重要籼型杂交稻亲本“93-11”稻米食味品质的改良
2014-03-20窦兰兰李建粤
窦兰兰,肖 璐,李建粤,2
(1.上海师范大学 生命与环境科学学院,上海 200234; 2.上海师范大学 植物种质资源开发中心, 上海 200234)
世界上有一半人口以稻米为主粮.近年来随着生活水平的日益提高,人们对水稻稻米的品质要求也越来越高[1].影响稻米食味品质最主要因素是直链淀粉含量(AC)[2].有研究报道认为,稻米直链淀粉含量还与影响稻米柔软程度的胶稠度指标也密切相关[3-4].
稻米直链淀粉合成主要受蜡质基因编码的淀粉合成酶控制[5-6].目前已有较多报道显示,导入水稻中的反义蜡质基因能够降低稻米直链淀粉含量[7-12],以此达到改良水稻食味品质的目的.籼型水稻“93-11”由江苏省里下河地区农科所培育而成.该品种目前在水稻生产中主要被用作两系杂交稻的父本.在生产上大面积推广应用的两系杂交稻“两优培九”、“广两优6号”等品种都是以“93-11”水稻为父本成功培育的.因此,利用转基因技术导入反义蜡质基因降低“93-11”稻米直链淀粉含量,由此改善“93-11”稻米的食味品质研究将具有良好的应用前景.然而目前的研究显示,籼稻为受体进行根癌农杆菌介导遗传转化,成功率相对较低[13].本实验室曾构建含有双份反义蜡质基因的表达载体:p13AWY-2[14],并成功转化粳型两系不育系水稻“261S”[15].
本研究以籼稻“93-11”和含有双份反义蜡质基因的纯合转基因水稻株系“B3”为供试材料,利用杂交及多次与“93-11”水稻进行回交,并采用常规及分子标记辅助,选育出各种农艺性状与“93-11”水稻接近,同时含有双份反义蜡质基因的改良型“93-11”水稻.开展本研究可为改良以“93-11”水稻为父本的杂交稻食味品质奠定了基础.
1 材料与方法
1.1 水稻亲本材料
本试验用亲本水稻材料分别为籼稻“93-11”以及将只含有双份反义蜡质基因、没有抗性基因和报告基因的质粒载体(图1)转化粳型两系不育系“261S”获得的纯合转基因株系“B3”水稻[15].
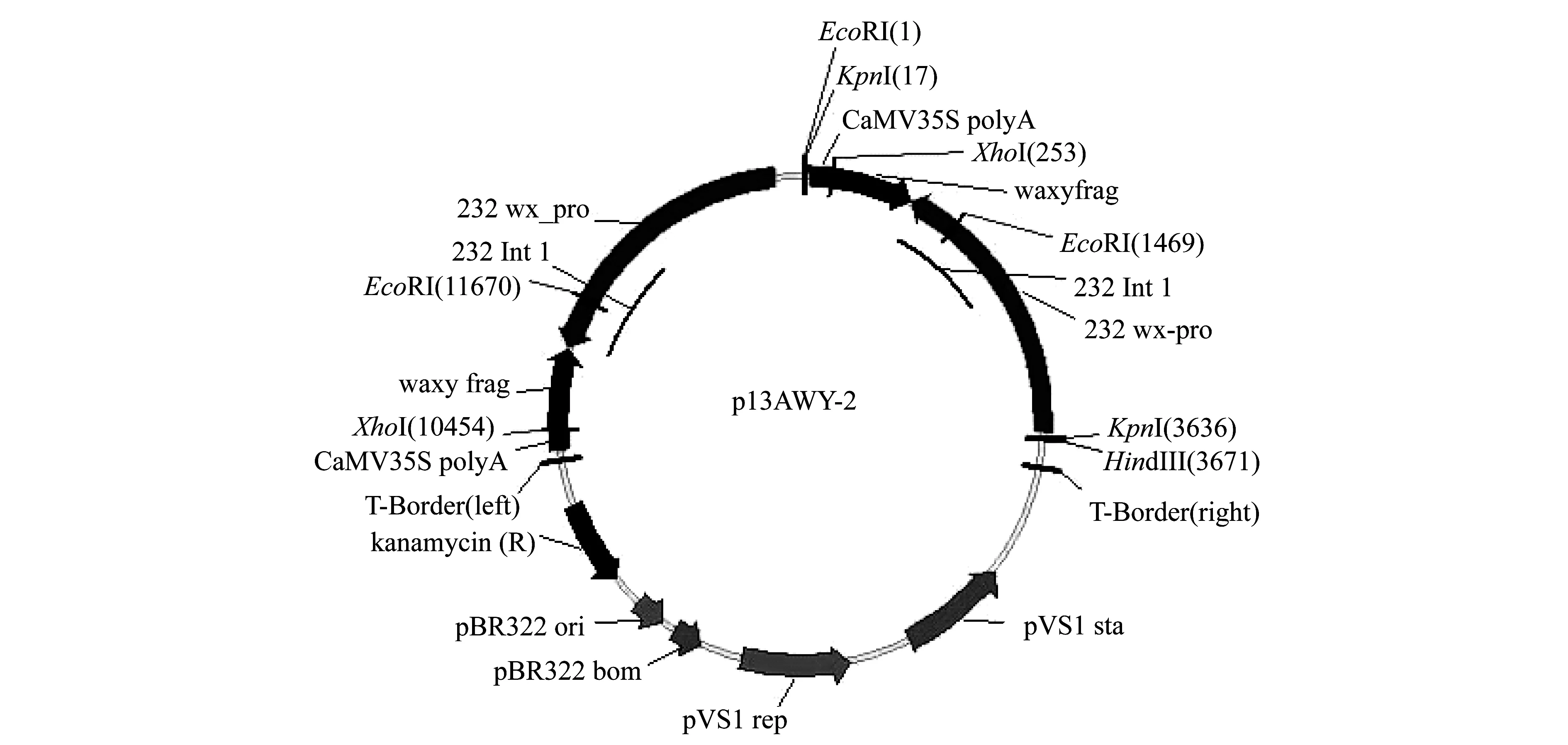
图1 含有两份双份反义蜡质基因质粒(p13AWY-2)载体结构
1.2 水稻植株DNA提取及反义蜡质基因的检测
取水稻叶片,采用CTAB法提取叶片总DNA.参照杨瑞报道[15]合成检测反义蜡质基因的引物.上、下游引物序列分别为:5′-CTCTCGGAGAGGTTCAGCTC-3′(YW5)和5′-CTTAATAACACATTGCGGACG-3′(YWf).PCR扩增反应程序为:94 ℃预变性5 min,94 ℃变性40 s,退火温度55 ℃,30 s、72 ℃延伸45 s,共30个循环,最后72 ℃延伸10 min.采用1%琼脂糖凝胶电泳对PCR产物进行检测.
1.3 改良后“93-11”水稻与对照农艺性状和拷种分析以及糙米性状考察
测量植株株高,记录每株有效穗数,考察每穗总粒数、空粒数和实粒数,并计算结实率,称量稻谷的千粒重和糙米重量;观察含有反义蜡质基因植株糙米外观;参考朱映东等报道[16],测量糙米体积.
1.4 改良后“93-11”水稻与对照糙米部分理化指标分析
取成熟种子的糙米,按照国标GB/T 15682—2008方法测定直链淀粉含量,按照国标GB/T 22294—2008检测糙米胶稠度[17].
2 结果与分析
2.1 改良后“93-11”水稻培育过程及反义蜡质基因的检测
以“B3”水稻为母本,“93-11”水稻为父本,进行杂交获得F1代种子.以F1代植株为母本与“93-11”水稻回交,获得BC1F1种子.在BC1F1植株种植期间,提取BC1F1植株叶片DNA,采用PCR检测反义蜡质基因.将含有反义蜡质基因的BC1F1植株为父本,“93-11”水稻为母本又进行回交,获得BC2F1种子.以同样的方式将BC2F1植株为父本再与 “93-11”水稻回交2次,先后获得BC3F1和BC4F1种子.在每次回交前也同样采用PCR分析植株的反义蜡质基因,而且选取株型与“93-11”水稻尽量相似的植株进行目的基因的检测和回交.在收获BC4F1种子的同时,从含有反义蜡质基因的BC3F1植株上收获自交BC3F2种子.继续种植BC4F1种子和BC3F2种子,两种对应植株小区种植编号分别为2011-59和2011-60.同样取编号为2011-59和2011-60小区植株叶片,进行反义蜡质基因检测,并以单株形式收获两类含有目的基因的2011-59(BC4F2)和2011-60(BC3F3)种子.
在采用PCR检测回交植株的反义蜡质基因时,均选用原先转化获得“B3”水稻质粒(p13AWY-2)DNA为阳性对照,“93-11”水稻DNA为阴性对照.图2分别显示2011-59(泳道18~32)和2011-60(泳道3~17)两种编号部分植株基因组DNA扩增反义蜡质基因的检测结果.反义蜡质基因的PCR产物为650 bp,含有与p13AWY-2质粒相同分子量条带的植株,表明其含有反义蜡质基因,而在“93-11”水稻中没有相应的目的条带(图2).
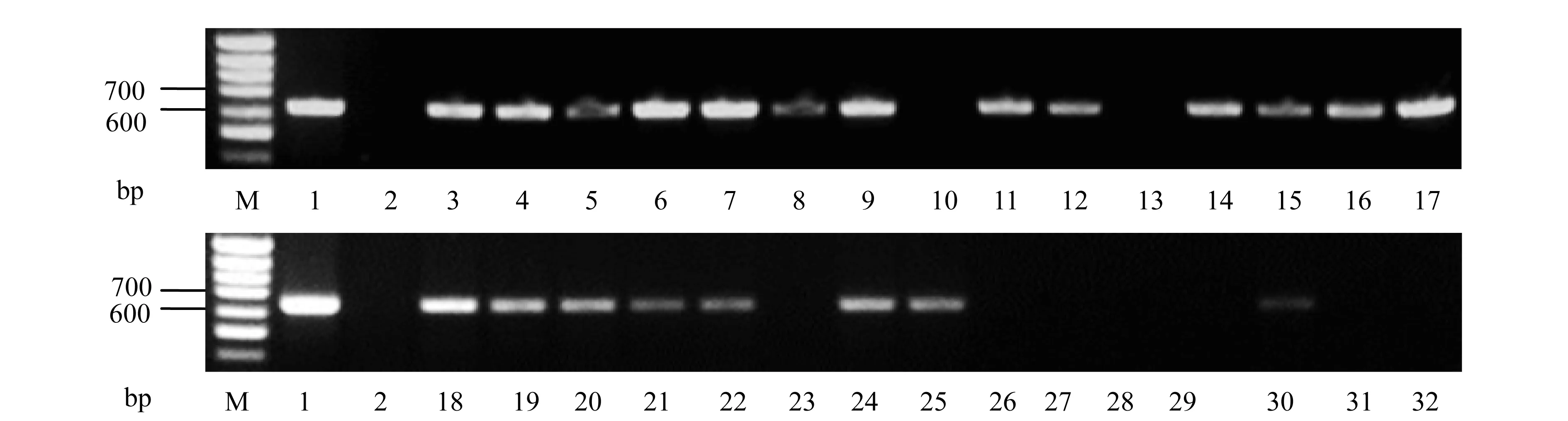
M:100 bp DNA Marker; 1:p13AWY-2质粒; 2:“93-11”对照水稻; 3-32:部分回交植株图2 部分回交后代植株反义蜡质基因PCR检测
2.2 改良后“93-11”水稻与对照主要农艺性状和拷种分析

A、B为9311主穗;C、D为2011-60-12植株主穗图3 “93-11”对照水稻与2011-59植株主穗比较
根据PCR检测结果,从2个小区各取4棵含有反义蜡质基因阳性植株,考察主要农艺性状并进行拷种分析.部分编号为2011-59类植株穗长度基本接近“93-11”对照水稻(图3).从平均值比较显示,2011-59类型植株每穗实粒数比对照低,但两者差异统计分析不显著.比较另外一些农艺性状和拷种性状的平均值,除了千粒重,两种改良后水稻的株高、有效穗和结实率也都与“93-11”对照水稻比较接近,统计分析无显著差异(表1).
2.3 改良后“93-11”水稻和对照糙米性状考察
剥去编号分别为2011-59和2011-60的BC4F2和BC3F3种子颖壳.根据糙米是否呈蜡质和非蜡质分离,可以判断植株反义蜡质基因位点是否为纯合状态.在含有反义蜡质基因的2011-59植株中,糙米都呈蜡质和非蜡质分离,即反义蜡质基因位点是杂合状态;在含有反义蜡质基因的2011-60植株中,糙米有纯合蜡质和杂合两种类型.
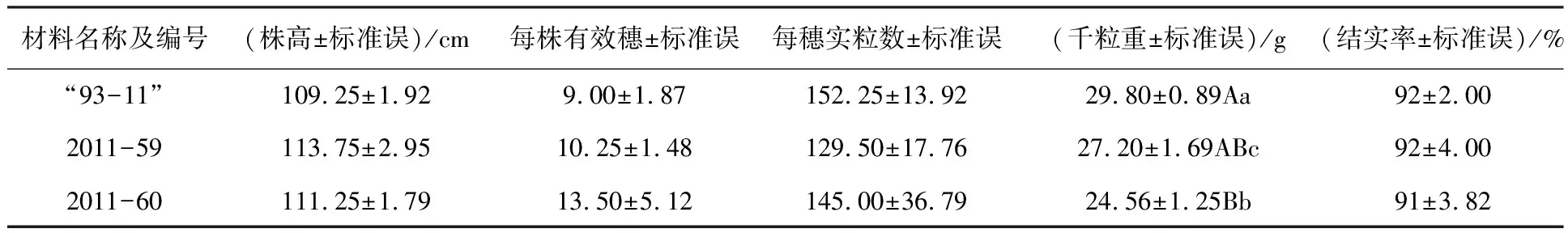
表1 “93-11”水稻及其改良后水稻主要农艺性状和拷种分析
注:同列表中不同大写和小写字母分别表示差异极显著(P<0.01)和显著(P<0.05),标有相同字母表示差异不显著(P>0.05),下同.
从2011-59小区中取含有反义蜡质基因植株2011-59-1,从编号为2011-60小区中取反义蜡质基因位点纯合植株:2011-60-12和“93-11”水稻植株,各随机取20粒糙米进行比较.结果显示,2011-60-12植株糙米明显小于“93-11”对照水稻,2011-59-1植株糙米大小比较接近于“93-11”对照水稻(图4).

图4 “93-11”对照(左)、回交4代反义蜡质基因杂合植株(中)和回交3代反义蜡质基因纯合植株(左)糙米比较
虽然从植株角度考察,2011-59-1植株的反义蜡质基因位点是杂合状态,但对于从2011-59-1植株上选出的蜡质糙米,反义蜡质基因位点已经纯合.再分别取40粒2011-59-1的蜡质和非蜡质糙米、40粒2011-60-12糙米,以及40粒“93-11”对照水稻糙米,分别测量糙米重量和体积.结果发现,两种改良后水稻糙米重量和体积都比“93-11”对照水稻极显著下降,而2011-59-1植株蜡质和非蜡质糙米重量和体积都比2011-60-12糙米极显著增加.在同一棵2011-59-1植株上收获的蜡质糙米比非蜡质糙米,重量和体积也都极显著下降(表2).
2.4 改良后“93-11”水稻和对照糙米部分理化指标分析
分析2011-59-1植株蜡质和非蜡质糙米、2011-60-12蜡质糙米,以及“93-11”对照水稻糙米的直链淀粉含量.结果显示,从编号为2011-59-1植株中选出非蜡质糙米的直链淀粉含量比“93-11”对照水稻略有下降,但统计分析差异不显著.两种蜡质糙米直链淀粉含量都比“93-11”对照水稻极显著地降低(P<0.01),降低幅度都超过了60%以上.2011-59-1植株非蜡质糙米胶稠度与“93-11”对照水稻也没有显著差异,两种蜡质糙米胶稠度都比“93-11”对照水稻极显著地增加(P<0.01),而且上升的幅度至少大于92%以上(表3).
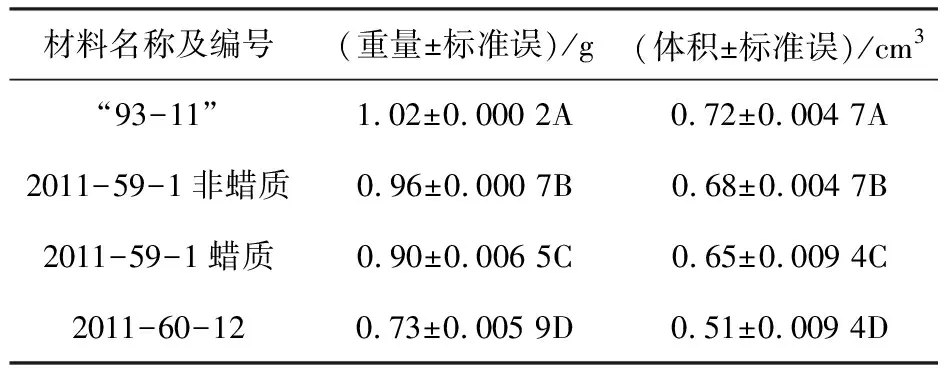
表2 “93-11”对照及其改良后水稻40粒糙米重量和体积比较

表3 “93-11”及其改良后水稻植株糙米部分理化指标分析
3 讨 论
杂交水稻稻米食味品质优劣与亲本稻米食味品质直接相关[18-19].本研究通过杂交及多代回交的方式将反义蜡质基因成功导入“93-11”水稻,不仅降低了稻米的直链淀粉含量,而且在更大程度上极显著地提高了“93-11”水稻稻米的胶稠度.胶稠度直接影响稻米柔软程度,高胶稠度稻米蒸煮的米饭更柔软,具有更好的食味品质[20-21].由本研究结果表明,改良后的“93-11”水稻,其稻米柔软程度会明显高于原“93-11”对照水稻.将改良后的“93-11”水稻作为父本与原先两系不育系杂交,由此获得的杂交组合稻米也将会有更好的食味品质.
随着分子标记技术的发展,目前已有较多研究者采用分子标记技术辅助改良水稻品质[6,22-23].胡德辉等[24]采用杂交及回交与分子标记辅助结合的方法,对水稻保持系“天B”和“龙特甫B”进行改良,选育出既保持原品种水稻的优良农艺性状,稻米品质又得到明显改良的“天B”和“龙特甫B”.侯立恒等[25]将供体水稻控制直链淀粉的Wx基因导入到不同恢复系中,得到了直链淀粉含量适中、并且主要农艺性状没有发生显著改变的后代.本文以含有反义蜡质基因的“B3”水稻为供体,目前生产上已广泛应用的“93-11”水稻为受体,同样利用杂交及回交和反义蜡质基因分子标记辅助筛选,获得了大多性状已经与“93-11”受体水稻非常接近的改良型“93-11”水稻.
Tian等[26]针对一般粳型或籼型水稻稻米直链淀粉与胶稠度比较分析认为,稻米直链淀粉含量与胶稠度之间存在着非常明显的反相关关系.Li等分析导入了反义蜡质基因的粳型水稻稻米直链淀粉含量与胶稠度关系时发现,反义蜡质基因对稻米胶稠度增加的幅度比降低稻米直链淀粉含量程度更明显[11].本研究通过杂交及多代回交将反义蜡质基因导入籼型水稻的研究发现,反义蜡质基因能够更加明显增加稻米胶稠度的作用,在籼型水稻中的作用效果与粳型水稻相同.
参考文献:
[1] 朱昌兰,沈文飚,翟虎渠,等.水稻低直链淀粉含量基因育种利用的研究进展[J].中国农业科学,2004,37(2):157-162.
[2] 舒庆尧,吴殿星,夏英武,等.稻米淀粉RVA谱特征与食用品质的关系[J].中国农业科学,1998,31(3):25-29.
[3] 隋炯明,李欣,严松,等.稻米淀粉 RVA谱特征与品质性状相关性研究[J].中国农业科学,2005,38(4):657-663.
[4] 张小明,石春海,富田桂.粳稻米淀粉特性与食味间的相关性分析[J].中国水稻科学,2002,16(2):157-161.
[5] SMITH A M,DENYER K,MARTIN C R.What controls the amount and structure of starch in storage organs?[J].Plant Physiol,1995,107(3):673-677.
[6] 刘巧泉,蔡秀玲,李钱峰,等.分子标记辅助选择改良特青及其杂交稻米的蒸煮与食味品质[J].作物学报,2006,32(1):64-69.
[7] 陈秀花,刘巧泉,王兴稳,等.反义Wx基因导入我国籼型杂交稻重点亲本[J].科学通报,2002,8(14):684-689.
[8] 李建粤,毛万霞,杨丽君,等.利用安全性转基因技术导入反义蜡质基因降低稻米直链淀粉含量[J].科学通报,2004,49(24):2556-2561.
[9] 沈革志,王新其,殷丽青,等.通过共转化和花药培养快速获得直链淀粉含量降低且无抗性标记的转基因水稻[J].植物生理与分子生物学学报,2004,30(6):637-643.
[10] 丁盛,刘清,黄志刚,等.反义Waxy基因转化籼稻9311的初步研究[J].湖南农业大学:自然科学版,2009,35(4):366-368.
[11] LI J Y,XU S Z,YANG L J,et al.Breeding elite japonica-type soft rice with high protein content through the introduction of the anti-Waxy gene[J].African Journal of Biotechnology,2009,8(2):161-166.
[12] YANG R,ZHOU Y G,CAO Y Q,et al.The transformation of the photo-thermo sensitive genic male-sterile line 261S of rice via an expression vector containing the anti-Waxy gene[J].Breeding Science,2013,63(2):147-153.
[13] ABE T,FUTSUHARA Y.Genotypic variability for callus formation and plant regeneration in rice (OryzasativaL.)[J].TheorAppl Genet,1986,72(1):3-10.
[14] 杨丽君,戎益泉,张善明,等.植物安全性表达载体的构建策略:以表达水稻反义蜡质基因的载体构建为例[J].分子植物育种,2009,7(5):1027-1031.
[15] 杨瑞.利用含有双份反义蜡质基因拷贝的表达载体改良水稻食味品质的研究[D].上海:上海师范大学,2010.
[16] 朱映东,时亚琼,周锋利,等.分子标记辅助选育香型巨胚水稻[J].上海师范大学学报:自然科学版,2013,42(6):623-628.
[17] 毛根武,董德良,杨瑞征,等.米饭质构特性测定方法的研究(Ⅲ)——米饭质构测试的最佳条件探讨[J].中国粮油学报,2013,28(9):6-10.
[18] 李欣,汤述翥,印志同,等.粳型杂交稻米品质性状的表现及遗传控制[J].作物学报,2000,26(4):411-419.
[19] 徐辰武,莫惠栋,张爱红,等.籼-粳杂种稻米品质性状的遗传控制[J].遗传学报,1995,22(3):192-198.
[20] 迟明梅.大米食用品质的研究进展[J].2005,30(1):48-51.
[21] 陈能,罗玉坤,朱智伟,等.优质食用稻米品质的理化指标与食味的相关性研究[J].中国水稻科学,1997,11(2):70-76.
[22] CHEN S,LIN X H,XU C G,et al.Improvement of bacterial blight resistance of ‘Minghui 63’,an elite restorer line of hybrid rice,by molecular marker-assisted selection[J].Crop Sci,2000,40(10):239-244.
[23] CHEN S,XU C G,LIN X H,et al.Improvement of bacterial blight resistance of ‘6078’,an elite restorer line of hybrid rice,by molecular marker-assisted selection[J].Plant Breeding,2001,120(2):133-137.
[24] 胡德辉,刘开雨,周萍,等.分子标记辅助选则改良天B和龙特甫B稻米品质[J].分子植物育种,2013,11(5):486-493.
[25] 侯立恒,夏明元,戚华雄,等.利用Wx基因分子标记辅助选择技术培育中等直链淀粉含量的水稻恢复系[J].中国农学通报,2009,25(14):32-36.
[26] TIAN Z X,QIAN Q,LIN Q,et al.Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities[J].Proc Natl Acad Sci USA,2009,106(51):21760-21765.
