农业生产的限制因素;陕南地区湿润温暖、降水充足。充足的水分和光热资源促使陕南豌豆容易形成大粒品种,而在陕北,因为干旱的限制促使豌豆形成了较小的籽粒和较多的单株粒数,或者,只有较小的籽粒、较多单株粒数的品种类型才能生存下来。
本研究结果显示,从陕北到陕南豌豆的遗传多样性在增加。首先,不同地区拥有的资源数量明显地反映了这一特点。虽然不同性状的遗传多样性指数变化趋势有所不同,但是多数性状的遗传多样性指数以及平均值都表现为从陕北到陕南增加的趋势。至于群体内部品种间的遗传距离,也是以陕南为最大,然后依次是关中、陕北。聚类结果显示,陕南品种的分布最为广泛,分布到聚类形成的5个亚组中,其次是关中、陕北品种。遗传多样性从北到南依次增大的特点,与陕西从北到南地形、地貌、气候的变化是一致的。陕西南北狭长,陕南主要是由秦岭山脉和巴山山脉组成的山区和丘陵河谷地带,气候温暖湿润,海拔和地形变化大、交通不便,形成了许多相对孤立的小流域和小气候,为生物多样性的产生、积累和保存创造了条件。陕北地区是广袤的黄土高原,深厚的土层被纵横交错的河道切割成大小不同的沟、峁、梁地貌,气候干燥、寒冷。由于环境严酷,只有少数耐旱性、耐寒性强的品种才能生存下来,导致陕北品种生物多样性减少。这与对陕西大豆资源的研究结果[14-15]有所不同,可能与采取的研究方法不同有关;而与陕南品种数量多、遗传多样性指数高于陕北和关中地区的研究结果一致,两者都表明陕南品种存在许多广泛而微小的变异。说明陕南地区气候环境特点有利于物种生物多样性的产生、积累和保存。因此,应该加强对陕南秦岭山区生物多样性聚集地的保护。
3.3 豌豆资源多样性与地理来源的相关性
关于种质资源遗传多样性分布与地理来源的关系,不同的研究往往给出不同的结果。研究发现,中国豌豆地方资源分化形成3个富集区,中国资源和外国资源属于明显不同的基因库,豌豆资源的多样性分布规律与品种的地理来源相关明显[11]。李玲等[12]对642份国内豌豆农艺性状的研究结果表明,品种之间的遗传距离与地理距离不完全一致,表现为有些地理来源相同的品种分布在不同的组中,地理来源不同的品种分布在相同的组中。Gemechu等[16]通过对埃塞俄比亚148份豌豆地方品种12个农艺性状的多样性分析发现,埃塞俄比亚豌豆地方品种具有较高的遗传多样性,但其多样性分布规律与地理来源没有明显的关联。本研究结果与李玲等[12]和Gemechu等[16]的结论相似:聚类分析和三维散点图都表明,虽然同一地区的品种有成片聚集的趋势,但是没有形成由同一地区品种聚集而成的相对集中的区域或是亚组,不同地区的品种常常穿插在一起。省内群体之间品种间的平均遗传距离,以陕南-关中为最大,然后依次是陕南-陕北、关中-陕北。地理距离相对较近的关中-陕南之间却具有最大的遗传距离,显示陕西豌豆的遗传距离与地理距离没有明显的相关,陕西豌豆多样性分布规律与地理来源的关联不明显。
4 结 论
陕西豌豆的形态以紫花、麻粒和白粒、圆粒性状为主,从陕北到陕南皱粒性状比例增加,粒色类型更加丰富;省外豌豆以白花、白粒为主,除了白粒和麻粒以外的其他颜色豌豆也占有较大比重,皱粒豌豆的比例明显高于陕西豌豆。省内豌豆和省外豌豆在多数数量性状上存在差异,与陕西豌豆相比,省外资源生育期缩短,株高降低,单株荚数、百粒质量、单株产量升高,省外资源的农艺性状优于省内资源。省内豌豆在单株荚数、单荚粒数和百粒质量3个性状上,不同地区间的差异显著,从陕北到陕南单株荚数、单荚粒数减少,百粒质量表现为陕南>陕北>关中。
陕西豌豆群体的平均遗传多样性高于省外群体,省内不同地区间的平均遗传多样性指数以陕南为高,其次是陕北、关中,显示陕南群体具有较高的遗传多样性水平。
省内群体的平均遗传距离以陕南为最大,其次分别是关中、陕北;省内不同群体间的平均遗传距离,以陕南-关中为最大,其次分别是陕南-陕北、关中-陕北。省外群体的平均遗传距离大于任一省内群体,而省外群体与省内地区间的平均遗传距离都大于陕西3个地区之间的平均遗传距离。
聚类分析将100份豌豆材料分成2类,第Ⅰ类主要由陕南、省外品种组成,第Ⅱ类主要由关中、陕北品种组成。聚类和PCA分析结果显示,陕南品种分布最为广泛,其次分别是省外、关中、陕北的品种,不同地区的品种没有形成相对独立的区域,而是相互穿插,界限模糊。
[参考文献]
[1] 龙静宜,林黎奋,侯修身,等.食用豆类作物 [M].北京:科学出版社,1989.
Long J Y,Lin L F,Hou X S,et al.Legume crops [M].Beijing:Science Press,1989.(in Chinese)
[2] 郑卓杰,王述民,宗绪晓.中国食用豆类学 [M].北京:中国农业出版社,1997:93-96.
Zheng Z J,Wang S M,Zong X X.Food legume crops in China [M].Beijing:China Agriculture Press,1997:93-96.(in Chinese)
[3] FAO.Food and agricultural commodities production [DB/OL].http://faostat.fao.org/site/339/default.aspx.2014.
[4] 郑卓杰,王景月,胡家蓬,等.中国食用豆类品种资源目录(第一集) [M].北京: 中国农业科技出版社,1987:386-396,418.
Zheng Z J,Wang J Y,Hu J P,et al.Catalog of Chinese food legume variety resources (Ⅰ) [M].Beijing:Chinese Press of Agricultural Science and Technique,1987:386-396,418.(in Chinese)
[5] 郑卓杰,胡家蓬,于增俊,等.中国食用豆类品种资源目录(第二集) [M].北京:中国农业出版社,1990:280-288.
Zheng Z J,Hu J P,Yu Z J,et al.Catalog of Chinese food legume variety resources (Ⅱ) [M].Beijing:China Agriculture Press,1990:280-288.(in Chinese)
[6] 胡家蓬,程须珍,王佩芝,等.中国食用豆类品种资源目录(第三集) [M].北京:中国农业出版社,1996:222.
Hu J P,Cheng X Z,Wang P Z,et al.Catalog of Chinese food legume variety resources (Ⅲ) [M].Beijing:China Agriculture Press,1996:222.(in Chinese)
[7] Choudhury P R,Tanveer H,Dixit G P.Identification and detection of genetic relatedness among important varieties of pea (PisumsativumL.) grown in India [J].Genetica,2007,130:189-191.
[8] Yadav V K,Kumar S,Panwar R K.Measurement of genetic di-ssimilarity in world pea (PisumsativumL.) genotypes using RAPD markers [J].Genetic Resources and Crop Evolution,2007,54(6):1285-1289.
[9] Baranger A G,Aubert G,Arnau G,et al.Genetic diversity withinPisumsativumusing protein- and PCR-based markers [J].Theoretical and Applied Genetics,2004,408(7):1309-1321.
[10] Burstin J,Deniot G,Potier J,et al.Microsatellite polymorphism inPisumsativum[J].Plant Breeding,2001,120(4):311-317.
[11] 宗绪晓,关建平,王述民,等.中国豌豆地方品种SSR标记遗传多样性分析 [J].作物学报,2008,34(8):1330-1338.
Zong X X,Guan J P,Wang S M,et al.Genetic diversity among Chinese pea (PisumsativumL.) landraces revealed by SSR markers [J].Acta Agronomica Sinica,2008,34(8):1330-1338.(in Chinese)
[12] 李 玲,孙文松.国内豌豆种质资源的形态多样性分析 [J].辽宁农业科学,2010(2):22-25.
Li L,Sun W S.Morphological diversity analysis of Chinese pea (PisumsativumL.) germplasm [J].Liaoning Agricultural Sciences,2010(2):22-25.(in Chinese)
[13] 贺晨帮,宗绪晓.豌豆种质资源形态标记遗传多样性分析 [J].植物遗传资源学报,2011,12(1):42-48.
He C B,Zong X X.Genetic diversity of pea (PisumsativumL.) resources revealed by morphological traits [J].Journal of Plant Genetic Resources,2011,12(1):42-48.(in Chinese)
[14] 刘萌娟,李鸣雷,赵惠贤,等.陕西大豆资源遗传多样性及变异特点研究 [J].植物遗传资源学报,2010,11(3):326-334.
Liu M J,Li M L,Zhao H X,et al.Genetic diversity and variation trait of Shaanxi soybean landraces [J].Journal of Plant Genetic Resources,2010,11(3):326-334.(in Chinese)
[15] 刘萌娟,胡胜武,赵惠贤,等.基于农艺性状和RAPD片段的陕西大豆种质资源遗传多样性研究 [J].西北农业学报,2009,18(5):159-166.
Liu M J,Hu S W,Zhao H X,et al.Genetic diversity of Shaanxi soybean landrace based on agronomic traits and RAPD data [J].Acta Agriculturae Boreali-Occidentalis Sinica,2009,18(5):159-166.(in Chinese)
[16] Gemechu K,Mussa J,Getnet D. Extent and pattern of gene-tic diversity for morpho-agronomic traits in Ethiopian highland pulse landraces:Ⅰ.Field pea (PisumsativumL.) [J].Genetic Resources and Crop Evolution,2005,52:539-549.
