2009/2010—2011/2012年度夏秋季南奥克尼群岛水域南极磷虾捕捞群体年龄结构时空变化
2014-03-08朱国平朱小艳李莹春徐怡瑛夏辉徐鹏翔许柳雄
朱国平 朱小艳 李莹春 徐怡瑛 夏辉 徐鹏翔 许柳雄
(1上海海洋大学海洋科学学院,上海201306;2国家远洋渔业工程技术研究中心,上海210306;3大洋渔业资源可持续开发省部共建教育部重点实验室,上海海洋大学,上海201306;4农业部大洋渔业资源环境科学观测实验站,上海海洋大学,上海201306)
0 引言
作为目前全球单一物种生物量最大的甲壳类浮游动物,南极大磷虾(通常称南极磷虾,Euphausia superba Dana,以下简称磷虾)在南大洋海洋生态系统中起着“关键种”的作用,其在生态和商业性渔业上均具有极为重要的地位[1]。了解磷虾的种群结构是相关渔业管理的主要事务之一,而确定不同年龄组的分布对于研究其种群结构非常关键[2],确定捕捞群体年龄组成也是开展渔区磷虾种群结构研究的基础[3]。尽管目前磷虾鉴龄工作尚需要进一步完善,但关于磷虾生长方面的研究已为基于体长频度法估算南极磷虾年龄结构提供了足够的基础[4-8],种群中捕捞群体的年龄结构对于确定资源性质及渔业对策有着非常重要的意义[9]。
自20世纪20年代初“发现号”调查以来,作为磷虾环南极分布的关键区域之一,斯科舍海一直起着非常重要的作用[4,10]。1981年,首次国际南极海洋系统和储量的生物调查试验(FIBEX)对斯科舍海的磷虾生物量进行了调查,并对该资源的大尺度结构进行了描述[11]。该调查显示,斯科舍海磷虾体长组分布并不统一,相对于仔虾而言,成体虾的分布水域更靠北[12]。此后,一系列的标准化中尺度长期调查,便开始对磷虾种群动力学的年际变化进行研究[13],结果表明磷虾的补充变化较大[14-15]。在这一系列调查中,南奥克尼群岛一直为重点调查区域。首先,南奥克尼群岛为精细管理单元(SSMU)概念的关键区域之一[16],该区内磷虾起源和个体输送问题是磷虾资源管理中需要解决的重要事宜,而这些问题的澄清对于以SSMU分配预防性磷虾渔获量的资源管理方式以及完善磷虾资源评估而言非常重要。其次,近年来,磷虾渔业从南设得兰群岛向东转移至南奥克尼群岛[17]。第三,考虑到陆基捕食者的丰度,16个SSMU中的3个集中于南奥克尼群岛水域[17],而该水域磷虾的生物量尚未得到系统的研究[18]。鉴于此,本研究以南奥克尼群岛为研究区域,以精细研究单元为手段,分析2009/2010—2011/2012渔季该水域磷虾的种群组成时空变化,从而为理解磷虾的种群动态提供科学信息,并为该水域的渔场形成机制提供基础数据。
1 材料与方法
1.1 调查海域和时间
调查时间和调查船见表1。调查水域主要分布于南极半岛东北部——南奥克尼群岛水域(图1)。取样工具均为商业性中层磷虾拖网,系四片式单船中层拖网(网囊网目尺寸均为2.0 cm)。本次调查为渔业调查,未设具体站点。作业时,按照南极海洋生物资源养护委员会(CCAMLR)要求,记录起放网时间和位置、开始(结束)作业时间和位置和拖曳深度等参数。
每站点随机取样本100或200尾,共计26 100尾,采用 Lockyer[19]的方法测量磷虾的标准体长(SL)[20]。

表1 调查相关数据Table 1.Summary on survey data
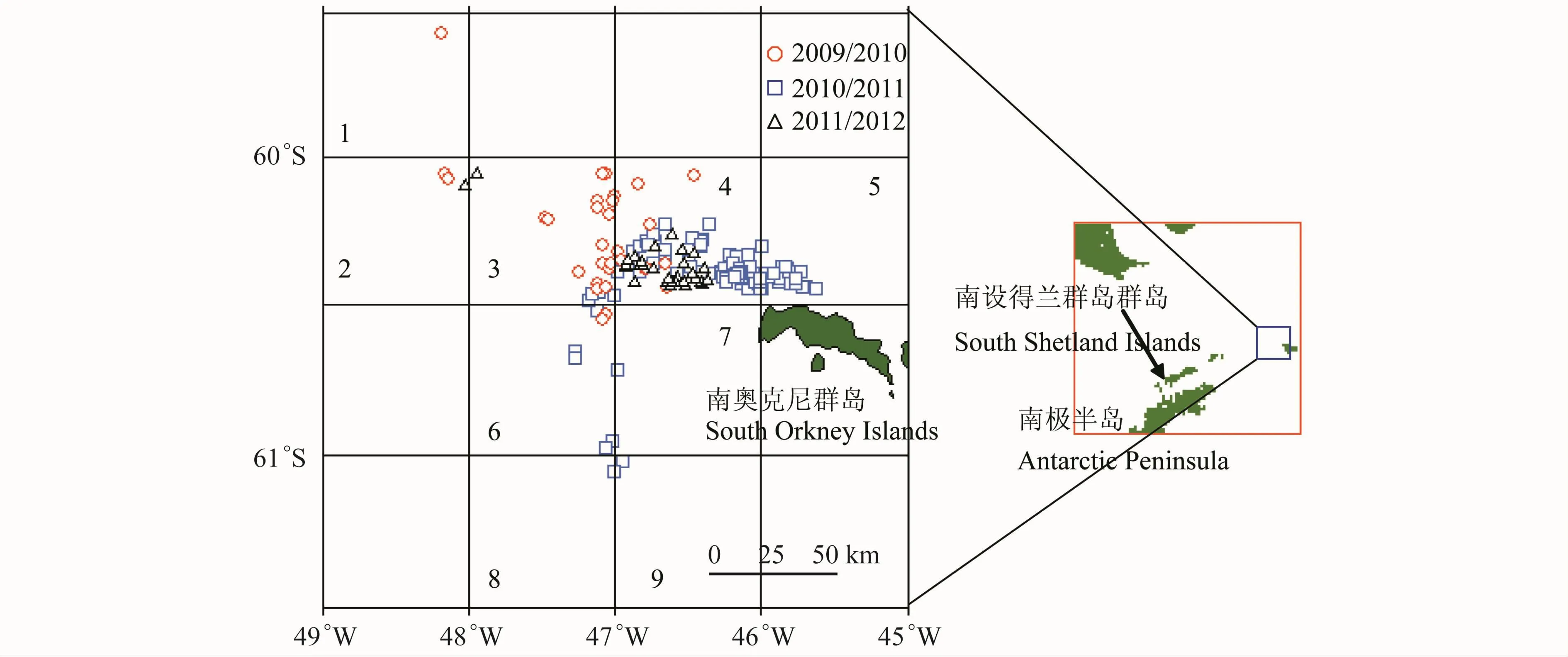
图1 调查站位及单元划分Fig.1.Survey locations and unit division
1.2 数据处理
(1)划分单元
为了更精确地了解南奥克尼群岛水域磷虾资源的种群组成变化,将研究区域依经纬度60′×30′划分成16个单元,但仅其中9个单元有调查站位,分别以数字1—9表示各单元(图1)。
(2)确定体长组距
确定体长分组组距是开展体长频率分析的必要步骤[21],不适当的组距划分可能会给估算结果带来较大的不确定性。通常,用于确定体长分组组距的方法有两种,分别为 Snedecor法[22]及 Sturges法[23],如下式(1)和(2):
组距(Snedecor)=Range/(Range/SD×4)(1)
组距(Sturges)=Range/(1+3.322×lg N)(2)其中,Range表示体长全距,即最大值与最小值之差;SD为体长的标准差;N表示样本量。Snedecor法估算的体长分组组距通常偏小,而Sturges法估算的结果偏大[21],故本研究综合两种方法,依具体情况选取较为合适的体长分组组距。
(3)确定年龄组
为了了解各单元磷虾种群年龄结构随时间的变化,本研究采用FiSAT II软件[24]计算不同单元各旬磷虾的年龄组。首先利用Bhattacharya法估算年龄,再利用最大似然法(NORMSEP)[25-27]改善 Bhattacharya法获得的平均体长、种群规模和标准差数据,具体可参考朱国平等[8]的方法。
(4)分析不同时间尺度(旬别和月份)及空间尺度(单元)磷虾体长百分比组成的相似性:利用百分比相似性指数(PSI)估算比例的相似性[28]。具体计算公式如下:
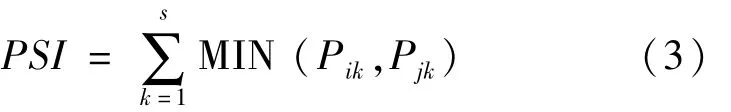
其中,PSI为各项目两两组别间体长组成百分比的相似性,Pik为第i个组别中体长组k所占的比例,Pjk为第j个组别中体长组k所占的比例,s为体长组个数。通常认为,当PSI值≥80时,即认为两者之间相似[29]。以S表示上旬,Z表示中旬,X表示下旬。
以上体长均若为平均值,则利用Bootstrap法估算该平均值的95%置信区间,每组数据进行重抽样(resampling),并迭代1 000次,以降低观察次数不足带来的不确定性[1]。通过变异系数(CV)反映各旬别中平均体长的波动情况。利用非参数多元维度分析(NMDS)划分各年份不同旬别磷虾体长组的类别。利用聚类分析了解各单元磷虾年龄组成分布情况,并采用Bray-Curtis相似性划分组别。利用双样本Kruskal Wallis检验(针对两个体长组,K-W检验)或多个独立样本Kolmogorov-Smirnov检验(针对3个或以上体长组,K-S检验)分析体长组成是否存在显著性差异(p=0.05)。利用单因子方差分析(ANOVA)或t检验分析不同时间尺度(年度及旬别)平均体长是否存在显著性差异(p=0.05)。Bootstrap计算在Resampling Stat in EXCEL程序中进行,其他统计检验则在SPSS 17.0中运算。
2 结果
2.1 组距确定及年度体长分布
通过Snedecor法估算的体长分组组距为1.5 mm,而 Sturges法估算的组距则为3.2 mm,取平均值及考虑分析简便后,本研究将体长分组组距确定为2 mm。
2009/2010年度,共取样2 200尾,磷虾体长范围为33.2—69.0 mm,平均体长为51.8 mm(95%置信区间为 51.6—52.1 mm,下同),优势体长组为52—60 mm(52.3%),另在44—48 mm有一个次峰区;2010/2011年度,体长范围为22.9—59.3mm(n=18 500),平均体长为 44.5 mm(44.4—44.6 mm),优势体长组为42—50 mm(56.8%);2011/2012年度,体长范围为 18.5—59.4 mm(n=5 400),平均体长为41.7 mm(41.5—41.8 mm),优势体长组为 36—46 mm(67.3%)(图2)。总的来讲,三个年度内,磷虾体长的变化幅度相差不大(CV=0.117 0—0.125 2),但体长频次分布并不具相似性(PSI=41.25—73.42)。由图2可知,2010/2011年度和 2011/2012年度,体长频次分布基本上呈单峰分布,而2009/2010年度却呈双峰分布,且三个年度体长频次分布(ANOVA,χ2=3 678.258,df=2,p<0.001)及平均体长(F=2 713.812,p<0.001)均存在着显著性差异。

图2 2009/2010—2011/2012年度,夏秋季南奥克尼群岛磷虾体长分布Fig.2.The distribution of standard length for Antarctic krill in the South Orkney Islands during the austral summer-fall from 2009/2010 to 2011/2012 fishing season
2.2 各旬体长分布
2009/2010年度,调查时间为1月下旬至2月中旬,此期间,平均体长虽呈增长趋势,从1月下旬的51.2 mm增加到52.5 mm,但各旬间仍不存在显著性差异(ANOVA,F=2.435,p=0.088>0.05),其中2月上旬的体长变化较大(CV=0.1211);2010/2011年度,2月上旬(SL=40.1 mm)至中旬(SL=47.6 mm)的平均体长趋势同2009/2010年度,此后平均体长逐步呈下降趋势,至3月中旬(SL=43.4 mm)为调查期间次低,之后平均体长又逐步增加,至4月上旬后呈小幅下降,其中2月上旬的体长变化最大(CV=0.147 1),而4月上旬的体长变化较小(CV=0.071 4),且各旬间平均体长具显著性差异(ANOVA,F=893.253,p<0.001);2011/2012年度,平均体长的变化趋势则与2010/2011年度同期相反,两个旬别的体长变化较为相似,但平均体长存在显著性差异(t检验,F=20.113,p<0.001)(图3和图4)。
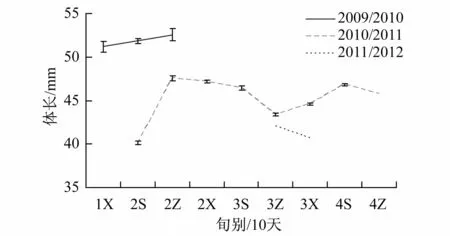
图3 2009/2010—2011/2012年度夏秋季南奥克尼群岛磷虾分旬别平均体长分布Fig.3. The distribution of average standard length by 10-day in a month for Antarctic krill in the South Orkney Islands during the austral summer-fall from 2009/2010 to 2011/2012 fishing season
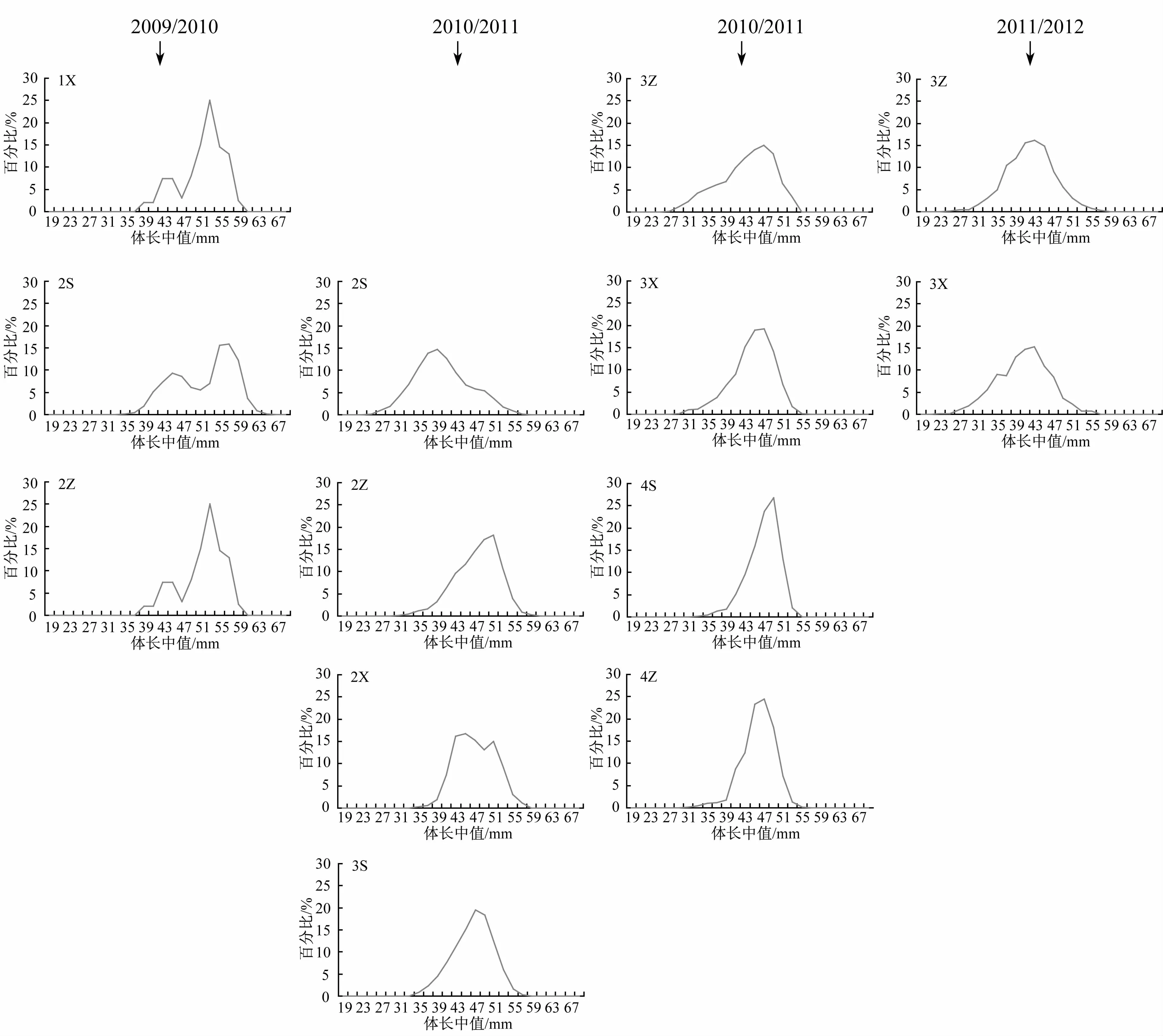
图4 2009/2010—2011/2012年度夏秋季南奥克尼群岛南极磷虾分旬别体长分布Fig.4.The distribution of standard length by 10-day in amonth for Antarctic krill in the South Orkney Islands during the austral summer-fall from 2009/2010 to 2011/2012 fishing season
2009/2010年度,各旬间体长组成存在显著性差异(K-S检验,χ2=10.230,p=0.006<0.05),但2月上旬与中旬体长频次分布具相似性(PSI=84.6),且体长组成也不存在差异(K-W检验,Z=1.282,p=0.075>0.05);2010/2011年度,28个旬别组中,仅4组旬别组体长频次分布具相似性(PSI=84.7—86.4),而2月上旬与4月中旬体长频次分布相似性最低(PSI=40.1),各旬间体长组成存在显著性差异(χ2=4 178.523,p<0.001);2011/2012年度,两个旬别间的体长频次分布较为相似(PSI=88.2),但体长组成却存在着显著性差异(Z=3.610,p<0.001)(图4)。
聚类分析表明,2009/2010—2011/2012年度各旬别磷虾体长组可分成3个组,其中2011年2月上旬(20112S)和2012年3月中旬(20123Z)可划分为一组,2010年1月下旬至2月中旬以及2011年2月中旬可归为一组,这几个体长组均呈双峰分布,其它的体长组划归为一组(图5,stress=0.05)。

图5 2009/2010—2011/2012年度,夏秋季南奥克尼群岛磷虾分旬别体长组聚类分析Fig.5.Clustering analysis on standard length classes by 10-day in amonth for Antarctic krill in the South Orkney Islands during the austral summer-fall from 2009/2010 to 2011/2012 fishing season
2.3 年龄结构
从不同年度同旬别上来看,2009/2010年度2月上旬以4+和6+虾为主,而2010/2011年度同期则以1+、2+和4+为主,有一部分幼虾补充到群体中;进入中旬,两个年度的磷虾均以产卵群体为主,但2009/2010年度多为3+龄虾,而2010/2011年度则以4+和5+龄为主;2010/2011年度,3月中旬,磷虾种群组成以4+和5+龄为主,而2011/2012年度同期则多为2+、3+和4+龄虾;进入3月下旬,两个年度的磷虾种群中均为4+龄占优,说明此时南奥克尼群岛水域出现的虾群多为产卵群体(表2)。

表2 2009/2010—2011/2012年度,夏秋季各旬别分单元南奥克尼群岛磷虾种群组成Table 2.Population composition of Antarctic krill in the South Orkney Islands during the austral summer-fall from 2009/2010 to 2011/2012 fishing season by 10-day and unit
从空间角度上来看,值得注意的是,1+龄虾(2010/2011年2月上旬和中旬以及2011/2012年3月中旬和下旬)均出现在第4单元,即离南奥克尼群岛最近的水域,而2+虾也多出现于离南奥克尼群岛较近的第4单元和第5单元。离南奥克尼群岛较远的单元中则多为4+龄以上的虾群(如,第1—3单元)。综合所有渔季的磷虾样本,针对其年龄组成的聚类分析可以看出,第2、7、8和9单元可划分为一组(Bray-Curtis相似性为74.42%),而第3—6单元可划为一组(Bray-Curtis相似性为45.43%),第1单元与其他两组间相似性较低(Bray-Curtis相似性仅为24.47)(图6)。

图6 2009/2010—2011/2012年度,夏秋季各单元南奥克尼群岛磷虾年龄组成聚类分析Fig.6.Cluster analysis on age composition of Antarctic krill population in the South Orkney Islands during the austral summer-fall from 2009/2010 to 2011/2012 fishing season
3 讨论
3.1 种群组成
磷虾的生命周期约在 3—8龄之间[4-5,7,30]。目前,研究人员多将体长25—30 mm的幼虾视为1+龄,2+龄虾的体长范围在40—45 mm,3+—5+龄虾的平均体长分别为50 mm,55 mm和58 mm,但这些估算结果仍受到区域性的水文气象因素、性别及性成熟度所影响[4,7]。由此推算,从年度上来看,本研究中2009/2010年度有两个体长组簇,一个为52—60 mm(4+—6+龄),另一个为44—48 mm(2+龄);2010/2011年度和 2011/2012年度仅有一个体长组簇,分别位于36—46 mm(2+龄)和42—50 mm(2+—3+龄)。基于表1的数据可知,2009/2010年度2月份主要以4+龄和6+龄的成虾为主,2010/2011年度3月份主要以2+—4+龄为主,若南奥克尼群岛水域磷虾为本地种,则2010/2011年度2月份该水域中的1+龄虾为主,2011/2012年度3月份该水域中的优势种群应为3+—5+龄。但事实上,2010/2011年度,该水域占优势的种群为2+龄和4+龄,2011/2012年度3月份的优势年龄结构为4+龄,两个年度推算结果与实际情况有所偏差。因此,可以认为南奥克尼群岛水域的磷虾应该是由其他产卵场随海流输送而来。基于2000年1—2月份南极西南大西洋水域磷虾种群的分析,Siegel等[13]表示该种群中共有3个体长组组成,第1簇主要由35 mm以下的磷虾组成,呈单峰(众数体长为26 mm)分布,其中绝大部分(79%)为幼体,并有少量未成体,这表明南奥克尼群岛水域磷虾的补充群体(1+龄虾)数量非常低(4%),且仅出现在CCAMLR 48.2亚区的东侧,45°W以西水域则完全未见到1+龄虾;第2簇主要由30—40 mm的磷虾个体(众数体长为48 mm)组成,其中55%为未成体,而39%为成体,主要为4+龄虾,而体长在30—45 mm的磷虾则多为2+—3+龄虾,但数量相对较少;另外一个体长组簇主要由大个体磷虾(82%,众数体长为52 mm)组成,这些磷虾约4+—5+龄。同时,Siegel等[13]还表示,体长较大的磷虾(众数体长为52 mm,年龄4+—6+龄)主要与南极绕极流(ACC)水团有关,而个体较小的磷虾(众数体长为26 mm,1+龄)则生活在维德尔海水域。Sologub和 Rameslo[2]也表示,2009/2010年度1—3月份南奥克尼群岛磷虾体长范围在 29—61 mm之间,呈双峰分布,优势体长组分别为43—47 mm和 53—55 mm。本研究中,2009/2010年度1—2月份磷虾体长范围为33.2—69.0 mm,平均体长为51.8 mm,优势体长组为52—60 mm(52.3%),另在44—48 mm有一个次峰区,这点与Sologub和Rameslo[2]的结果较为相似,但在年龄界定方面仍有所差异。
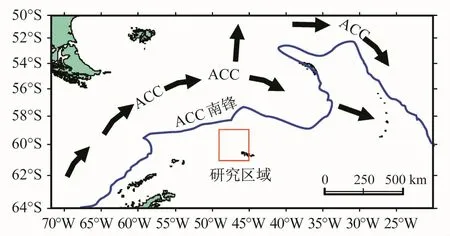
图7 南极绕极流(ACC)及其南锋流向示意图(参考Lee等[33])Fig.7.The sketch map of Antarctic Circumpolar Current(ACC)and Southern ACC Front(Reproduced from Lee et al.[33])
通常认为,较大体长组的磷虾(3+、4+和5+龄)主要分布于 ACC水团中[2,31-32],该水团从德雷克海峡沿斯科舍海南侧向东流,并延伸至南奥克尼群岛北部水域[33](图7),中等尺度(30—40 mm)的体长组则出现在南奥克尼群岛南部,并在维德尔海水团流出处将大型成体分布隔离开来,而较小体长组的磷虾则被输送到南桑德维奇群岛西北水域,并进入南乔治亚岛水域[13]。这也正好印证了本研究的结论,即处于南奥克尼群岛北侧的第1单元中多为5+龄以上的磷虾,而南奥克尼群岛南侧较远的第7—9单元中则以4+龄以下的个体占优,第2单元之所以被划归到与第7—9单元同组,一个可能的原因是第2单元的样本数量相对较小(n=157),导致年龄组成上出现一定的偏差。由此亦可进一步看出,高龄虾的分布偏向于外海无冰区,而低龄虾偏向冰区和冰缘区,一些学者在南极半岛地区和普里兹湾均发现过这种现象[34-35]。
本研究中,不同年度磷虾体长及年龄组成存在着较大的差异,这在很大程度上可能是由海况和食物状况的大尺度变化所造成。Sologub和Remeslo[2]表示,南奥克尼群岛不同年度磷虾体长-年龄组成及成熟状况的差异可由食物及温度状况年际变化解释,而这两者之间也存在着较好的内在联系。
3.2 1+龄虾的补充
根据本研究的结果可知,三个调查航次中,尽管调查时间由1月下旬持续到4月中旬,但1+龄世代的磷虾个体非常少,有些年份甚至未出现1+龄个体的磷虾(如2009/2010年)(图8)。Sologub和Rameslo[2]发现2009年和2010年1—3月份南奥克尼群岛水域并未发现1+龄出现。Siegel等[13]也曾表示,45°W以西未发现有稚虾(26 mm以下个体),该体长组主要出现于南奥克尼群岛以东水域,并向北延伸至南乔治亚岛南部。早期一些学者也得出了类似的结果,即不足30 mm的磷虾个体(1+龄世代)很少出现在 CCAMLR 48.2小区[36]。

图8 2009/2010—2011/2012年度夏秋季各旬 30mm以下(1+龄)磷虾个体所占比例Fig.8.The percentage of standard length less than 30 mm(age 1+)by 10-day in a month during the austral summerfall from 2009/2010 to 2011/2012 fishing season
至于南奥克尼群岛水域较少出现1+龄虾的原因可能有以下几个方面。第一,商业性拖网选择性。通常商业性磷虾拖网网目尺寸较科学调查用拖网(如,IKMT、RMT8及 bongo net等)大,因此不排除部分幼虾从网囊中逃逸[37]。第二,海况或食物来源条件所致。一些研究表明,南极半岛和南乔治亚岛水域1+龄虾的出现概率较高,甚至出现1+龄虾的猬集[14-15,38-41],而南奥克尼群岛水域缺少1+龄世代的磷虾也显示前一年输送源磷虾产卵成功率较低,这可能是受到维德尔海不佳的食物来源及海况因素(较低的水温)致使维德尔海磷虾仔体和仔体后期生长较慢所影响[42]。
需要指出的是,本研究采用体长频次法估算了南极磷虾的种群年龄。尽管该方法曾多次在南极磷虾种群结构分析中得到了应用[2-3,8-9,13],但考虑到南极磷虾特殊的生长特性(如,负生长[43]及蜕壳[44]等)以及体长频次分析法本身所存在的一些缺陷(如,大龄个体体长分布重叠较为严重[1,45])等,相关结论仍需要进一步地验证。目前,一些学者采用各种手段,如实验室暂养[46]、脂褐素含量分析[47-48]等估算南极磷虾的年龄,相关研究所提供的信息亦可为体长频次法估算年龄提供较好的验证和支撑。
致谢 感谢上海开创远洋渔业有限公司及“开欣”轮、“开顺”轮和大连远洋渔业有限公司“安兴海”轮、“连兴海”轮船长和船员在取样过程中给予的大力配合。感谢中国南极磷虾渔业科学观察员在海上数据收集过程中所付出的辛勤劳动。感谢美国Old Dominion University海洋、大气与地球科学系Molly Mikan女士给予英文摘要润色。本研究得到了农业部南极海洋生物资源开发利用项目及上海海洋大学骆兆尧大学生科技创新基金项目(A-0204-13-001912)的部分支持。
1 朱国平.南极磷虾种群生物学研究进展Ⅰ-年龄、生长与死亡.水生生物学报,2011,35(5):862—868.
2 Sologub D O,Remeslo A V.Distribution and size-age composition of Antarctic krill(Euphausia superba)in the South Orkney Islands region(CCAMLR Subarea 48.2).Science Journal Paper:CCAMLR Science,2011,18:123—134.
3 左涛,赵宪勇,黄洪亮,等.南极半岛邻近水域南极大磷虾商业捕捞群体的年龄结构时空变化.渔业科学进展,2012,33(4):1—10.
4 Marr JW S.The Natural History and Geography of the Antarctic Krill(Euphausia superba Dana).Cambridge:University Press,1962:33—464.
5 Aseev Y P.Size structure of krill populations and life span in the Indian Ocean sector of the Antarctic.Hydrobiology Journal,1984,6:89—94.
6 Siegel V.Age and growth of Antarctic Euphausiacea(Crustacea)under natural conditions.Marine Biology,1987,96(4):483—495.
7 Candy SG,Kawaguchi S.Modelling growth of Antarctic krill.II.Novel approach to describing the growth trajectory.Marine Ecology Progress Series,2006,306:17—30.
8 朱国平,吴强,冯春雷,等.基于渔业调查的南极半岛北部水域南极磷虾种群年龄结构分析.生态学报,2011,31(16):4620—4627.
9 王荣,鲁北伟,李超伦,等.南极磷虾年龄组成的体长频数分布混合分析.海洋与湖沼,1995,26(6):598—605.
10 Mackintosh N A.Distribution of post-larval krill in the Antarctic.Discovery Reports,1973,36:95—156.
11 El-Sayed S Z.History,organization and accomplishmentsof the BIOMASSprogramme//El-Sayed SZ.Southern Ocean Ecology.The BIOMASSPerspective.Cambridge:Cambridge University Press,1994:1—8.
12 BIOMASS.Non-acoustic Krill Data AnalysisWorkshop,Cambridge.BIOMASSReport Series,1991,66:1—54.
13 Siegel V,Kawaguchi S,Ward P,etal.Krill demography and large-scale distribution in the southwest Atlantic during January/February 2000.Deep Sea Research PartⅡ:Topical Studies in Oceanography,2004,51(12-13):1253—1273.
14 Siegel V,Loeb V.Recruitment of Antarctic krill Euphausia superba and possible causes for its variability.Marine Ecology Progress Series,1995,123:45—56.
15 Watkins J.A composite recruitment index to describe interannual changes in the population structure of Antarctic krill at south Georgia.CCAMLR Science,1999,6:71—84.
16 Hewitt R P,Watters G,Trathan P N,et al.Options for allocating the precautionary catch limit of krill among small-scalemanagement units in the Scotia Sea.CCAMLR Science,2004,11:81—97.
17 Kawaguchi S,Nicol S,Taki K,et al.Fishing ground selection in the Antarctic krill fishery:trends in patterns across years,seasons and nations.CCAMLR Science,2006,13:117—141.
18 Brierley A S,Watkins JL.Acoustic targets at South Georgia and the South Orkney Islands during a season of krill scarcity.Marine Ecology Progress Series,1996,138:51—61.
19 Lockyer C.Wetweight,volume and length correlation in the Antarctic krill,Euphausia superba.Discovery Reports,1973,36:152—155.
20 CCAMLR.Scheme of international scientific observation.Hobart,Australia:CCAMLR.2011.http://www.ccamlr.org/en/system/files/obsman.pdf.
21 陈国宝,李永振,陈丕茂,等.鱼类最佳体长频率分析组距研究.中国水产科学,2008,15(4):659—666.
22 Snedecor GW,Cochran W G.Statistical Methods.Ames,Iowa:Iowa State University Press,1967:593.
23 Sturges P.Suverys and Sampling//Fife-Schaw C,Smith JA,Hammond S,et al.Research Methods in Psychology.London:SAGE,2006:552.
24 Gayanilo FC,Sparre P,Pauly D.FAO-ICLARM stock assessment toolsⅡ (FISATⅡ).Revised version.User’s guide.FAO Computerized Information Series(Fisheries)No 8.Rome,2005:168.
25 Hasselblad V.Estimation of parameters for amixture of normal distributions.Technometrics,1966,8(3):431—444.
26 Abramson N J.Computer Programs for Fish Stock Assessment.FAO Fisheries Technical Paper,1971:101.
27 Gayanilo FC,Sparre P,Pauly D.FAO-ICLARM stock assessment tools.(FiSAT):referencemanual.FAO Computerized Information Series(Fisheries).NO.8,Rome:FAO,1997:262.
28 Whittaker R H.Communities and Ecosystems.New York:Macmillan,1975:386.
29 Newman R M.Comparison of encountermodel predictionswith observed size-selectivity by stream trout.Journal of the North American Benthological Society,1987,6(1):56—64.
30 Ikeda T.Development of the larvae of the Antarctic krill(Euphausia superba Dana)observed in the laboratory.Journalof ExperimentalMarine Biology and Ecology,1984,75(2):107—117.
31 Ichii T,Naganobu M.Surface water circulation in krill fishing areas near the south Shetland islands.CCAMLR Science,1996,3:125—136.
32 Hofmann E E,Klinck JM,Locarnini R A,et al.Krill transport in the Scotia Sea and environs.Antarctic Science,1998,10(4):406—415.
33 Lee J I,Yoon H I,Yoo K C,etal.Late Quaternary glacial-interglacial variations in sediment supply in the southern Drake Passage.Quaternary Research,2012,78(1):119—129.
34 Wang R,Chen SH.Population structure of Antarctic krill(Euphausia superba Dana)in the waters north and westof Antarctic Peninsula.Chinese Journal of Oceanology and Limnology,1990,8(2):101—108.
35 王荣,仲学锋,孙松,等.普里兹湾邻近海域大磷虾的种群结构研究.极地研究,1993,5(4):1—11.
36 Jackowski E.Distribution and size of Antarctic krill(Euphausia superba Dana)in Polish commercial catches taken in the Atlantic sector of the Southern Ocean from 1997 to 1999.CCAMLR Science,2002,9:83—105.
37 朱国平,冯春雷,吴强,等.南极磷虾调查CPUE指数变动的影响因素初步分析.海洋渔业,2010,32(4):368—373.
38 Siegel V.A concept of seasonal variation of krill(Euphausia superba)distribution and abundance west of the Antarctic Peninsula//Sahrhage D.Antarctic Ocean and Resources Variability.Berlin:Springer-Verlag,1988:219—230.
39 Loeb V,Siegel V.Status of the krill stock around Elephant Island in 1991/1992 and 1992/1993.CCAMLR Science,1994,1:7—33.
40 Kawaguchi S,Ichii T,Naganobu M.Catch per unit effort and proportional recruitment indices from Japanese krill fishery data in Subarea 48.1.CCAMLR Science,1997,4:47—63.
41 Quetin L B,Ross RM.Episodic recruitment in Antarctic krill Euphausia superba in the Palmer LTER study region.Marine Ecology Progress Series,2003,259:185—200.
42 Melnikov IA,Spiridonov V A.Antarctic krill under perennial sea ice in the western Weddell Sea.Antarctic Science,1996,8(4):323—329.
43 Nicol S,Stolp M,Cochran T,et al.Growth and shrinkage of Antarctic krill Euphausia superba from the Indian Ocean sector of the Southern Ocean during summer.Marine Ecology Progress Series,1992,89:175—181.
44 Buchholz F.Moult cycle and growth of Antarctic krill Euphausia superba in the laboratory.Marine Ecology Progress Series,1991,69:217—229.
45 Siegel V,Nicol S.Population parameters//Everson I.Krill:Biology,Ecology and Fisheries.Oxford:Blackwell Science Ltd.,2000:103—149.
46 Nicol S.Understanding krill growth and aging:the contribution of experimental studies.Canadian Journal of Fisheriesand Aquatic Sciences,2000,57(S3):168—177.
47 Harvey H R,Ju SJ,Son SK,etal.The biochemical estimation of age in Euphausiids:laboratory calibration and field comparisons.Deep Sea Research PartⅡ:Topical Studies in Oceanography,2010,57(7-8):663—671.
48 McGaffin A F,Nicol S,Virtue P,etal.Validation and quantification ofextractable age pigments for determining the age of Antarctic krill(Euphausia superba).Marine Biology,2011,158(8):1743—1755.
