广西金钟山国家级自然保护区景观破碎化分析
2014-03-22蒋爱伍李秀明原宝东覃国乐
蒋爱伍,李秀明,原宝东,覃国乐
(1.广西大学林学院,广西南宁530004;2.中国林业科学研究院森林生态环境与保护研究所,北京100091;3.河池学院化学与生物工程学院,广西宜州546300)
0 引言
景观格局是各种生态过程在不同尺度上作用的结果[1],是理解生态空间变化的一种主要景观生态学工具[2-3]。景观斑块的类型、形状、大小、数量和空间组合影响着该区域的生态过程和边缘效应[4]。景观格局分析可以借助具体景观格局指数来描述抽象的空间格局,从一些看似无序的景观要素中得出景观格局产生的机制[5]。人为干扰对自然景观格局影响较大,进而导致的景观破碎化或动物灭绝的机制成为保护生物学的热点问题[6]。在自然干扰或者人为活动的影响下,大面积连续分布的生境会被分割成小面积不连续的生境斑块,而地栖性的雉科鸟类迁移和扩散能力弱,因此生境破碎化对雉类的影响尤其严重[7]。
金钟山自然保护区位于广西壮族自治区西北部的隆林和西林县境内,是我国唯一以国家Ⅰ级重点保护动物黑颈长尾雉(Syrmaticus humiae)为主要保护对象的国家级保护区。黑颈长尾雉在我国仅分布于云南、广西西北部和贵州东南部[8-10]。在国外见于泰国西北部、印度阿萨姆邦东北部和缅甸。黑颈长尾雉被IUCN列为全球性近危物种,其种群数量被估计在10 000~19 999只之间[11]。金钟山保护区范围内及周边林区均有黑颈长尾雉分布,保存有我国数量最大的野生种群。本文对广西金钟山国家级自然保护区的景观格局进行了调查分析,通过对金钟山保护区基底和景观破碎化程度的调查,为片断化生境里的黑颈长尾雉保护提供科学依据。
1 研究地区及方法
1.1 研究地区
本文选取广西金钟山国家级自然保护区、金钟山林场以及周边有黑颈长尾雉活动的集体林地进行调查,地理坐标为104°46'19″~104°59'56″E,24°32'42″~24°45'07″N,海拔为670~1 818 m。当地属中亚热带季风气候,年平均气温19.1℃,年均降雨量1 170 mm左右,5~9月为雨季,10月至翌年4月为旱季。植物区系成分复杂,为泛北极植物区向古热带植物区交汇处。海拔1200 m以上为山地常绿阔叶林;海拔850~1 250 m为以细叶云南松(Pinus yunnanensis)占优势或与栓皮栎(Quercus variabilis)、麻栎(Quercus acutissima)、高山栲(Castanopsis delavayi)等组成针阔混交林;海拔850 m下为细叶云南松林。
1.2 研究方法
根据2009年至2012年对研究区域的野外调查和2009年森林资源二类调查资料,使用ArcGIS 9.3软件对拍摄于2009年LANDSAT卫星遥感图进行矢量化,研究区域的斑块可分为11种景观类型(见表1)。选取16个景观指数,通过斑块平均面积、总周长、最大斑块所占景观面积的比例、整个景观的斑块密度和斑块个数了解各景观对整体景观的破碎化影响程度;通过景观组分斑块密度、周长面积分维、连接度、景观形状指数了解各景观的破碎化程度,运用香农多样性指数、Simpson多样性指数、香农均匀度指数、蔓延度值分析景观多样性。运用景观结构分析软件Fragstats 3.3(栅格版)计算这些相关景观指数,Cellsize选择5 m×5 m,Cell assignment type选择CELL_CENTER。运用ArcGIS 9.3软件绘制景观格局分布图,用Excel 2007整理Fragstats 3.3运算的结果。
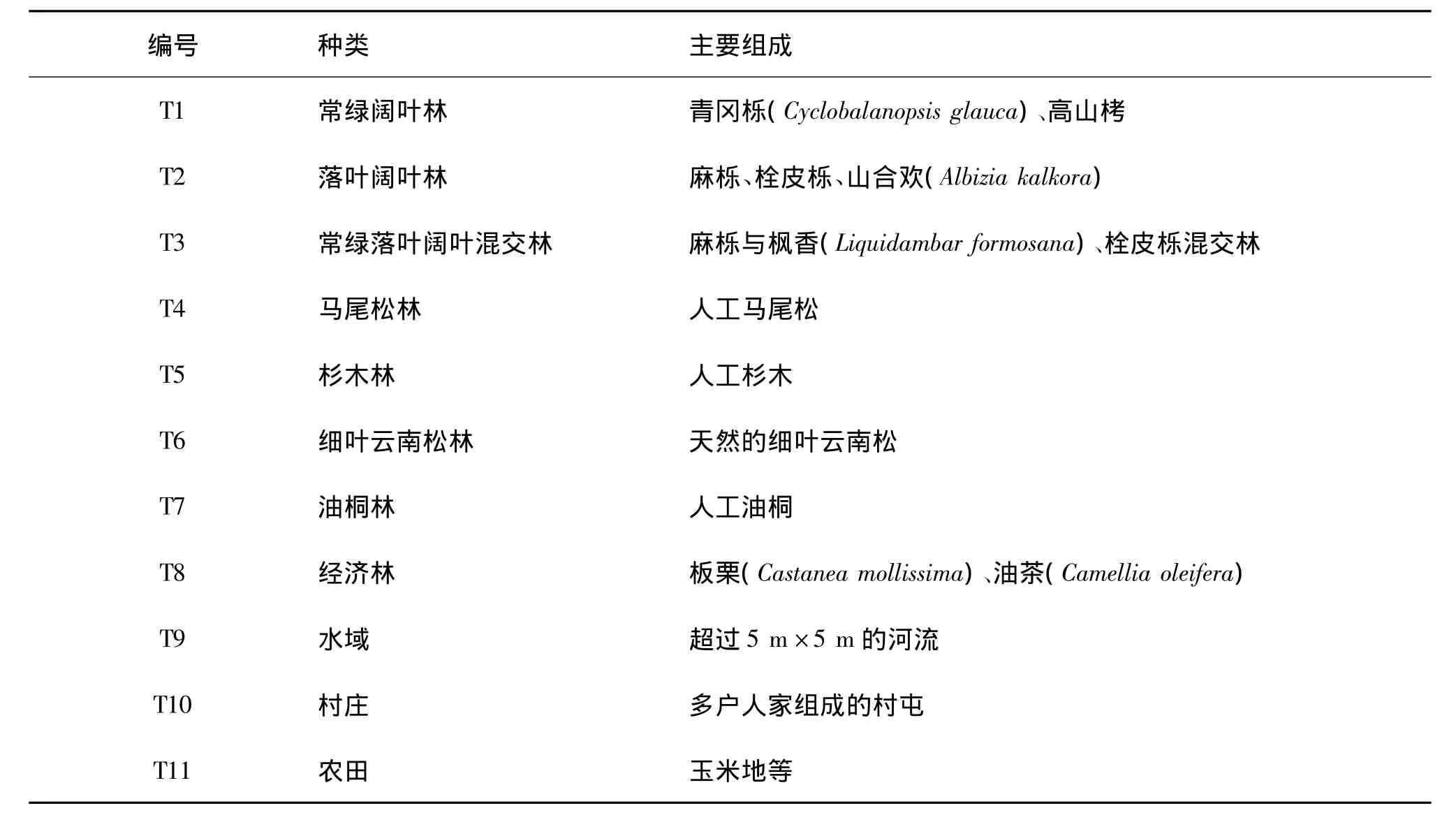
表1 广西金钟山保护区景观类型
2 结果与分析
研究地面积共计38 716.6 hm2,共有斑块1 708个,景观总周长526.5 km。其中黑颈长尾雉可栖息面积为29 552.3 hm2,占总面积的76.3%;斑块个数为986个,占总斑块数目的57.7%(见表2)。金钟山保护区以阔叶林(包括常绿阔叶林、落叶阔叶林、常绿落叶阔叶混交林)为基质,因此天然植被在控制景观整体结构、功能和动态过程中起着主要作用(见图1)。水域呈带状分布,主要分布金钟山保护区的边缘,对景观破碎化影响较小。农田、经济林、油桐和杉木林对基质的破碎化程度起到主要作用,这些人工景观对自然景观内动植物起到干扰作用。油桐林和杉木林受人为干扰严重,边缘较为规则,破碎化程度高。
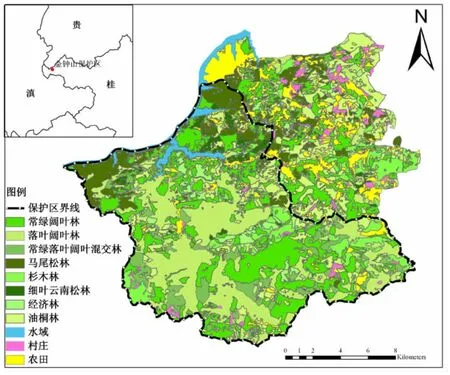
图1 广西金钟山自然保护区景观格局分布
2.1 景观组分面积特征
景观类型所占整个景观面积的比例表示这一景观在相对意义上对整个景观的贡献率。金钟山保护区以常绿阔叶林面积占总面积比例最大,村庄所占比例最小(见表2)。落叶阔叶林、常绿阔叶林及常绿落叶阔叶混交林面积占整个景观的55.3%。马尾松林、杉木林、农田和村庄的平均面积均小于整个景观斑块平均面积,表明它们的破碎化程度大。细叶云南松林所占面积比例较小,但它的平均斑块面积却接近常绿阔叶林与油桐林,表明其破碎化程度相对较低。水域在研究地边缘,在整个景观中所占比例较小,对其他景观影响小。
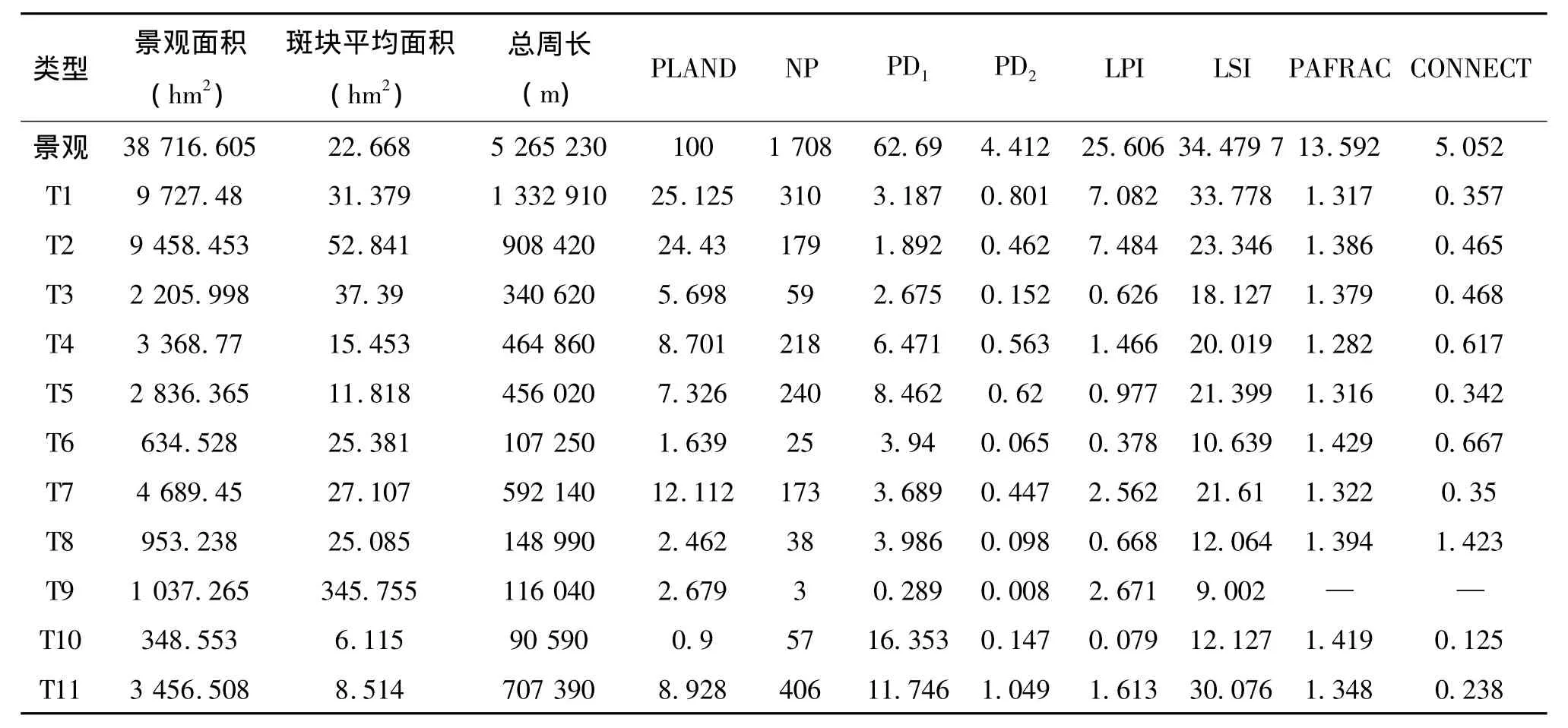
表2 广西金钟山保护区的景观特征
2.2 景观组分周长
各景观组分周长大小顺序为:T1>T2>T11>T7>T4>T5>T3>T8>T9>T6>T10,其总趋势与景观组分面积趋势略有不同。常绿阔叶林在各景观类型中拥有最大周长,达1 332.9 km。各景观类型周长与景观总周长的比例,对于一些分布在边界的物种研究很有意义。金钟山保护区内比例最高的为常绿阔叶林25.3%,其次是落叶阔叶林,其余均不超过15.0%。表明常绿阔叶林和落叶阔叶林能容纳较多边界物种。
2.3 景观组分的斑块数量
金钟山保护区内各景观组分的斑块数量极不均衡。其中农田拥有406个斑块,数量最多,占总斑块数的23.8%;其次是常绿阔叶林、杉木林、马尾松林、落叶阔叶林、油桐林;其余类型的斑块数量均小于10%。斑块平均面积最大的水域仅有3个斑块。农田斑块数远多于常绿阔叶林和落叶阔叶林,但其总面积远小于两者。农田的面积同马尾松林面积占总面积比例相近,斑块平均面积相差不大,但其斑块数也远高于马尾松林斑块数,表明农田斑块数量多而面积小,破碎化程度高。
各个景观组分的斑块密度PD1(景观组分的斑块数/景观组分面积)直接地反映了景观组分的破碎化程度。斑块密度最高的是村庄,远大于整个景观的平均斑块密度(总斑块数/总面积),对其他景观有较大影响,但是其面积占总面积比例(PLAND)仅为0.9%,不能对整个景观破碎化起主导作用。油桐林斑块密度与经济林相近,表明其破碎化程度与经济林相近,但其面积远大于经济林,对整个景观影响最大。
类型斑块数与景观总面积之比表示景观基质被该类型斑块分割的程度,即这一景观组分在整个景观上的斑块密度PD2(亦称孔隙度),这一指标对生物保护、物质和能量分布具有重要影响。各景观组分斑块密度大小顺序为T11>T1>T5>T4>T2>T7>T3>T10>T8>T6>T9。斑块密度最大的农田为1.0个/km2,即整个景观基质阔叶林被农田分割程度较大。面积较大的落叶阔叶林、常绿阔叶林、常绿落叶阔叶混交林面积之和在整个景观面积中比例超过50.0%,而常绿阔叶林斑块密度大于其他阔叶林,说明常绿阔叶林在阔叶林中起主导地位。油桐林与落叶阔叶林斑块密度相近,但油桐林的面积却是落叶阔叶林的一半,可见油桐林在整个景观中影响超过了落叶阔叶林。
2.4 景观组分分维与形状指数
分维数(PAFRAC)可直观地理解为不规则几何形状的非整数维数,这一指标用于揭示各景观组分的边界褶皱程度。金钟山保护区各景观组分维数差别不大,说明斑块边界褶皱程度差别不大。农田的斑块数<10,没有实际意义,故不计算。最大分维数为细叶云南松林,马尾松林分维数最小。表明细叶云南松林斑块边界褶皱程度最为曲折复杂,而杉木林、油桐林、马尾松林等人工林相对分维指数相对低,是因为人工种植时总会将其开垦得比较整齐。整个景观水平上的周长面积分维数为1.34(见表3),表明整个景观边界的褶皱程度低。

表3 广西金钟山保护区景观指标
景观形状指数反映景观组分斑块的复杂程度。形状指数越大,斑块形状就越不规则,曲折程度就越高,反之,斑块形状越规则越平滑。作为建群种的细叶云南松林斑块平均形状指数较高,说明其形状复杂。而整个森林景观形状指数中常绿落叶阔叶林最大。经济林与常绿阔叶林斑块平均形状指数相近,但经济林面积远小于常绿阔叶林,表明常绿阔叶林景观较为复杂,生态系统更为稳定。
2.5 景观多样性分析
多样性指数是景观中各类斑块的复杂性和变异性的量度,通常随着多样性指数的增加,景观结构组成成分的复杂性也趋于增加。金钟山保护区的景观多样性指数。从景观内部看其香农均匀度指数为0.85,说明各景观组分面积比较接近。蔓延度值为55.34,说明各景观类型所占比例差别大,景观成分分配不均匀,特别是3类阔叶林与农田、村庄之间较大的差别,导致了均匀指数比较大。香农多样性指数为2.03,表明研究区景观破碎化已经较为严重。
3 讨论
景观格局分析是景观生态学中用于分析景观过程的重要方法,而景观指数是满足景观格局分析的重要指标[12]。景观指数表明金钟山保护区景观破碎化严重,特别是保护区外的林场及集体林地。在整个景观尺度上,阔叶林占50%以上,而主要影响景观破碎化的农田和村庄所占比例小于10%,并不影响阔叶林的优势地位,但是金钟山的村庄斑块密度最大,农田的斑块数量最多,因此人为干扰可能是造成栖息地景观破碎化的主要原因。黑颈长尾雉适宜的栖息地连接度都不高,表明已经遭到一定程度的破坏。在运用景观格局指数进行分析时,多数的研究只考虑到空间尺度,忽略了时间尺度[13],这使得在分析景观过程中所表现出来的特征不能代表景观的本质规律,这也是没有单独通过景观指数分析景观过程的原因之一。
在景观尺度上,斑块的面积、形状和周长影响着物种多样性、生物迁移、内部种和边缘种的扩散等[14-15]。黑颈长尾雉在金钟山主要栖息地类型为细叶云南松林及针阔混交林、常绿落叶阔叶混交林,但也可在油桐林和人工杉木林活动[16-17]。金钟山保护区有大片适宜其栖息的破碎化程度较低的阔叶林和连通性较好的细叶云南松林斑块,但黑颈长尾雉也会选择人为干扰严重、破碎化程度高的的杉木林和油桐林,可能是因为油桐林和杉木林中有适宜的水源、隐蔽性好、易于躲避天敌和丰富的食物等优良条件,同为针叶林但连接度较大的马尾松林内并没有发现黑颈长尾雉的生存,可能是马尾松林受到的人为干扰特别严重,已经超过了黑颈长尾雉的耐受范围。
动物的生活史研究能为预测和理解整个景观变化提供理论支持[18],斑块资源的季节性变化反映了动物在不同生活史阶段对资源需求的差异,在同一斑块往往不能满足整个生活史的需求[19]。在繁殖季节,有些鸟类甚至通过迁徙寻找合适的生境。野外调查发现繁殖季节黑颈长尾雉会常常在不同斑块间移动,有时候甚至离开长期栖息的斑块1个月之久,斑块资源的不均匀性和单一性迫使黑颈长尾雉花费更多的能量寻找合适的资源。
生境丧失和生境退化以及偷猎是黑颈长尾雉最主要的威胁[10,17],此次研究表明金钟山地区仍广泛生存着黑颈长尾雉,只是各斑块种群难以交流,形成了一定程度的栖息地片段化。最近在贵州省发现的黑颈长尾雉新分布点,其栖息地同样受到破碎化的威胁[9],此外,火灾和气候变化也威胁着在泰国的黑颈长尾雉的生存[20]。随着人口的增多,人为干扰强度的加大,金钟山地区黑颈长尾雉栖息地在不断缩小,被分割成破碎化的生境斑块,使得黑颈长尾雉各个种群栖息地斑块之间仍面临着被孤立的危险,有些种群甚至面临着生境丧失的威胁。同时,金钟山保护区外的金钟山林场及集体林也发现有黑颈长尾雉存在,这些地区传统的农耕火种观念严重,毁林开荒和偷捕滥猎现象屡禁不止。因此,当务之急是评估黑颈长尾雉种群生存现状,控制金钟山地区已经出现的破碎化景观,并通过生境恢复、廊道重建及增强群众保护意识等措施加强对黑颈长尾雉的保护。
[1] 傅伯杰,陈利顶,马克明.景观生态学原理及应用[M].北京:科学出版社,2001:1-396.
[2] Fu B,Chen L.Agricultural landscape spatial pattern analysis in the semi-arid hill area of the Loess Plateau,China[J].Journal of Arid Environ-ments,2000,44(3):291-303.
[3] 傅伯杰.黄土区农业景观空间格局分析[J].生态学报,1995,15(2):113-120.
[4] 王亚明,薛亚东,夏友福.滇西北滇金丝猴栖息地景观格局分析及其破碎化评价[J].林业调查规划,2011,36(2):34-37.
[5] 肖笃宁,李秀珍,高峻,等.景观生态学[M].北京:科学出版社,2003:1-351.
[6] 杨勇,温俊宝,胡德夫.鸟类栖息地研究进展[J].林业科学,2011,47(11):172-180.
[7] 丁平,姜仕仁,诸葛阳.浙江西部白颈长尾雉栖息地片断化研究[J].动物学杂志,2000,21(1):65-69.
[8] 李飏,于晓平.云南开远市鸟类多样性[J].四川动物,2011,30(3):415-420.
[9] Jiang A,Cheng Z,Liang X.Discovery of Hume’s Pheasant(Syrmaticus humiae)in Guizhou Province,southwestern China[J].Chinese Birds,2012,3(2):143–146.
[10] 韩联宪.云南黑颈长尾雉(Syrmaticus humiae)分布及栖息地类型调查[J].生物多样性,1997,5(3):185-189.
[11] Groombridge B.IUCN Red list of Threatened Animals[M].Gland:IUCN,1994:42.
[12] 陈文波,肖笃宁,李秀珍.景观空间分析的特征和主要内容[J].生态学报,2002,22(7):1 080-1 087.
[13] Gustafsson EJ.Quantifying landscape spatial pattern:what is the state of the art?[J].Ecosystems,1998,1(2):143-156.
[14] Baker WL,Cai ZM.The role programs for multi-scale analysis of landscape structure using the GRASS geographical information system[J].Landscape Ecology,1992,7(4):291-302.
[15] 张明.榆林地区脆弱生态环境的景观格局与演化研究[J].地理研究,2000,19(1):30-36.
[16] 蒋爱伍,周放,陆舟,等.广西黑颈长尾雉对夜宿地的选择[J].动物学研究,2006,27(3):249-254.
[17] 刘小华,周放,潘国平,等.黑颈长尾雉繁殖习性的初步研究[J].动物学报,1991,37(3):332-333.
[18] Hansen AJ,Urban DL.Avian response to landscape pattern:The role of species’life histories[J].Landscape Ecology,1992,7(3):163-180.
[19] Zhou W,Li N,Deng Z,et al.Modeling foraging habitats of Hume’s Pheasant(Syrmaticus humiae)in Dazhong Mountain,Yunnan,southwestern China[J].Chinese Birds,2010,1(4):236-243.
[20] Iamsiri A,Gale G.Breeding Season Habitat Use by Hume’s Pheasant Syrmaticus humiae in the Doi Chiang Dao Wildlife Sanctuary,Northern Thailand[J].Zoological Studies,2008,47(2):138-145.
