饲粮能量水平对杜泊羊养分利用率和产热量的影响
2014-03-19张崇玉赵敏孟杨在宾杨维仁姜淑贞张桂国朱文广张吉林
张崇玉,赵敏孟,杨在宾,杨维仁,姜淑贞,张桂国,朱文广,张吉林,巩 峰
1.山东农业大学动物科技学院,泰安 271018 2.山东凯银集团股份有限公司,东营 257000)
研究动物产热量的方法有直接测热法和间接测热法,直接测热法原理简单,但测热室的制作技术却很复杂,造价也很昂贵;相反呼吸面具法设备简单,造价低,而且适合于野外测定,是最简单易行的开路式间接测热法[1]。杜泊羊原产南非,因其适应性强、早期生长发育快、遗传性稳定、胴体质量好而闻名于世。杜泊羊推广到我国温带各气候类型地区,都表现出良好的适应性,耐热抗寒,尤其是突出的产肉性能是目前我国引进的和国产的其他肉用绵羊品种都不可比拟的[2]。测定杜泊羊的养分利用率和产热量,进而估算能量需要量,可以为杜泊羊饲养标准的制定提供科学数据,同时也为我国肉羊饲养标准的修订提供有关参数。长期以来,动物营养学家都致力于动物产热量的研究,Miron等[3]和Meccawi等[4]分别测定了奶牛和绵羊、山羊的产热量,杨诗兴等[5]测定了湖羊的产热量。但是关于饲粮不同能量水平对杜泊羊养分利用率和产热量规律的影响却未见报道。(拟解决的关键问题)本试验旨在通过消化代谢试验和呼吸测热试验相结合的方法,研究不同能量水平对杜泊羊养分利用率和产热量的影响,为测定杜泊羊的维持代谢能(Metabolism energy for maintenance,MEm)提供参数,同时也为我国肉羊饲养标准的修订提供有关参数。
1 材料与方法
1.1 试验动物及试验设计
试验采用2× 3析因试验设计,即2种性别(公羊和母羊)× 三种能量水平(低、中、高)。选取断奶后15 d纯种杜泊羊18只,公母各半,平均体重:公羊为(50.97± 0.89)kg,母羊为(48.33± 0.22)kg,公母各分为3个处理组,每个处理3个重复,每个重复1只。试验饲粮设计参考NRC(2007)绵羊饲养标准[6],分别按其推荐量的90、100和110%供给能量,饲粮消化能水平分别为:9.78、10.66、11.62 MJ/kg。其他饲粮组分和营养水平一致。饲粮组成及营养水平见表1。
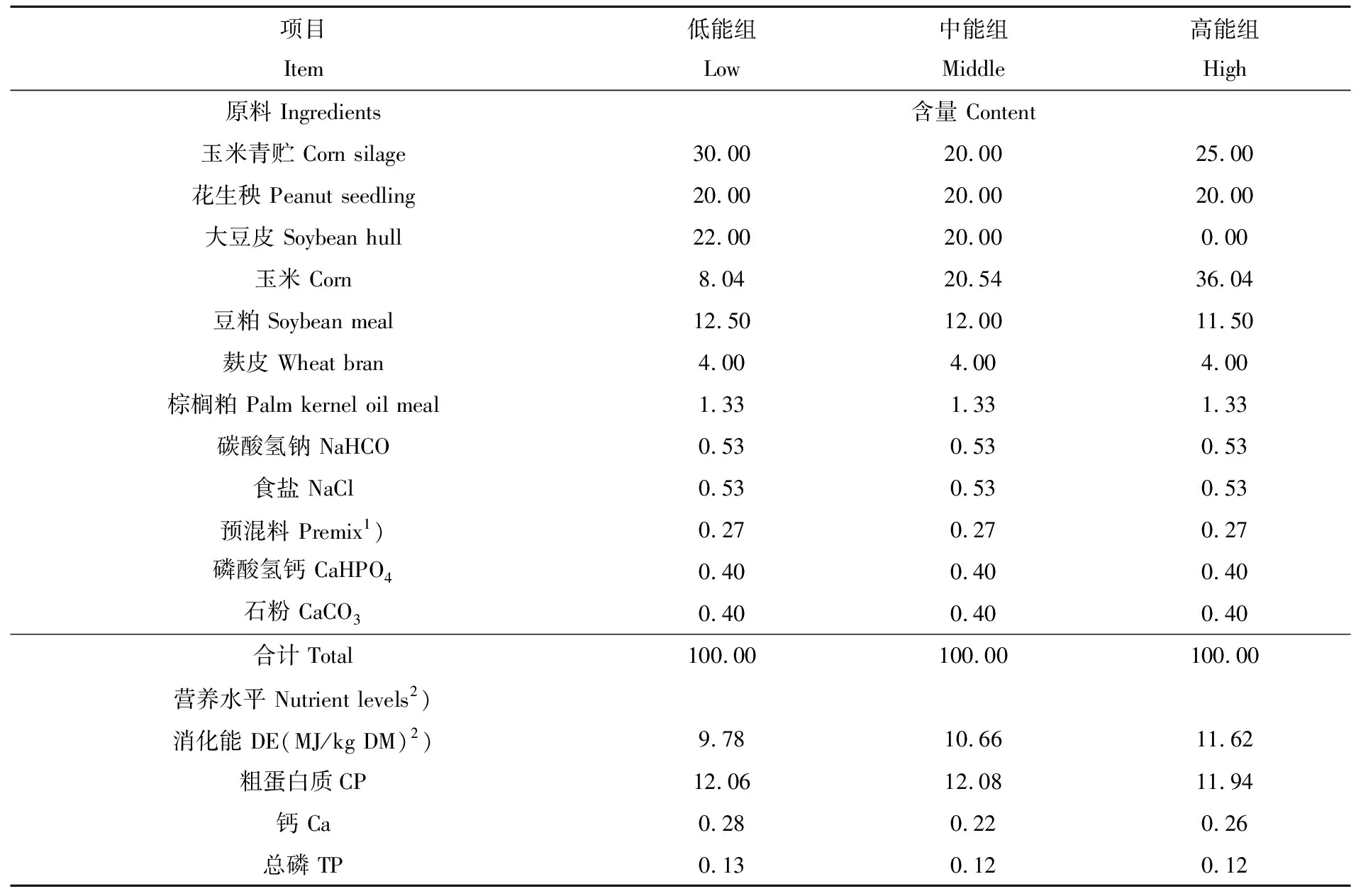
表1 基础饲粮组成及营养水平(风干基础) %
1)预混料为每千克饲粮提供The premix provided the following per kg of diets:VA 1250 KIU,VD3270 KIU,VE 5000 IU,烟酸 nicotinic acid 2000 mg,Mn 3000mg,Fe 3170 mg,Zn 14280 mg,Cu 3040 mg,I 180 mg,Se 100 mg,Co 40 mg。
2)消化能为计算值,其余为实测值。DE was a calculated value and others were measured values.
1.2 试验方法
1.2.1 消化代谢试验 消化代谢试验采用全收粪、尿法,在专用消化代谢笼中进行。试验预试期7 d,正试期7 d,定时投料,自由饮水。试验采用限饲法,控制每只试验羊采食量基本一致。正试期时每天准确记录每只试验羊的采食量。每天08:00由专用消化代谢笼的收粪盘和收尿桶收取粪尿,并准确称量,4℃保存。试验结束,每只试验羊均匀混合7 d所收粪样。分2步进行取样:①取鲜粪样50 g,加入10%的盐酸10 mL用于测定鲜粪状态下粗蛋白质;②另取500 g鲜粪样,烘干制成风干样用以测定其他成分;正试期每天收集尿量总体积的10%并混合均匀,供营养成分分析。饲料和粪、尿中的干物质和能量参考贺建华[7]方法进行测定,包括干物质(DM)、有机物(OM)、粗灰分(CA)、粗蛋白(CP)、中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)和总能(GE)的测定。
1.2.2 呼吸测热试验 呼吸测热试验与消化代谢试验同步进行。采用传统开路式呼吸面具测热法,步骤参考Blaxter[8]的方法,预试期时每天训练羊采气,正试期每天分别于06:00、09:00、11:00、15:00、18:00和21:00采气6次,每次采气5 min(采气时全部试验羊均为站立状态),使用呼吸面具将呼出气体收集于专用贮气囊内,用LML-1型湿式气体流量计测定呼出气体总量,用取样袋采取气样,即用SB67-1型半自动气体分析仪测定气样中CO2和O2浓度,按所得百分比求出呼吸墒(RQ);因气体分析器的燃烧瓶灵敏度不高,故甲烷气体产生量及其能值按Blaxter公式[9]推算。产热量的计算采用RQ法,根据单位时间内呼出气体的总量(用气态方程换算成标准状态体积)求出CO2排出量和O2消耗量,根据每升O2与RQ值相对应的氧热价,求出被试羊一天的总产热量。试前对气体分析仪、气体流量计均经严格校验。正试期时记录气温、气压、相对湿度等环境指标。
产热量的计算还采用Brouwer(1965)[10]公式进行估测。产热量HP=3.866O2+1.200CO2-0.518CH4-1.431N,式中,O2、CO2、CH4和N分别为单位时间内的耗氧量、二氧化碳排出量、甲烷气体产生量和尿氮排出量。
体蛋白分解量/g=尿氮量×6.25;蛋白质产热量/kJ=体蛋白分解量×18 kJ/g;蛋白质CO2排出量/L=体蛋白分解量×0.78;蛋白质O2消耗量/L=体蛋白分解量×0.97;非蛋白质CO2排出量/L=CO2排出量-蛋白质CO2排出量;非蛋白质O2消耗量=O2消耗量-蛋白质O2消耗量,CO2排出量和O2消耗量由呼吸测热试验测得[11]。
1.3 数据统计分析
数据采用SAS 9.1软件进行统计学处理,方差分析采用2×3的析因方差分析,P< 0.05者为差异显著。
2 结果与分析
2.1 环境指标
试验期平均气温为24.2 ℃,相对湿度为65.7%,气压99.48 kPa。
2.2 饲粮能量水平对杜泊羊生长期养分利用率的影响
饲粮能量水平对杜泊羊养分利用率的影响见表2。由表可知,性别和能量水平的交互作用对杜泊羊DM、OM、CP、NDF、ADF的表观消化率的影响均不显著(P> 0.05),对粗CP的表观代谢率的影响也不显著(P> 0.05)。公羊对于DM、OM、CP的表观消化率极显著高于母羊(P< 0.01),但NDF和ADF的表观消化率公母羊之间差异不显著(P> 0.05)。中能组和高能组的DM、OM、NDF和ADF的表观消化率显著高于低能组(P< 0.05),这表明提高饲粮能量水平可以提高杜泊羊对DM和OM的利用率,但到达一定能量水平后则不在增加。不同能量水平间CP的表观消化率和代谢率差异不显著(P> 0.05),这表明本试验设置的能量梯度范围内,能量水平并没有影响杜泊羊对CP的利用率。
2.3 饲粮能量水平对杜泊羊生长期气体产量的影响
饲粮能量水平对杜泊羊气体产量的影响见表3。由表可知,性别和能量水平的交互作用对杜泊羊CO2排出量、O2消耗量、非蛋白质CO2排出量、非蛋白质O2消耗量、蛋白质CO2排出量、蛋白质O2消耗量以及CH4产量的影响均不显著(P> 0.05)。公羊的CO2排出量极显著高于母羊(P< 0.01),可以看出是因为蛋白质CO2排出量公羊高于母羊的结果,其根本的原因可能是因为公羊的尿氮排出量高于母羊的缘故。不同能量水平之间对比可以看出,中能组的CO2排出量、O2消耗量均极显著高于其他两组(P< 0.01),从表中可以看出是因为非蛋白质CO2排出量、非蛋白质O2消耗量中能组显著高于其他两组的缘故,不同能量水平之间蛋白质CO2排出量、蛋白质O2消耗量之间差异不显著(P> 0.05),其可能原因是因为不同能量水平之间尿氮量差异不显著。
2.4 饲粮能量水平对杜泊羊生长期产热量和呼吸熵的影响
饲粮能量水平对杜泊羊产热量和呼吸熵的影响见表4。由表可知,性别和能量水平的交互作用对杜泊羊单位代谢体重的产热量、蛋白质产热量、非蛋白质产热量、呼吸熵、非蛋白呼吸熵的影响均不显著(P> 0.05)。单位代谢体重的产热量公母羊之间差异并不显著,但蛋白质产热量公羊极显著高于母羊(P< 0.01),非蛋白质产热量母羊极显著高于公羊(P< 0.01);蛋白质产热量占总产热量的百分比公羊为16.02%,母羊为13.88%,公羊极显著高于母羊(P< 0.01)。公羊的呼吸熵与非蛋白质呼吸熵均显著高于母羊(P< 0.05)。不同能量水平之间对比可以看出,中能组和高能组的产热量显著高于低能组,这说明提高能量水平提高了动物的产热量,分析原因可以看出这来自非蛋白产热量的不同,各组间蛋白质产热量差异并不显著(P> 0.05)。蛋白质产热量占总产热量的百分比不同能量水平之间差异不显著(P< 0.05),平均值为14.95%。

表2 饲粮能量水平对杜泊羊生长期养分利用率的影响

表3 饲粮能量水平对杜泊羊生长期气体产量的影响
2.5 绝食代谢产热量和维持代谢能的估测
绝食状态产热量(Fasting heat production,FHP)是指绝食状态下的动物产热,绝食代谢产热量还可根据沉积能(RE)与代谢能摄入量(MEI)的回归公式求得,可得如下公式:
公羊:RE = 0.758(MEI)- 328.698(R2= 0.982,P< 0.001)[1]
母羊:RE = 0.742(MEI)- 320.385(R2= 0.963,P< 0.001)[2]
由公式[1]和[2]求得,杜泊羊生长期公羊的绝食代谢产热量为328.70 kJ/KgW0.75;母羊的绝食代谢产热量为320.39 kJ/KgW0.75。代谢能用于维持的效率公羊为0.758,母羊为0.742。则公羊MEm=328.698/0.758=433.87 kJ/kgW0.75,母羊MEm=320.385/0.742=431.91 kJ/kgW0.75。
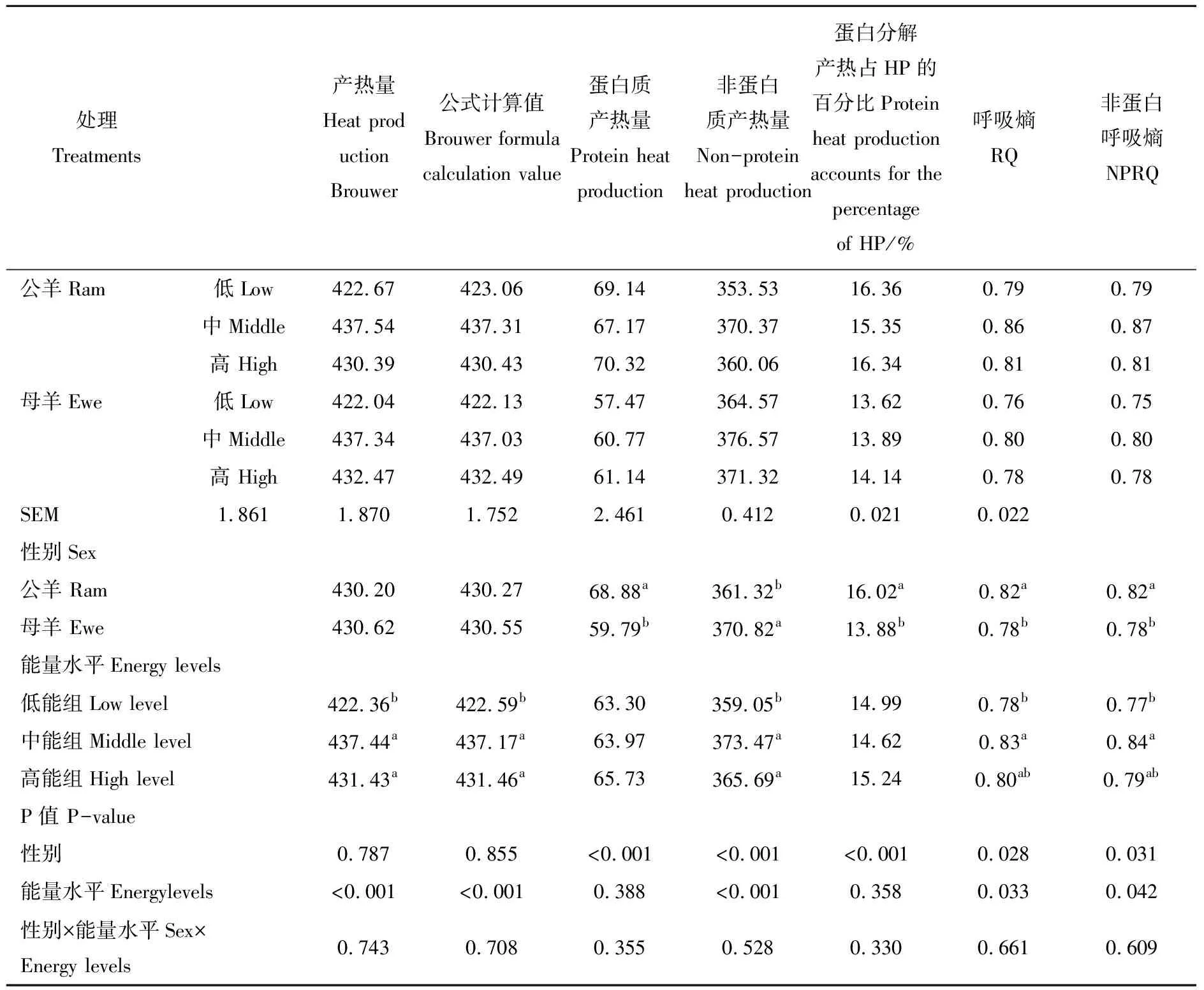
表4 不同能量水平对杜泊羊生长期产热量和呼吸熵的影响
3 讨论
3.1 能量水平对杜泊羊生长期养分利用率的影响
一般来讲,干物质的消化率是反刍动物对饲粮利用率的综合反映,本试验结果表明一定范围内提高饲粮能量水平提高了杜泊羊对干物质和有机物的利用率,提高了对中性洗涤纤维和酸性洗涤纤维的利用率,纤维物质是反刍动物能量的重要来源,本试验中能组的能量利用率的提高可能与提高了纤维物质的利用率有关。正常情况下粪能是反刍动物饲料能量中损失最大的部分,反刍动物采食精料时粪能损失为20%~30%,采食粗饲料时为40%~50%,采食低质粗饲料时可达60%,本试验中杜泊羊公羊粪能损失平均值为36.93%,母羊为38.41%,本试验采用的是粗料和精料混合饲喂,因此粪能损失符合反刍动物对能量的利用效率。反刍动物尿能的损失量比较稳定,影响尿能损失的因素主要是饲料结构,特别是饲料中蛋白质水平、氨基酸平衡状况及饲料中有害成分的含量。饲料蛋白质水平增高,氨基酸不平衡,氨基酸过量等,均可提高尿氮排泄量,增加尿能损失,降低代谢能值[12]。本试验求得杜泊羊公羊的尿能损失占总能的比例为2.90%,母羊为2.65%,略低于一般反刍动物4%~5%的水平,与金功亮等[13]求得的Sannan山羊尿能损失为2.45%,孙玉贤等[14]求得的青山羊尿能损失为2.40%接近,这可能是由于本试验采用的的粗蛋白水平较为合理,因此尿能损失较低。
本研究得出公羊生长期代谢能的维持效率为0.758,母羊为0.742。孙家义等[15]求得生长期小尾寒羊代谢能的维持效率为0.710,ARC(1980)[16]使用的均值为0.720,杨在宾等[17]报道大尾寒羊生长期代谢能的维持效率为0.792。本试验的结果与上述研究结果接近,这可能是由于反刍动物代谢能的维持效率具有相对稳定性。
3.2 能量水平对杜泊羊产热量、绝食代谢产热量及热增耗的影响
本研究采用呼吸面具间接测热法对杜泊羊的总产热进行了测定,结果显示Brouwer公式计算值与RQ法测得的结果很接近,证明本试验采用RQ法计算的结果是较为准确的。本研究结果显示中能组和高能组的产热量显著高于低能组,原因可能是能量摄入量提高后试验羊的代谢水平提高导致产热量增加。10.66 MJ/kg能量水平下单位代谢体重的产热量为437.44 kJ/kgW0.75,与Meccawi等[18]用山羊的测定结果432.00 kJ/kgW0.75相比接近;而与金功亮[13]等用Saanen奶山羊的测定结果581.30 kJ/kgW0.75相比则偏低。原因可能是由于品种差异的缘故;也可能是由于本试验期处于夏天,环境温度较高,因此产热量偏低,这也符合动物生理特点,即在动物上下临界温度范围内,如果环境温度上升则动物产热量减少,如环境温度下降则动物产热量增加[11]。本研究结果显示蛋白质产热量各组间差异不显著,这与杨在宾等[19]用青山羊泌乳母羊得到的结果一致。
Luo等[20]报道山羊FHP为298 kJ/kgW0.75,Haque等[21]报道Cheghu绒山羊FHP为326 kJ/kgW0.75,杨在宾等[22]利用青山羊空怀母羊测得FHP为260.54 kJ/kgW0.75。本试验求得杜泊羊公羊的FHP为328.70 kJ/kgW0.75,母羊的FHP为320.39 kJ/kgW0.75。本试验的结果与Haque的结果接近而比杨在宾、Luo等测得的结果偏高,ARC(1980)[16]报道绵羊的年龄增加有促使绝食代谢产热量下降的趋势。本试验的结果可能与此原因有关,本试验选取的试验羊均为断奶后不久年,处于生长期,因此绝食代谢产热量偏高。
3.3 杜泊羊生长期维持代谢能需要量的研究
维持是指动物生存过程中的一种基本状态,合理平衡维持需要与生产需要之间的关系,尽可能减少维持消耗,可提高生产效率。在动物生产潜力允许范围内,增加饲料投入,可相对降低维持需要,从而增加生产效益。维持能量需要不仅包括绝食代谢的能量,也包括随意活动的采食量以及必要的抵抗应激环境所需要的能量[12]。本试验求得杜泊羊生长期公羊维持代谢能需要量为433.87 kJ/kgW0.75,母羊维持代谢能需要量为431.91 kJ/kgW0.75。Kamalzade等[23]报道swifter绵羊MEm为340~480 kJ/kgW0.75;Al Jassim等[24]报道阿华西绵羊MEm为342~482 kJ/kgW0.75;Prieto等[25]求得生长期哥伦比亚山羊MEm为443 kJ/kgW0.75;Luo等[26]推算得安哥拉山羊单羔和多羔的MEm分别为533、442.75 kJ/kgW0.75;赵玉民[27]测得育成期的细毛羊母羊的MEm为481.18 kJ/kgW0.75;ARC(1980)[16]推荐绵羊MEm为420~450 kJ/kgW0.75。综合报道,维持代谢能需要量各报道不尽相同,可能主要是由于测定方法、品种差异、饲粮组成和外界环境的不同所致。本试验的结果符合ARC绵羊标准的推荐范围。
4 结论
4.1 公羊对于DM、OM、CP 、GE的表观消化率高于母羊,但NDF和ADF的表观消化率公母羊之间差异不显著。
4.2 本试验设置的能量梯度范围内,提高饲粮能量水平提高了杜泊羊对DM和OM的利用率;提高了GE的表观消化率和代谢率,但较高能量水平又降低了GE的表观消化率和代谢率;能量水平没有影响杜泊羊对CP的利用率。
4.3 单位代谢体重的产热量公母羊之间差异并不显著,蛋白质产热量公羊极显著高于母羊,非蛋白质产热量母羊极显著高于公羊。
4.4 间接测热试验方法求得杜泊羊生长期公羊维持代谢能需要量为433.87 kJ/kgW0.75,母羊为431.91 kJ/kgW0.75。
[1] 冯仰廉,MOLLISON,SMITH J,等.新闭路循环式面具呼吸测热法的研究[J].北京农业大学学报,1985,1:17-27
[2] 赵有璋.杜泊羊及其在我国初步利用效果[J].现代畜牧兽医,2011(1) :21-23
[3] MIRON,ADIN G,SOLOMON R,etal.Effects of feeding cows in early lactation with soy hulls as partial forage replacement on heat production,retained energy and performance[J].Animal Feed Science and Technology,2010,155:9-17
[4] El-MECCAWI S,KAM M,BROSH A,etal.Energy intake,heat production and energy and nitrogen balances of sheep and goats fed wheat straw as a sole diet[J].Livestock Science,2009,125:88 -91
[5] 杨诗兴,彭大惠,张文远,等.湖羊能量与蛋白质需要量研究[J].中国农业科学,1988,2:73-81
[6] NRC.Nutrient Requirements of small ruminants:sheep,goats,cervids,and new world camelids[S].Washington,D.C.:National Academy Press,2007
[7] 贺建华.饲料分析与检测[M].北京:中国农业出版社,2008
[8] BLAXTER K L,MCC GRAHAM N,ROOK J A F,etal.Apparatus for the determination of the energy exchange of calves and of sheep[J]. The Journal of Agricultural Science,1953,45:101
[9] BLAXTER KL,CLAPPERTON J L.Prediction of the amount of methane produced by ruminants[J].British Jounal of Nutrition,1965(19):511
[10] BROUWER E.Report of the sub-committee on constants and facors[C].In:Blaxter,K L.ed Energy Metabolism of Farm Animals.London: Academic Press,11:441-443
[11] 陈 杰.家畜生理学[M].北京:中国农业出版社,2003
[12] 杨 凤.动物营养学[M].北京:中国农业出版社,2008
[13] 金功亮,赵路明,李青旺,等.西农莎能奶山羊泌乳期能量消化率和代谢率的测定[J].畜牧兽医杂志,1983,(2):18-27
[14] 孙玉贤,方国玺,李凤双.生长青山羊能量需要的研究[J].山东农业大学学报,1987,(1):9-19
[15] 孙家义.小尾寒羊生长期能量代谢规律的研究[D].泰安:山东农业大学,1993
[16] Agricultural Research Council.The Nutrient Requirements of Ruminant Livestock[S]. Commonwealth Agricultural Bureaux,Farnham Royal,England,1980
[17] 杨在宾,杨维仁,张崇玉,等.大尾寒羊生长期能量需要量及代谢规律研究[J].山东农业大学学报,1999,(2):97-103
[18] El-MECCAWI S,KAM M,BROSH A,etal.Heat production and energy balance of sheep and goats fed sole diets of Acacia saligna and Medicago sativa[J].Small Ruminant Research,2008,75(2-3):199-203
[19] 杨在宾,杨维仁,李凤双.青山羊空怀及泌乳母羊的绝食代谢研究[J].中国养羊,1991,4: 17-19
[20] LUO J,GOESTCH A L,NSAHLAI I V,etal.Maintenance energy requirements of goats:predictions based on observations of heat and recovered energy[J].Small Ruminant Research,2004,53: 221-230
[21] HAQUE,N.MURARILAL,M Y.KHAN,M Y,etal.Metabolizable energy requirements for maintenance of pashmina producing Cheghu goats[J].Small Ruminant Research,1998,27(1):41-45
[22] 杨在宾,李凤双,杨维仁.青山羊泌乳期母羊的能量需要量研究[J].畜牧兽医学报,1993,24(5):391-398
[23] KAMALZADEH A.Energy and Nitrogen Metabolism in Lambs during Feed Restriction and Realimentation[J].Journal of Agricultural Science and Technology,2004(6):21-30
[24] Al JASSIM R A M,S A HASSAN,A N Al-ANI,etal.Metabolizable energy requirements for maintenance and growth of Awassi lambs[J].Small Ruminant Research,1996(20):239-245
[25] PRIETO C,AGUILERA J F,LAEA L,etal.Protein and energy requirements for maintenance of indigenous Granadina goats[J].The British Journal of Nutrition,1990,63(2):155-163
[26] J LUO,A L GOESTCH,I V NSAHLAI,etal.Prediction of metabolizable energy and protein requirements for maintenance,gain and fiber growth of Angora goats[J].Small Ruminant Research,2004(53):339-356
[27] 赵玉民.中国美利奴羊育成母羊能量和蛋白质需要量研究[D].延边:延边大学,2005
