无患子实生群体种实表型性状变异研究
2014-03-25刁松锋邵文豪姜景民董汝湘孙洪刚
刁松锋,邵文豪,姜景民,董汝湘,孙洪刚
(中国林业科学研究院 亚热带林业研究所,浙江 杭州 311400)
表型指基因型在个体水平上的表达,是基因型和环境因素互作的结果。表型性状是检测遗传变异最直接和最简便的方法。形态学性状是表型的一部分,表型性状还包括生物化学成分特征、生化代谢特征、生理活性特征及环境适应能力等。通常利用的表型性状有2类:一类是符合孟德尔遗传规律的单基因性状(质量性状、稀有突变等),另一类是由多基因决定的数量性状。在天然和实生种群中,单基因性状较少,故用其作为遗传标记来研究遗传多样性,目前多见于对一些有重要经济价值的农作物、林木、园艺作物及其野生近缘种的研究[1-2]之中。对于植物表型性状的研究主要是从2个方面展开的:一个是从遗传上进行研究,其对象多为重要的经济性状,侧重于寻找具优良性状的遗传材料,为遗传改良奠定基础;另一个是从生态上开展研究,其对象主要是在生态上十分重要的性状,注重探究物种的适应性[3]。植物的表型性状包括种实、花、叶及树体性状等,但对于经济林产果类植物而言,主要表型性状体现为种实性状。种实性状是植物有性生殖系统的重要组成部分,是生存和繁衍适应性的集中表现[4]。
无患子(SapindusmukorossiGaertn.)又名肥皂树、洗手果等,为无患子科(Sapindaceae)无患子属落叶大乔木,大型圆锥花序顶生,花小量多,花期5-6月;果实多近圆球形,熟时橙黄色,干时褐色,果期9-11月[5]。无患子立地适应能力强,广泛分布在亚洲和美洲的热带、亚热带低山丘陵及石灰岩地区[6]。无患子果皮皂苷含量丰富,高达10.76%[7],具有良好的起泡性和去污能力,是天然的优良洗涤剂[8]。无患子种仁含油率高,可达42.7%[9],是开发生物燃料,应对能源危机的理想原料[10]。由于无患子具有开发生物柴油的巨大潜力,国家相关部委已将其列为重要的生物质能源树种。近年来,随着生物质产业的发展,无患子作为新兴工业原料树种越来越受到关注,从事无患子大规模种植的企业也相继增加。因此,为规范无患子原料林的培育利用活动,国家林业局印发了《无患子原料林可持续培育指南》([2012]129号)[11]。
目前,有关无患子的研究和开发利用尚处于起步阶段,研究内容多集中在园林绿化[12]、繁育[13]、化学成分及其提取工艺[9,14]和医药[15]等方面,而对无患子种实表型性状的变异规律尚未见报道。为此,本试验以无患子实生群体种实表型性状为研究对象,探讨无患子表型性状特别是主要经济性状的变异规律,以期为后续无患子优树选择、高产果用林的营建提供理论基础,并为以无患子果实为原料的新兴产业发展奠定基础。
1 材料与方法
1.1 试验地概况
试验材料来自浙江省天台县满园春农林开发有限公司无患子原料林基地,该基地为利用采自本地无患子老树种子育苗营建的8年生无患子实生林,采种树约75株,胸径19.1~35.3 m,树高12.4~21.6 m,树龄不详,但根据对村民的查访树龄大约为30年。基地面积约100 hm2,处于北纬28°57′02″、东经120°41′24″,海拔55~95 m,黏性红黄壤,土层厚度20~70 cm,pH=5.3~6.0;年平均气温16.8 ℃,降雨量1 320 mm,1和7月份平均温度分别5.1和28.5 ℃,年无霜期平均232 d。由于海拔差异不大、地势较为平坦、林分立地条件相差不大,林下植被植物多以络石(Trachelospermumjasminoides)、小果蔷薇(Rosacymosa)、山莓(Rubuscorchorifolius)、铁芒萁(Dicranopterislinearis)、枸骨(Ilexcornuta)等为主。2012年,该林分郁闭度约为75%,株行距4 m×4 m。
1.2 无患子种实性状及其测量方法
2011-11,从全林分中随机选择102株生长健康的无患子单株,株间水平距离5倍株高以上,以保证取样的均匀性。每个采样植株树体均涂刷红漆并挂牌标记以便于跟踪调查。2012-12,重复调查2011年选择的102棵单株。选择遗传相对稳定、易于获得和测定的表型性状,包括单株产量(Yield per plant,YPP,kg)、果实质量(Fruit weight,FW,g)、果实纵径(Fruit vertical diameter,FVD,mm)、果实横径(Fruit horizontal diameter,FHD,mm)、果实侧径(Fruit side diameter,FSD,mm)、果指数(FVD/FHD)、果皮厚(Pericarp width,PW,mm)、种子质量(Seed weight,SW,g)、种子纵径(Seed vertical diameter,SVD,mm)、种子横径(Seed horizontal diameter,SHD,mm)、种子侧径(Seed side diameter,SSD,mm)、种指数(SVD/SHD)、种仁鲜质量(Kernel fresh weight,KFW,g)、果皮皂苷含量(Saponin contents in the peep,SCP,%,质量分数)、种仁油脂含量(Oil contents in the kernel,OCK,%,质量分数)等共计15个性状,果皮含皂率和种仁含油率分别称总皂苷和总油脂。其中单株产量、种皮含皂率和种仁含油率的测定方法为:
(1)单株产量。在南北两面树冠的上、中、下3个位置,各选3个能代表结果平均状况的样枝,统计每个样枝的果实质量,计算3个样枝的平均质量,单株产量=结果枝数×样枝平均质量。果实成熟后,在树冠南面的中上部采样,分株采收、存放。果实自然风干后,采用GB/T 3543.7-1995的百粒四分法随机抽取30粒果实,用电子游标卡尺测量果实和种子纵径、横径、侧径和果皮厚,测量精度为0.01 mm,每粒种实的每性状测6次取平均值;计算果指数和种指数,果/种指数=纵径/横径;用1/1 000电子天平称量30粒果实的质量,称量精度为0.001 g;剥去果皮和种皮称种子和种仁的质量。
(2)果皮含皂率(总皂苷)。无患子果皮皂苷质量测定采用高效液相色谱分析法(HPLC)[16]。将30个果皮混合均匀后用微型粉碎机粉碎成粉末状,再放入70 ℃的电热恒温鼓风干燥箱中保持干燥。称取0.3 g果皮粉末,装入20 mL的带密封塞子的试管中,将甲醇用移液管定容至15 mL,立即盖上试管塞,浸泡20 h后,用漏斗过滤,并对滤液进行HPLC检测。HPLC检测以0.4 mg/mL常春藤皂苷元甲醇溶液为对照[14]。
色谱条件:色谱柱SymmetryTMC18 (3.9 mm×150 mm);流动相CH3CN∶H2O (H2O:90%~20%,30 min);色谱柱温度40 ℃;流速1 mL/min;检测波长210 nm。用下式计算果皮含皂率:
果皮含皂率=(纯皂苷质量/果皮干质量)×100%。
(3)种仁含油率(总油脂)。无患子种仁油脂含量采用索氏提取法[9]测定。将30个种仁混合均匀后用微型粉碎机粉碎,在80 ℃的电热鼓风干燥箱中烘干至恒质量,称取10 g种仁粉末,置于索氏提取器中,加正己烷200 mL,90 ℃回流提取9 h,至回流液无色(取回流液滴于滤纸上检查,待溶剂挥发尽,滤纸上不留油迹为提取终点)。用旋转蒸发仪回收正己烷,经干燥后即可得到黄色清亮的无患子籽油。按下式计算种仁含油率:
种仁含油率=(提取前种仁质量-提取后残渣质量) /提取前种仁质量×100%。
1.3 数据统计与分析
通过有效的采样、合理的统计分析,采用遗传上较稳定的性状,可以揭示群体的遗传规律和变异大小[17]。常用的研究方法主要包括聚类分析、相关性分析和主成分分析等,现已成功应用于多种作物资源的分类和育种研究中[18-19],对于植物资源利用和品种选育发挥了较好的指导作用。
本研究基础数据统计采用EXCEL软件。采用SPSS18.0软件对102份材料的15个性状进行类平均法系统分类、性状相关性分析和主成分分析,在进行聚类分析时需先对调查统计和测定数据进行标准化处理。
2 结果与分析
2.1 无患子实生群体种实表型性状的变异分析
常用变异系数(CV)比较量纲不同性状间的变异程度,变异系数越大则性状值离散程度越大[20]。从表1可以看出,无患子种实各性状指标在不同单株间变异程度不同,其变异系数由大到小依次为:单株产量(65.67%)>种仁鲜质量(27.54%)>总皂苷(23.96%)>果皮厚(23.14%)>果实质量(22.11%)>种子质量(16.20%)>果实横径(8.96%)>总油脂(8.18%)>果实侧径(7.89%)>种子横径(7.68%)>种子侧径(6.75%)>果实纵径(5.95%)>种子纵径(5.53%)>果指数(5.25%)>种指数(4.91%)。其中单株产量的变异系数最大,而果指数和种指数的变异系数较小,这说明无患子种实不同性状的离散程度不同,稳定性也不同。15个表型性状中,无患子单株产量在不同单株间变异最大,而果指数和种指数在不同单株间的变化相对稳定,但种指数的变异系数小于果指数,这说明种子形态较果实形态稳定。
2.2 无患子实生群体种实表型性状的相关分析
表2表明,无患子种实表型性状间的相关性比较复杂,大多存在显著和极显著相关性。其中单株产量与果实质量(P<0.01,r=0.832)、果实纵径(P<0.01,r=0.895)、果实横径(P<0.01,r=0.911)、果指数(P<0.01,r=0.820)、果皮厚(P<0.01,r=0.718)、种子质量(P<0.01,r=0.633)、种子纵径(P<0.01,r=0.794)和种子横径(P<0.01,r=0.600)均呈极显著正相关关系(P<0.01),与种指数呈显著正相关关系(P<0.05,r=0.218)。这说明单株产量受果实性状的影响较种子性状大;而在果实性状中,果实横径对单株产量影响最大。

表1 无患子实生群体种实表型性状的变异
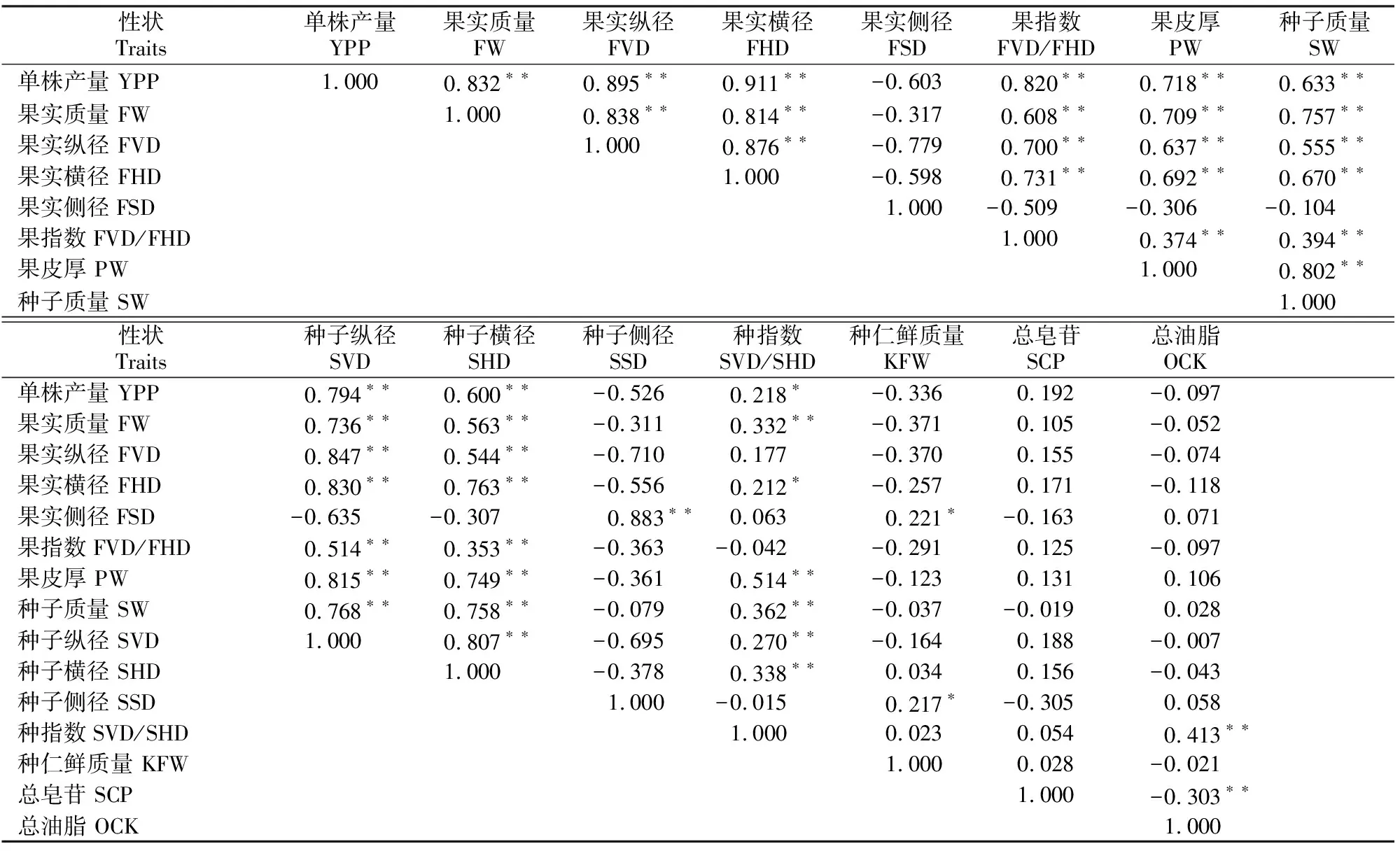
表2 无患子实生群体种实表型性状间的相关性
由表2可知,果实质量与果实纵径(P<0.01,r=0.838)、果实横径(P<0.01,r=0.814)、果指数(P<0.01,r=0.608)、果皮厚(P<0.01,r=0.709)、种子质量(P<0.01,r=0.757)、种子纵径(P<0.01,r=0.736)、种子横径(P<0.01,r=0.563)及种指数(P<0.01,r=0.332)均呈极显著正相关关系;种子质量与果实纵径(P<0.01,r=0.555)、果实横径(P<0.01,r=0.670)、果指数(P<0.01,r=0.394)、果皮厚(P<0.01,r=0.802)、种子纵径(P<0.01,r=0.768)、种子横径(P<0.01,r=0.758)及种指数(P<0.01,r=0.362)也均呈极显著正相关关系。这表明果实质量受果实形态性状影响较大,而种子质量受种子形态性状影响较大,其中纵径较横径对果实质量和种子质量的影响大。
在与果实质量或种子质量呈显著正相关的性状中,果指数和种指数对其影响均较小,但果指数较种指数对果实质量和种子质量的影响大。除种指数与总油脂呈极显著正相关关系(P<0.01,r=0.413)外,总皂苷和总油脂2种经济性状与果实质量性状及形态性状无显著的相关性,而总油脂与总皂苷间则呈极显著的负相关关系(P<0.01,r=-0.303),在本研究中,无患子群体的立地条件和栽培管理条件基本一致,则总皂苷和总油脂主要受遗传因素所影响。综上可知,无患子单株产量主要由果实性状决定,而总皂苷和总油脂则由遗传因素决定,这对筛选培育目标不同的无患子优良单株具有一定指导意义。
2.3 无患子单株组型的划分
由于无患子种实15个表型性状指标存在不同的量纲和数量级单位,为了使不同量纲、不同数量级的数据能放在一起进行比较,先将102棵无患子单株的15个数量性状数据进行标准化处理后,采用类平均法进行聚类分析,将不同性状特征的单株进行分组聚类。图1反映了无患子个体间的亲疏关系、表型变异层次和各组单株的特征,可见无患子性状分类在遗传距离为10.25时形成6个组,计算各组性状值的平均数用于各组特征分析,结果见表3。
图1表明,第1组共有29个单株。本组表型性状的基本特征是果实质量较大,组内果实的平均质量为5.90 g,其中88和99号的单株果实质量分别为7.42和7.98 g;该组果实体形较大,果实纵径和横径在各组中最大,分别达到20.94和25.05 mm,但本组果指数在6个组中最小,为0.84,说明本组果实形态更趋向于椭圆形;果皮厚和种仁鲜质量在各组中均最大,分别为2.52 mm和1.35 g;但单株产量和总皂苷居中后位置,总油脂仅次于第6组而居第2位。
第2组共有33个单株。本组的表型性状值在所有6组中大多较小,如组内果实质量为2.61~5.13 g,大部分低于4.30 g,仅45号单株单果质量超过5.00 g,且均值仅为4.04 g;单株产量11 kg,除显著低于第6组外,与其他组无显著差异。本组特征是果实小、产量和内含物含量较低,生产利用价值不大。
第3组共有35个单株。本组的表型性状值在6个组中大多位居第4位;组中单株的总皂苷大多在5.20%以上,其中68号单株最大,达10.27%,本组的均值为5.87%,居6个组之首;果皮厚和单株产量均较大,位居6个组的第2位;种仁鲜质量均在1.40 g以下,均值仅为0.93 g,处于6个组之尾。由于本组的总皂苷最大,且单株产量较高,在以皂苷为利用目的时参考价值较大。

图1 102株无患子单株表型性状的聚类分析
第4、5组均各有2个单株。第4组仅包括单株9和27,其特征是单株产量低、种子体形和质量大;另外,本组单株产量相差较大,2单株产量分别为15和3 kg;种子的表型性状在各分组中均较大,如质量、纵径、横径、侧径等;总皂苷居中,为5.78%;而总油脂在各分组中最低,仅为26.80%。第5组包括单株4和81。本组数量性状相对其余5组比较均匀,特征不太明显,与第4组一样生产利用价值不大。

表3 无患子单株聚类分组中6组种实表型性状均值的比较
利用类平均法聚类后,57号单株独自聚为一类,为第6组。本组特征是单果最圆最小、单株产量最大、总油脂最高(达39.80%),而总皂苷最低(仅为4.09%)。本组中,果实质量为3.21 g,约为第1组的54.4%;代表果实形态的纵径、横径和侧径在各组中均为最低;果形指数为0.92,在各组中最接近1,其形态是最近圆形的一组;而种指数为1.07,在各组中最远离1.00,说明本组无患子果实形态和种子形态无直接线性关系;本组单株产量为50 kg,约为其他各组的5倍。本组即第57号单株,在以油脂为利用目的的生产加工方面有较高的培育价值,可以作为无性繁殖材料利用。
2.4 无患子单株主要经济性状的主成分分析
根据102株无患子单株的种实表型性状,应用主成分分析法分析不同性状在无患子经济性状构成中的重要性,得到其特征值的贡献率和累计贡献率如表4所示,以累计方差贡献率达到80.000%以上确定主成分个数并建立主成分方程。

表4 无患子种实表型性状的主成分分析
由表4可以看出,在所有经济性状构成的主分量性状中,前4个主成分(PC-1、PC-2、PC-3、PC-4)的累计贡献率达到81.790%,且特征值大于1,说明4个主成分包含了所有性状的大部分信息,可以代替原来15个性状的信息。第1主成分(PC-1)单株产量、果实质量、果实纵径、果实横径、果皮厚、种子质量、种子纵径和种子横径等质量载荷较高,从经济性状出发,PC-1主要表征的是产量因子;同理,第2主成分(PC-2)主要表征的是化学成分因子,第3主成分(PC-3)和第4主成分(PC-4)主要表征了以种实指数和种实侧径为代表的种实形态因子。
PC-1和PC-2的贡献率分别是50.646%和 14.178%,说明PC-1和PC-2可以分别代表无患子产量和化学成分累积量。以PC-1和PC-2分别绘制不同表型性状主成分值散点图和单株主成分值样点图(图2、3)。图2和图3可以直观揭示无患子种实经济性状间的关系和单株经济性状间的差异状况,能更直观、简便地显示各单株经济性状的综合评价结果。在散点图上,横坐标与纵坐标值越大,表明无患子单株产量与总油脂和总皂苷的综合性状越好。

图2 无患子种实表型性状的第1和第2主成分值散点图
无患子种实表型性状的前2个主成分反映的是各单株经济性状的综合信息,与基于15个性状反映的结果基本一致,从而在性状选择的过程中达到了简化变量的目的。而群体的经济性状变异程度越大,则从中选择出优良单株的潜力就越大。在育种工作中,选配亲本材料应依据主成分的排序,具体分析与全面评价每个亲本材料综合指标的优劣,根据不同培育目标合理选配组合,以便尽快选育出无患子定向培育的新品种。
3 讨论与结论
表型性状受环境及栽培因素影响较大,在相同的生态环境下,植物不同性状的多样性能够较好地揭示其遗传本质,在实生繁殖条件下,种实各性状的遗传稳定性不一致或许是不同自然条件长期作用造成的[21]。本试验材料全部来自同一地点,可以消除生态环境差异引起的种实性状变异。已有的报道表明,不同产地无患子果皮皂苷含量差异显著,果皮皂苷含量与产地经度间存在显著性正相关关系[7],种实性状在不同地理分布区差异也很大[22]。
在本研究中,无患子15个种实表型性状的平均变异系数为4.91%~65.67%,大于树上干杏(“Shushanggan” apricot)的5.31%~26.22%[23]和毛梾(Conuswalter)的12.30%~16.61%[24],小于同科植物文冠果(XanthocerassorbifoliaBunge)人工种群种实的6.13%~78.36%[25]。本研究中,无患子变异系数最大和最小的性状分别是单株产量(65.67%)、种仁鲜质量(27.54%)、总皂苷(23.96%)和果指数(5.25%)、种指数(4.91%),而文冠果人工种群则为单株产量(78.36%)、单株种子产量(76.97%)、种子质量(30.05%)和种指数(8.66%)、种子横径(8.58%)、总油脂(6.13%)。这表明质量性状在2个树种的人工种群中均存在较大的变异,但无患子质量性状的变异幅度小于文冠果;在形态性状中,无患子和文冠果种实纵径的变异系数均大于种实侧径和横径,无患子果指数和种指数的变异系数(23.14%和27.54%)均大于文冠果的果指数和种指数 (13.25%和8.66%)[25]。种实指数是反映种实形态的一个重要指标,与纵、横径有密切关系,而纵、横径的变化具有一定的协同关系,这也保证了种实形态的基本稳定[26]。比较文冠果和无患子种实形态的变化,可见无患子种实形态存在较大的变异,其稳定性低于文冠果,无患子丰富的种实形态变异为优良品种的选育提供了可能。
本研究采用类平均法将102株无患子单株聚类为6个组群,每个组群形态或者内含物等都有自己的特征,这就从形态特征和经济性状角度初步明确了个体间的亲缘关系,为不同培育目标优良单株的选择奠定了基础[20]。对无患子种实15个表型性状进行主成分分析发现,前4个主成分累计方差贡献率为81.79%,基本可以综合反映15个性状指标的信息,因此在培育不同经济目标的优良单株时,可以通过计算主成分得分值进行快速准确的选择。通过对无患子种实表型性状相关性的分析发现,无患子种实性状间均存在比较密切复杂的关系,主要经济性状中单株产量、果实质量、种子质量与多数性状间呈显著或极显著相关关系,而总油脂和总皂苷与其他性状相关关系不太明显。因此,在相关性极强的性状间进行选择时,对其中一个性状进行改良,可以同时影响其他性状[18,23-24]。根据表2,改良果实质量和果实纵横径可以对单株产量产生较大影响。而对相关性不显著的性状进行选择时,一个性状的改良对其他性状影响不大,所以在性状选择上应当有不同的改良方向和选择标准[25-26]。如由于总皂苷与单株产量的相关性不明显,若要提高无患子单株皂苷产量,则需同时提高总皂苷和单株产量,因此需要从遗传改良和提高栽培管理措施等多方面入手。
表型性状相关性是植物资源研究和利用的重要内容。通过对性状间的相关性研究,确定不同性状之间的关系,可以在选择某一性状的同时预测其对其他性状可能产生的影响或选择压。表型性状的变异在适应性和进化上均具有重要意义[27]。植物种群中保持大的变异蓄积对种群是有利的,可使种群在整体上适应可能遇到的大多数环境条件[28]。本研究对无患子实生林15个种实表型性状的系统分析结果表明,该群体表型变异丰富,这些多层次的变异为不同利用目标的人工林培育提供了选择基础。依据表型分类形成的组群均有明显的形态独特性或独特基因型,加之数量性状一般呈连续性分布,每一类群内均有某一个或多个性状的极端个体,这在育种过程中具有较大的实际应用价值[27]。根据不同育种目标,利用不同类型的优良基因资源进行基因重组和轮回选择,对于种质改良、创新以及杂交种的组配均具有重要的指导意义。
[参考文献]
[1] Allard R W.The genetics of inbreeding populations [J].Advances in Genetics,1968,14:55-131.
[2] Price S C,Shumaker K M,Kahler A L,et al.Estimates of population differentiation obtained from enzyme polymorphisms and quantitative characters [J].Journal of Heredity,1984,75(2):141-142.
[3] 曾 杰,白嘉雨.植物天然居群表型变异研究的几个热点问题 [J].广西林业科学,2007,36(2):65-70.
Zeng J,Bai J Y.Some hot issues on phenotypic variation of natural plant populations [J].Guangxi Forestry Science,2007,36(2):65-70.(in Chinese)
[4] Wheeler N C,Guries R P.Population structure,genetic diversity,and morphological variation inPinuscontortaDougl [J].Canadian Journal of Forest Research,1982,12:595-606.
[5] 《中国植物志》编辑委员会.中国植物志:47卷第1分册 [M].北京:科学出版社,1998:14-15.
Flora of China Editorial.Flora republicae popularis sinicae:Vol.47-1 [M].Beijing:Science Press,1998:14-15.(in Chinese)
[6] Chhetri A B,Tango M S,Budge S M,et al.Non-edible plant oils as new sources for biodiesel production [J].International Journal of Molecular Sciences,2008,9(2):169-180.
[7] 邵文豪,姜景民,董汝湘,等.不同产地无患子果皮皂苷含量的地理变异研究 [J].植物研究,2012,32(5):627-631.
Shao W H,Jiang J M,Dong R X,et al.Geographic variation of Saponins contents inSapindusmukorossipeels from different habitats [J].Bulletin of Botanical Research,2012,32(5):627-631.(in Chinese)
[8] Ghagi R,Satpute S K,Chopade B A,et al.Study of functional properties ofSapindusmukorossias a potential biosurfactant [J].Indian Journal of Science and Technology,2011,4(5):530-533.
[9] 黄素梅,王敬文,杜孟浩,等.无患子籽油脂肪酸成分分析 [J].中国油脂,2009,34(12):74-76.
Huang S M,Wang J W,Du M H,et al.Fatty acid composition analysis ofSapindusmukorossiGaerth. seed oil [J].China Oils and Fats,2009,34(12):74-76.(in Chinese)
[10] Sun S D,Ke X Q,Cui L L,et al.Enzymatic epoxidation ofSapindusmukorossiseed oil by perstearic acid optimized using response surface methodology [J].Industrial Crops and Products,2011,33(3):676-682.
[11] 国家林业局.国家林业局关于印发灌木能源林培育利用指南和无患子原料林可持续培育指南的通知[EB/OL].(2012-05-12) [2013-09-24].http://www.forestry.gov.cn/portal/main/govfile/13/govfile_1935.htm.
State Forestry Administration of China.The notice about printing and distributing cultivate guide of shrub forests of energy andSapindusmukorossiraw material forests continued to be used from the State Forestry Administration of China [EB/OL].(2012-05-12) [2013-09-24].http://www.forestry.gov.cn/portal/main/govfile/13/govfile_1935.htm.(in Chinese)
[12] 黄 顺.优良造景树:无患子 [J].园林,2004(9):42.
Huang S.Good landscape tree:Sapindusmukorossi[J].Garden,2004(9):42.(in Chinese)
[13] 张凤龙.无患子组织培养研究 [J].山地农业生物学报,2005(2):119-123.
Zhang F L.Study on tissue culture for soapberry [J].Journal of Mountain Agriculture and Biology,2005(2):119-123.(in Chinese)
[14] 魏凤玉,方 春.酶法提取无患子皂苷的工艺研究 [J].应用化工,2010,39(8):1149-1151.
Wei F Y,Fang C.Enzyme-assisted aqueous extraction ofSapindussaponins[J].Applied Chemical Industry,2010,39(8):1149-1151.(in Chinese)
[15] Kanchanapoom T,Kasai R,Yamasaki K.Acetylated triterpene saponins from the Thai medicinal plant,Sapindusemarginatus[J].Chemical and Pharmaceutical Bulletin,2001,49(9):1195-1197.
[16] Qi L W,Yu Q T,Li P,et al.Quality evaluation ofRadixastragalithrough a simultaneous determination of six major active isoflavonoids and four main saponins by high-performance liquid chromatography coupled with diode array and evaporative light scattering detectors [J].Journal of Chromatography A,2006,1134(1):162-169.
[17] 肖海峻,徐 柱,翟利剑,等.鹅观草居群表型多样性研究 [J].西北植物学报,2007,27(11):2222-2227.
Xiao H J,Xu Z,Zhai L J,et al.Phenotypic diversity ofRoegneriakamojiOhwi population [J].Acta Bot Boreal-Occident Sin,2007,27(11):2222-2227.(in Chinese)
[18] 郭宝林,杨俊霞,李永慈,等.主成分分析法在仁用杏品种主要经济性状选种上的应用 [J].林业科学,2000,36(6):53-56.
Guo B L,Yang J X,Li Y C,et al.The application of principal component analysis on mainly economic characters and superior variety selection of apricot for nucleolus using [J].Scientia Silvae Sinicae,2000,36(6):53-56.(in Chinese)
[19] 郭军战,张 敏,费昭雪,等.文冠果数量性状的主成分分析及聚类分析研究 [J].西北林学院学报,2012,27(2):66-69.
Guo J Z,Zhang M,Fei Z X,et al.Principal component analysis and cluster analysis of quantitative characters ofXanthocerassorbifolia[J].Journal of Northwest Forestry University,2012,27(2):66-69.(in Chinese)
[20] 苗锦山,刘彩霞,戴振建,等.葱种质资源数量性状的聚类分析、相关性和主成分分析 [J].中国农业大学学报,2010,15(3):41-49.
Miao J S,Liu C X,Dai Z J,et al.Clustering correlation and principal component analyses in welsh onion (AlliumfistulosumL.) germplasm resources [J].Journal of China Agricultural University,2010,15(3):41-49.(in Chinese)
[21] Greipsson S,Davy A J.Seed mass and germination behaviour in populations of the dune-building grassLeymusarenarius[J].Annals of Botany,1995,76(5):493-501.
[22] 辜夕容.不同种源无患子的种子品质差异分析 [J].西南大学学报:自然科学版,2009,31(6):51-54.
Gu X R.Analysis of the differences in seed characters ofSapindusmukorossiGaertn with five different provenances [J].Journal of Southwest University:Natural Science Edition,2009,31(6):51-54.(in Chinese)
[23] 田治国,王 飞,王 朴,等.新疆树上干杏形态学形状的多样性研究 [J].西北农林科技大学学报:自然科学版,2012,40(3):147-153.
Tian Z G,Wang F,Wang P,et al.Morphological diversity of ‘shushanggan’ apricot in Xinjiang [J].Journal of Northwest A&F University:Nat Sci Ed,2012,40(3):147-153. (in Chinese)
[24] 康永祥,赵宝鑫,贠玉洁,等.毛梾天然群体种实表型多样性研究 [J].西北农林科技大学学报:自然科学版,2011,39(9):107-117.
Kang Y X,Zhan B X,Yun Y J,et al.Study on phenotypic diversity of seeds and fruits’ characteristics inCornuswalteri[J].Journal of Northwest A&F University:Nat Sci Ed,2011,39(9):107-117.(in Chinese)
[25] 柴春山,芦 娟,蔡国军,等.文冠果人工种群的果实表型多样性及其变异 [J].林业科学研究,2013,23(2):181-191.
Chai C S,Lu J,Cai G J,et al.Fruit phenotypic diversity and variation ofXanthocerassorbifoliaartificial population [J].Forest Research,2013,23(2):181-191.(in Chinese)
[26] 周 旋,何正飚,康宏樟,等.温带-亚热带栓皮栎种子形态的变异及其与环境因子的关系 [J].植物生态学报,2013,37(6):481-491.
Zhou X,He Z B,Kang H Z,et al.Variations of seed morphology related to climate forQuercusvariabilisacross temperate-subtropical China [J].Chinese Journal of Plant Ecology,2013,37(6):481-491.(in Chinese)
[27] Baeilieri R,Ducousso A,Kremer A.Genetic morphological ecological and phonological differentiation betweenQuercuspetraea(Matt.) Label. andQuercusroburL. in a mixed stand of Northwest of France [J].Silvae Genetica,1995,44(1):1-10.
[28] 杨生超,徐绍忠,闻国松,等.灯盏花种质资源群体表型多样性研究 [J].西北植物学报,2008,28(8):1573-1579.
Yang S C,Xu S Z,Wen G S,et al.Phenotypic diversity of populations in germplasm resources ofErigeronbreviscapus[J].Acta Bot Boreal-Occident Sin,2008,28(8):1573-1579.(in Chinese)
