中街山列岛海洋保护区岛礁生境鱼类资源及群落多样性季节变化*
2014-03-19王伟定虞宝存张洪亮
梁 君 王伟定 虞宝存 张洪亮
(浙江省海洋水产研究所 农业部重点渔场渔业资源科学观测实验站 浙江省海洋渔业资源可持续利用技术研究重点实验室舟山 316100)
大陆架浅海区分布有适宜鱼类生存的各种栖息地, 支持了数量众多、种类多样的生物资源, 每种栖息地都对某些特定鱼种的繁衍起着不可替代的作用(Gillanderset al, 2003)。这种为某些鱼类种群的幼体或成体的摄食、生长、产卵和成熟提供依靠和保障,并在此过程中决定该种群的大小和群体补充是否成功的栖息地被称为鱼类关键生境(Benaka, 1999)。世界上已建的海洋生物保护区有河口型、珊瑚礁型、海洋型、岛礁型和海岸型等5种类型, 其中, 岛礁型海洋保护区的数量仅次于海岸型而居于第2位(徐嵩龄等, 1995)。在当前国家重视海岛及海岸带整治修复的大背景下, 有必要深入研究岛礁生境的鱼类资源。我国关于天然岛礁生境鱼类资源的调查与研究多见于南海诸岛, 尤以西沙群岛(孙典荣等, 2005a, b; 史赟荣等, 2010)和南沙群岛为重点(李永振等, 2003, 2004;邵广昭等, 2011), 而东海区仅有郁尧山等(1986a,1986b)在1981年对浙江北部岛礁(黄龙岛和蚂蚁岛)周围海域鱼类资源进行了报道, 所使用的网具是张网; 在东海区其他生境研究方面, 国内研究者主要针对人工鱼礁(汪振华等, 2010; 赵静等, 2010)、潮下带岩礁(汪振华等, 2012)、沙地(汪振华等, 2011a)及多种生境组合(汪振华等, 2011b; 王蕾等, 2011)等方面进行了大量报道。
中街山列岛海洋保护区位于舟山群岛新区东部,处于舟山渔场的中心区域, 历史上素有“鱼仓”之美誉。该海域流系复杂, 一方面为长江、钱塘江径流所控制, 另一方面受南下的黄海水团和北上的台湾暖流的交汇影响, 大陆径流和长江口的“丛生流”给海区带来了大量的营养物质, 其水体交换快, 饵料充裕,是众多海洋生物繁殖、摄食和生长的重要栖息地(徐开达等, 2012)。继2006年5月中街山列岛国家级海洋特别保护区成立以来, 该海域逐渐成为渔业管理部门和相关科研单位关注的焦点, 并一致希望通过多方努力来保护并维持这一特殊海域的生态系统多样性, 近年海洋牧场暨碳汇渔业实验区建设项目正在该保护区内实施。中街山列岛海洋保护区所在的海域为典型的岛礁区(吴常文等, 1995), 对于该海域的相关研究多见诸于海藻(王志铮等, 2002; 孙建璋等,2006)、产卵场(唐逸民等, 1984)、水质环境(阳丹等,2012)等的调查与分析, 对于专门针对该保护区鱼类资源调查的资料少见(张龙等, 2012)。因此, 本文基于2年8个航次定置刺网调查数据, 对中街山列岛海洋保护区岛礁生境鱼类资源及群落多样性进行研究,以丰富我国鱼类群落生态学内容, 为保护区渔业资源的可持续利用和管理及海洋牧场建设提供科学依据。
1 材料与方法
1.1 研究区域和站点布设
2011年2月(冬)、5月(春)、8月(夏)、11月(秋)(I年度)和2012年2月、5月、8月和11月(II年度)在中街山列岛海洋保护区进行了8个航次的定置刺网调查, 调查范围如图1所示。该海域海洋性气候明显,四季分明, 常年为正规半日潮。本研究中, 根据海洋牧场建设主体区随机站点和捕捞干扰最小化原则,沿潮流主轴设置定置刺网采样带: 庙子湖岛西侧为贻贝养殖区(St.01), 庙子湖岛东侧为海藻床修复区或拟修复区(St.02), 青浜岛西南侧为网箱养殖区或拟建区(St.03), 青浜岛北侧为人居区和临时筏式养殖区(湾口)(St.04), 4个样带分别代表了该保护区的大部分栖息类型, 青浜岛东侧中部为沙地生境, 但由于流速太大, 定置刺网无法作业, 故未作考虑。
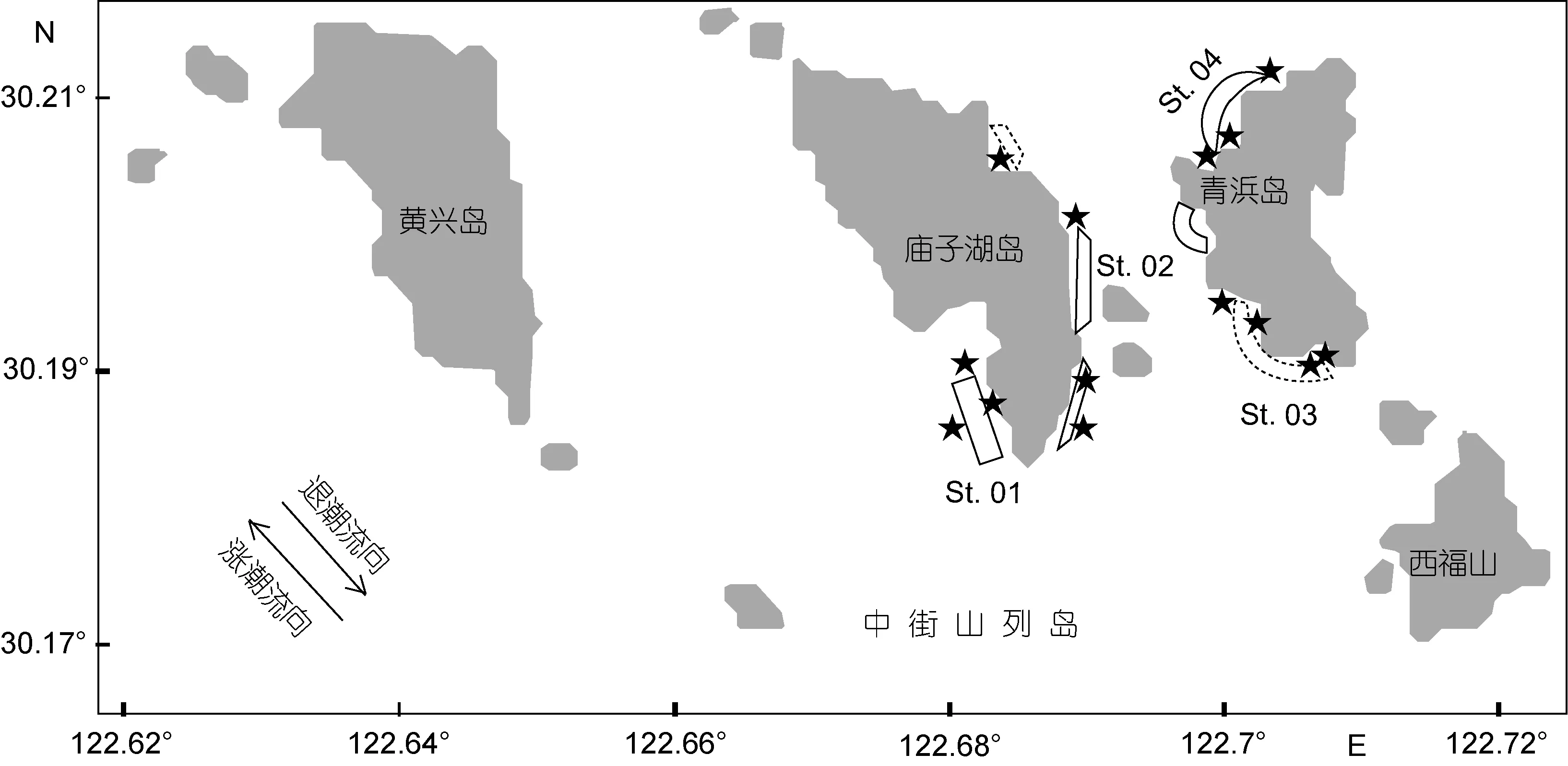
图1 采样区域和站点分布Fig.1 The study area and distribution of stations
1.2 采样方法和样品处理
调查船为浙普渔4126号, 该船长19m、宽3.3m,主机功率58.8kW; 调查时间为每航次(大潮汛来临前)3d, 每个季节均对4个采样带进行调查, 各点的定置刺网放置时间约为24h, 覆盖了昼夜2个时间段,受天气和海况等外界因素影响, 少数采样点和采样时间出现小幅波动。定置刺网为三重刺网, 单片网衣长45m、高1.2m, 中间网衣网目尺寸5cm, 两侧网衣网目尺寸各21cm, 5片为一组。作业时网具两端系有沉石, 以使其下缘触底并固定渔具, 起网时偏顺流起网, 并始终保持网列在起网舷一侧。此外, 进行鱼类样品采集的同时, 采用CTD仪(SEB-37SM)采集温度与盐度数据, 每航次每采样带各1次。对所得鱼类样品进行全部取样, 种类鉴别至其分类地位最低单元, 样品分析按《海洋调查规范》(GB/T 12763.6—2007)(国家质检总局等, 2007)进行, 体质量与体长精度分别为0.1g和1mm。
1.3 数据处理和分析方法
1.3.1 生态类群特征划分 为全面了解鱼类组成的功能群特征, 所有鱼类按适温性分为暖温种、暖水种和冷温种; 按其分布水层, 分为底层、近底层(或中下层, 下同)和中上层(汪振华等, 2012)。
1.3.2 相对资源丰度 鱼类相对丰度(RA, relative abundance)和相对生物量(RB, relative biomass)统一换算成该季度持续放置24h每一网次的平均渔获数量和平均渔获生物量(汪振华等, 2011a, 2012)。
1.3.3 生态优势度 利用Pinkas相对重要性指数(IRI)(Pinkaset al, 1971)确定鱼类在群落中的重要性。
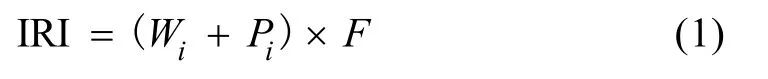
式中,Wi为某种渔获物的重量占总渔获重量的百分数(%),Pi为某种渔获物尾数占总渔获尾数的百分数(%),F为某种渔获物在各航次定置刺网总次数中出现的频率, 即出现次数与总网次数之百分比(%)。
1.3.4 群落的季节更替指数AI和迁移指数MI(金显仕等, 2009)
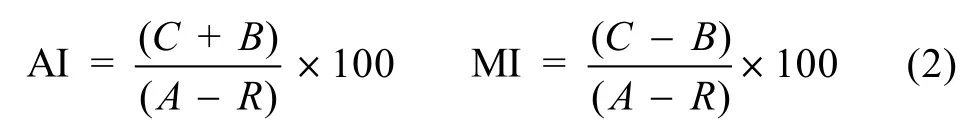
式中,A为各季节调查实际种类数;C为该季节调查新迁入种类数;B为该季节调查迁出种类数;R为年度调查中均出现的种类数。AI反映鱼类季节更替导致群落稳定性降低的节律, 其值越大则表示群落稳定性小; MI定义为研究系统外迁入与迁出种的2种成分在群落中的相对比例,C>B时, MI为正, 即表示迁入种大于迁出种, 当C接近于B时, MI为0, 意味着群落动态平衡。
1.3.5 Margalef丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数 采用Margalef丰富度指数(D) (Margalef, 1968)、Shannon-Wiener多样性指数(H′) (Whil, 1968)和Pielou均匀度指数(J′) (Pielou,1966)来研究调查站点出现鱼类的多样性:
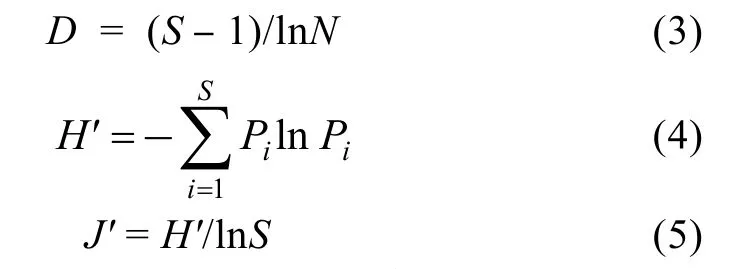
式中,S为种类数,N为总尾数,Pi为第i种占总尾数的比例。
1.3.6 数据处理 建立“种类×采样带”丰度矩阵(使用原始丰度数据, 为保证采样信息的真实性, 不作任何转化), 计算各采样带间的Bray-Curtis相似性系数以形成相似性矩阵, 采用等级聚类(UPGMA)和非度量多维标度(nMDS)作为鱼类群落的分类和排序方法, 结合两者分析各采样带的鱼类群落格局, 并采用相似性分析(ANOSIM)检验不同格局间差异的显著性。以上聚类、排序及多样性分析均在在PRIMER v6软件中实现。显著性检验与相关性分析分别采用SPSS 19.0的单因素方差分析法(ANOVA)和曲线回归分析法(CURVE ESTIMATION), 以上统计显著水平为0.05, 极显著水平为0.01。
2 结果与分析
2.1 种类组成和相对资源丰度变化特征
2011—2012年度在中街山列岛海洋保护区岛礁生境4个主要采样带共放置28网次, 总计放网长度6300m, 采集各种鱼类2820尾, 总生物量105165g。采集到的鱼类隶属8目31科48属58种(表1, 附录I)。其中鲈形目鱼类共计26种, 占所有种类的44.8%,成为绝对优势门类; 其次是鲱形目、鲽形目和鲉形目,合计占所有种类的41.4%; 其他目中, 除了灯笼鱼目的龙头鱼(Harpodon nehereus)和鳗鲡目的海鳗(Muraenesox cinereus)在各个季度分别维持一定的捕捞数量和生物量外, 其他种类均较为单一, 且数量稀少(附录I)。
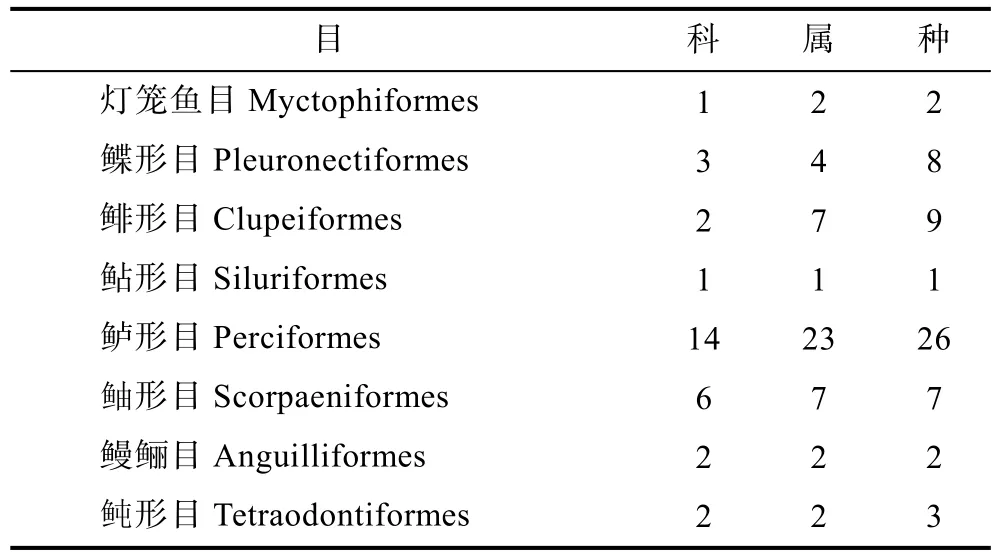
表1 中街山列岛海洋保护区鱼类数量组成Tab.1 Composition of fish in the protection area of Zhongjieshan Islands
不同生态类型的鱼类组成方面(表2), 该岛礁生境种类以暖温性和暖水性为主, 2种温度属性鱼类占所有种类的95%。其中, 暖温种数量除秋季稍偏低外,其他季节变化不明显; 暖水种具有明显季节变化特征, 与水温同步。从分布水层上看, 底层和近底层鱼类的数量差别不大, 而中上层鱼类数量在夏季明显多于其他季节。所有种类中, 趋礁性鱼类为46种, 占79%, 其种类构成中, 除了全部底层鱼类外, 还包括石首鱼科(如黄姑鱼(Nibea albiflora)、皮氏叫姑鱼(Johnius belengerii)和鱼(Miichthys miiuy)等)、鲷科(黑鲷(Sparus macrocephalus)、真鲷(Pagrosomus major)和黄鳍鲷(Sparuslatus Houttuyn))和鲀科(横纹东方鲀(Takifugu oblongus)和铅点东方鲀(Takifugu alboplumbeus))等营近底层生息的种类, 以及暖水性特征明显的种类(如褐篮子鱼(Siganus fuscescens)和花尾鹰(Goniistius quadricornis)等), 同时, 岛礁环境还吸引了以细纹狮子鱼(Liparis tanakai)为代表的多在深水层活动的种类。
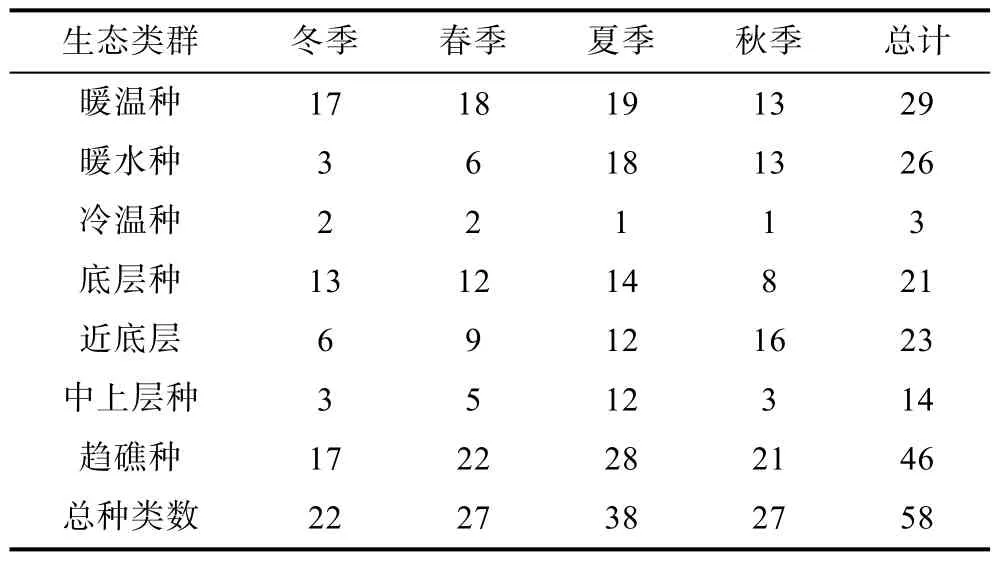
表2 保护区各种生态类群鱼类数量的季节变化Tab.2 Seasonal variations in fish species number in each eco-group in the protected area
从总种类和总网次2个方面比较了不同季节鱼类相对资源丰度的变化, 由图2可以看出, 2种不同方式求得的RA和RB与季节变化一致, 均为夏季最高, 春季其次, 冬季最低。如从单个种类对RA和RB的贡献角度来看, 秋、冬季褐菖鲉(Sebastiscus marmoratus)对RA贡献最大, 春、夏季鳀(Engraulis japonicus)对RA贡献最大(附录I), 而对RB贡献较大的种类主要为各季节的优势种。
对保护区岛礁生境各季节鱼类的种类数、RA和RB与该海域平均底层温度之间的关系进行耦合分析,曲线回归分析结果显示, RB与温度之间呈复合曲线关系(图3), 相关性显著(P=0.022<0.05), 而RA和种类数分别与温度之间呈弱相关(P=0.049,P=0.044, <0.05)。
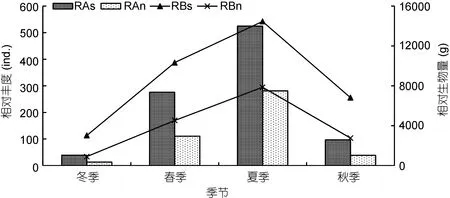
图2 不同季节鱼类相对资源丰度Fig.2 The relative resource abundance in each season
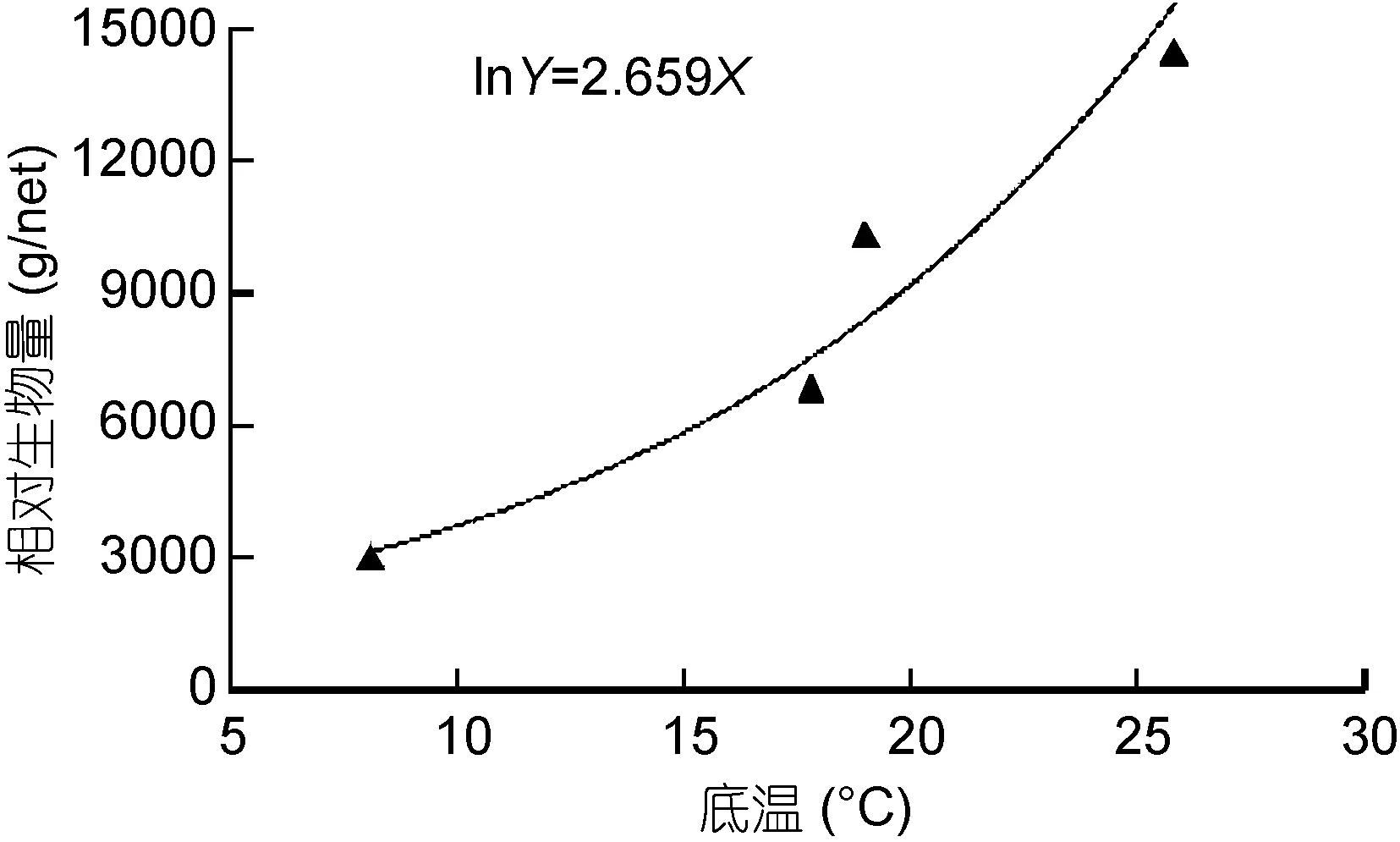
图3 相对生物量与平均底层温度的相关性Fig.3 Correlation between relative biomass and average sea bottom temperature
2.2 主要优势种的季节变化
本文将IRI≥500, 且在≥50%网次捕获的鱼类定为优势种。中街山列岛海洋保护区岛礁生境按季节分组共采集优势鱼类13种, 其中底层鱼类5种, 近底层5种, 中上层3种(表3)。从2年度总优势度来看, 保护区出现4种优势鱼类, 其中底层1种, 近底层2种,中上层1种(表3, 附录I)。
优势种组成随水层的不同而变化, 底层优势种以趋礁性鱼类为主, 鲉形目鱼类主导着底层鱼类群落, 其中, 褐菖鲉是2个年度唯一优势种, 并在各季节均成为优势种, 特别是在每年的冬季和春季占有绝对优势; 近底层优势群体中全部为石首鱼科鱼类,其中皮氏叫姑鱼和黄姑鱼为2个年度的优势种, 从季节角度看, 小黄鱼(Larimichthys polyactis)、黄姑鱼和棘头梅童鱼(Collichthys lucidus)共同构成冬季主要优势种群, 在春季被唯一的优势种黄姑鱼替代, 皮氏叫姑鱼则成为夏季和秋季的绝对优势种; 中上层优势鱼类主要集中在春、夏季, 以季节性洄游性鱼类(鳀、赤鼻棱鳀(Thrissa kammlensis)和日本鲭(Scomber japonicus))为主, 调查发现春季日本鲭和鳀在该岛礁区出现集群现象, 个别网次这2种鱼类占总种类数的80%以上, 而鳀(IRI=3563)成为夏季各水层中仅次于皮氏叫姑鱼(IRI=5004)的“极端”优势种(章守宇等,2008)。
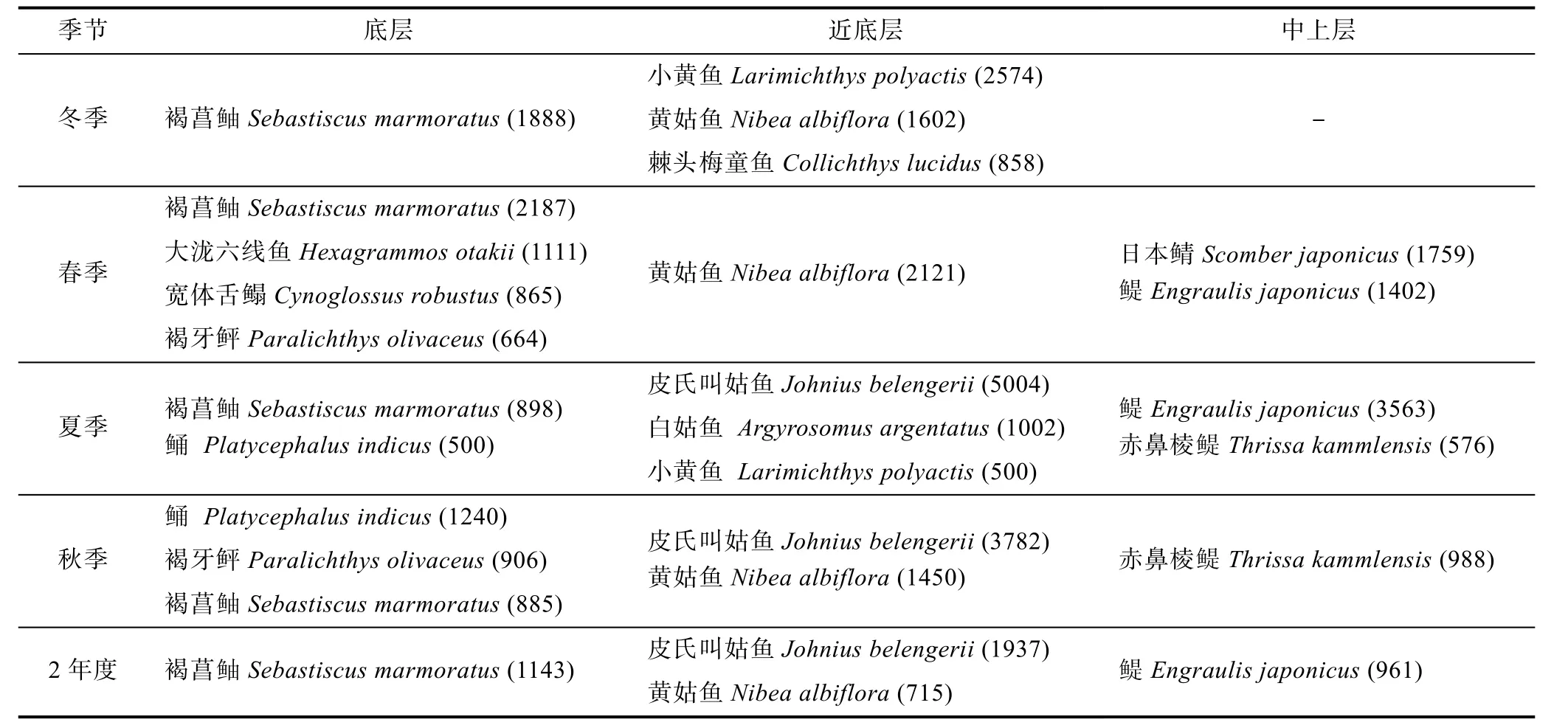
表3 保护区优势种的季节变化Tab.3 Seasonal variations of dominant species in the protected area
2.3 多样性特征的季节变化
从图4可以看出, 中街山列岛海洋保护区岛礁生境D值和H′值均为夏季最高, 秋季其次, 冬季最低,而各季节J′呈现相反的特点。ANOVA分析结果表明,就J′值而言, 冬季与夏季之间差异极显著(P=0.006<0.01), 与秋季之间具有显著性差异(P=0.022<0.05),而与春季没有显著性差异(P=0.176>0.05); 就D值和H′值而言, 各季节间差异均不显著, 最大值分别为最小值的1.24倍和1.09倍。
2.4 群落稳定性的季节变化

图4 保护区鱼类多样性指数均值的季节变化Fig.4 Seasonal variations of mean values of the diversity indices in the protected area
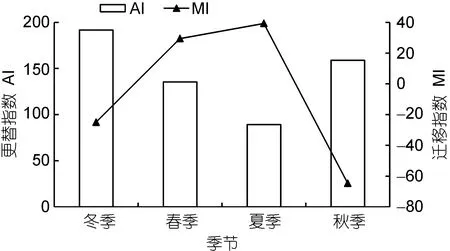
图5 各季节鱼类的更替指数(AI)和迁移指数(MI)Fig.5 The alternation index and migration index of fish in each season
各季节AI和MI值变化如图5所示, AI的年变化呈“v”型, 稳定性最差的是冬季, 其AI值高达192, 而夏季稳定性最好, AI值仅为89; MI的年变化呈“n”型,春季和夏季MI值为正, 但秋季和冬季为负, 说明季节变化引起的温度改变对岛礁生境的鱼类群落组成具有明显影响。
2.5 鱼类群落组成格局
从保护区鱼类群落聚类和排序结果来看(图6),形成4种不同的群落组成格局(GlobalR=0.797,P<0.01)。分别为以日本鲭和鳀为绝对优势种的中上层群落①, 以黄姑鱼为优势类群的近底层群落②, 以褐菖鲉为代表的底层群落③, 和以赤鼻棱鳀、皮氏叫姑鱼和为优势种的分层群落④, 且群落①、②和③皆为典型的冬、春季鱼类组成格局, 群落④为典型的夏、秋季鱼类组成格局, 格局①、②和③均与④存在极显著差异(GlobalR=0.907、0.877和0.731,P<0.01)。
3 讨论
3.1 鱼类组成、资源丰度与底层温度的关系
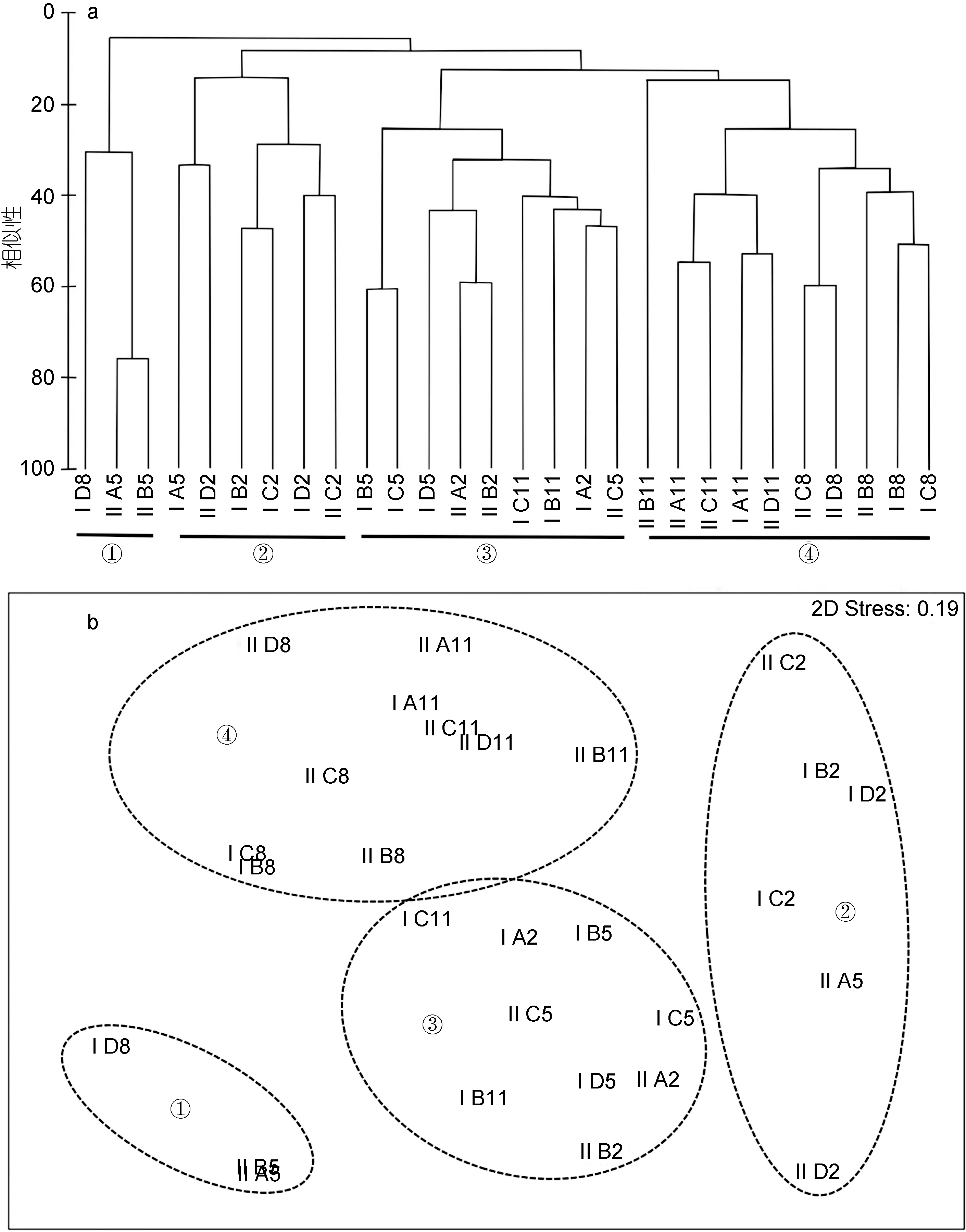
图6 基本丰度数据的各航次鱼类群落结构多元分析Fig.6 Multivariate analysis on fish community structures in each voyage based on fish abundance
从曲线回归分析结果可知, 中街山列岛海洋保护区鱼类生物量、丰度和种类数的高低与底层温度之间关系密切, 该结论进一步印证了暖水性鱼类与暖温性鱼类种类数相当且与水温同步的事实。图7列出了各季节鱼类区系组成随不同水温的变化趋势, 从种类所属区系百分比的季节变化来看, 冷温种在冬季和春季所占的比例稍高于夏季和秋季, 夏、秋季该岛礁生境对暖水种具有明显的诱集性, 比例增加明显, 此外, 暖水种同时也成为趋礁性鱼类的重要组分(表2, 附录I)。
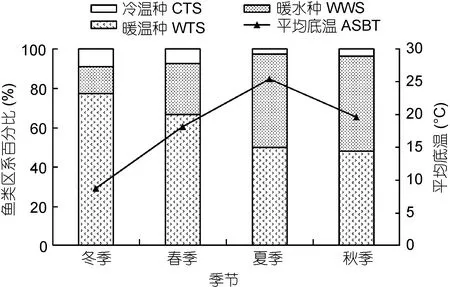
图7 各季节鱼类区系组成与平均底层温度的相关性Fig.7 Correlation between fish fauna composition and average sea bottom temperature in different seasons
3.2 影响保护区鱼类群落格局的内因
中街山列岛海洋保护区是传统经济种类如小黄鱼、带鱼、银鲳的主产区, 每年各季鱼汛期都有大量经济鱼类洄游经过(张龙等, 2012)。聚类和排序结果显示, 保护区鱼类格局主要受季节影响, 其次受控于不同水层优势种类。对不同采样带进行ANOSIM检验发现, GlobalR皆为负值, 差异均不显著(P>0.05),说明不同栖息类型对中街山保护区岛礁生境鱼类组成格局影响不大, 保护区鱼类群落组成更依赖于整个岛礁大环境, 且受季节变化影响更大, 这一结论与俞存根等(2010)提出的舟山渔场及邻近海域的鱼类以洄游性种类或季节性种类为主的结论一致。中街山列岛海洋保护区岛礁生境可维系趋礁性鱼类栖息, 还能吸引暖水性种类和多在深水层活动的种类, 说明该保护区是一个饵料丰富的小型生态系统, 同时也是良好的鱼类庇护场所。
3.3 岛礁生境维系鱼类物种多样性和稳定性的现状和趋势
中街山列岛海洋保护区岛礁D值和H′值均为夏季最高, 秋季其次, 冬季最低,J′值呈现相反规律, 研究结果与处于同一经度上的马鞍列岛岩礁生境的变化规律一致(汪振华等, 2012), 与1981年黄龙岛夏、秋季高于春、冬季的结论也相同(郁尧山等, 1986a)。本研究Shannon-Wiener多样性变化范围为0.76—2.58,平均值为1.61, 略高于2009年马鞍列岛的0.7—2.24(汪振华等, 2012)。从陆架水深跨度上来看, 远小于1981年东海深海底层鱼类群落(沈金鳌等, 1987)的H′值(平均值介于2.0—3.0)。从空间跨度上来讲, 本研究结果与刘勇等(2006)提出的同属东海区的北部和南部也存在差异, 南部多样性要高于北部的论点相一致。从时间跨度上来讲, 过度捕捞是造成岛礁生境鱼类多样性下降的主因(宋普庆等, 2012; 汪振华等,2012)。群落的稳定性是研究群落间相互作用过程的重要指标, 是群落成员在长期变化中维持基本特征的适应能力(金显仕等, 2009)。从AI值来看, 春季和夏季稳定性最好, 从MI值来看, 保护区春季和夏季有明显种类迁入的迹象, 特别是夏季暖水种明显高于其他季节(表2), 分析原因除了本文提及的温度因子的影响外, 还可能是一方面由于伏季全面休渔制度的实施, 另一方面可能更得益于近年来在该保护区内规划并实施的海洋牧场建设。调查结果发现, 该保护区内共捕获浙江省近年放流的鱼类8种, 分别为大黄鱼、黄鳍鲷、条石鲷(Oplegnathus fasciatus)、黑鲷、日本黄姑鱼(Nibea japonica)、黄姑鱼、褐牙鲆(Paralichthys olivaceus)和半滑舌鳎(Cynoglossus semilaevis), 其中, 定置刺网曾少量捕获到挂有标志牌的黄鳍鲷、条石鲷和黑鲷, 而日本黄姑鱼为东海近乎绝迹的品种, 2005年开始集中在舟山朱家尖情人岛周围海域(距离中街山列岛海洋保护区西南侧约24海里处)实施放流, 在保护区内捕获个体基本可认定为是放流种。此外, 从优势种组成上看, 黄姑鱼成为该保护区各季节优势种, 而近海洄游性的大黄鱼幼体作为偶见种亦在春季和秋季保护区内出现。汪振华等(2012)认为岛礁生境既是趋礁性鱼类终年依赖的关键生境, 又是各种洄游性鱼类幼体趋于季节性集中的重要栖息地, 对维系岛礁海域物种多样性和保护重要渔业资源群体起着不可忽视的生态作用。因此, 笔者期待有关部门加快保护区内海洋牧场建设步伐,进行岛礁生态环境的修复, 促进岛礁渔业的科学开发, 并开展针对性的管理措施, 以构建可持续性岛礁渔业和独特的海岛经济。
王志铮, 张义浩, 吴常文等, 2002. 中街山列岛底栖海藻的资源调查. 水产学报, 26(2): 189—192
王 蕾, 章守宇, 汪振华等, 2011. 枸杞岛近岸3种生境鱼类群落组成及岩礁区底栖海藻对鱼类群落结构的影响. 水产学报, 35(7): 1037—1049
刘 勇, 李圣法, 程家骅, 2006. 东海、黄海鱼类群落结构的季节变化. 海洋学报(中文版), 28(4): 108—114
孙典荣, 林昭进, 邱永松, 2005a. 西沙群岛重要岛礁鱼类资源调查. 中国海洋大学学报, 35(2): 225—231
孙典荣, 林昭进, 邱永松等, 2005b. 西沙群岛重要珊瑚礁海域鱼类区系. 南方水产, 1(5): 18—25
孙建璋, 余 海, 陈万东等, 2006. 浙江底栖海藻记录. 浙江海洋学院学报(自然科学版), 25(3): 312—321
阳 丹, 陈应华, 吴常文, 2012. 中街山列岛及其邻近海域海水质量评价. 浙江海洋学院学报(自然科学版), 31(1):49—53
李永振, 林昭进, 陈丕茂等, 2003. 南沙群岛中北部重要岛礁鱼类资源调查. 水产学报, 27(4): 315—321
李永振, 陈国宝, 袁蔚文, 2004. 南沙群岛海域岛礁鱼类资源的开发现状和开发潜力. 热带海洋学报, 23(1): 69—75
邵广昭, 陈正平, 陈静怡等, 2011. 南海东沙岛及太平岛鱼类种类组成和动物地理学特点. 生物多样性, 19(6):737—763
沈金鳌, 程炎宏, 1987. 东海深海底层鱼类群落及其结构的研究. 水产学报, 11(4): 293—306
宋普庆, 张 静, 林龙山等, 2012. 台湾海峡游泳动物种类组成及多样性. 生物多样性, 20(1): 32—40
汪振华, 章守宇, 王 凯等, 2010. 三横山人工鱼礁区鱼类和大型无脊椎动物诱集效果初探. 水产学报, 34(5):751—759
汪振华, 章守宇, 陈清满等, 2012. 马鞍列岛岩礁生境鱼类群落生态学. I.种类组成和多样性. 生物多样性, 20(1):41—50
汪振华, 王 凯, 赵 静等, 2011a. 枸杞岛潮下带沙地生境鱼类群落结构和季节变化. 应用生态学报, 22(5):1332—1342
汪振华, 王 凯, 章守宇, 2011b. 马鞍列岛多种生境中鱼类群聚的昼夜变化. 生态学报, 31(22): 6912—6925
吴常文, 吕永林, 1995. 中街山岛礁区渔业综合开发建议. 海洋开发与管理, 12(1): 68—71
国家质检总局, 国家标准化委员会, 2007. 海洋调查规范第6部分: 海洋生物调查. 北京: 中国标准出版社, 56—62
张 龙, 徐开达, 张洪亮等, 2012. 中街山海域渔业动物群落结构的季节变化. 浙江海洋学院学报(自然科学版), 31(4):290—294
金显仕, 单秀娟, 郭学武等, 2009. 长江口及其邻近海域渔业生物的群落结构特征. 生态学报, 29(9): 4761—4772
郁尧山, 张庆生, 陈卫民等, 1986a. 浙江北部岛礁周围海域鱼类群聚特征值的初步研究. 水产学报, 10(3): 305—313
郁尧山, 张庆生, 陈卫民等, 1986b. 浙江北部岛礁周围海域鱼类优势种及其种间关系的初步研究. 水产学报, 10(2):137—149
俞存根, 陈全震, 陈小庆等, 2010. 舟山渔场及邻近海域鱼类种类组成和数量分布. 海洋与湖沼, 41(3): 410—417
赵 静, 章守宇, 汪振华等, 2010. 嵊泗人工鱼礁海域鱼类和大型无脊椎动物群落结构特征. 中国水产科学, 17(5):1045—1056
徐嵩龄, 孙建璋, 钟晓东等, 1995. 岛礁型海洋生物保护区(IMPA)的设计和管理: 理论和实例研究. 生态学报, 15(1):95—103
徐开达, 张洪亮, 谢汉阳等, 2012. 中街山列岛水域甲壳类资源及其群落多样性. 海洋渔业, 34(3): 308—315
唐逸民, 郑佩玉, 李永明等, 1984. 中街山曼氏无针乌贼产卵场生态环境及其资源保护. 浙江水产学院学报, 3(2):132—140
章守宇, 梁 君, 汪振华等, 2008. 浙江马鞍列岛海域潮间带底栖海藻分布特征. 应用生态学报, 19(10): 2299—2307
Benaka L R, 1999. Fish Habitat: Essential Fish Habitat and Rehabilitation. American Fisheries Society Symposium,Bethesda, US: 1—22
Gillanders B M, Able K W, Brown J Aet al, 2003. Evidence of connectivity between juvenile and adult habitats for mobile marine fauna: an important component of nurseries. Marine Ecology Progress Series, 247: 281—295
Margalef R, 1968. Perspectives in Ecological Theory. University of Chicago Press, Chicago, US: 1—111
Pielou E C, 1966. The use of information theory in the study of ecological succession. Journal of Theoretical Biology, 10:370—383
Pinkas L, Oliphant M S, Iverson I L K, 1971. Food habits of albacore, bluefin tuna, and bonito in California waters. Fish Bulletin, 152: 1—105
Whil M J L, 1968. Use of biomass units in Shannon’s formula.Ecology, 49: 153—156
