重症监护室革兰阴性杆菌分布及耐药性分析*
2014-03-08高玉红李丰良
高玉红,李丰良
(云南省第一人民医院检验科,昆明650034)
革兰阴性(G-)杆菌是医院感染的重要病原菌,也是重症监护室(ICU)的重要致病菌。近年来随着抗菌药物的广泛应用,非发酵菌在G-杆菌中所占比例越来越高。为了解ICU G-的感染及耐药趋势,本文对本院2010~2012年ICU分离到的636株G-杆菌的分布及耐药性进行分析,为临床用药提供帮助。
1 材料与方法
1.1 材料 收集2010年1月到2012年12月本院ICU患者各类送检样本中分离到的636株的G-杆菌,每位患者只统计首次分离株。
1.2 方法
1.2.1 细菌培养、鉴定及药敏 将标本接种于生物梅里埃公司的哥伦比亚血平板、麦康凯平板和巧克力平板上,分别在35℃孵箱和7%CO235℃孵箱孵育24h后选取阳性菌落用梅里埃VITEK2自动鉴定仪及相应的鉴定卡及药敏卡进行鉴定及药敏实验
1.2.2 超广谱β-内酰胺酶(ESBLs)表型确证实验对VITEK2提示为产ESBLs表型的菌株,按CLSI 2010要求用双纸片扩散法确证[1]。方法为在涂有菌液的MH平板上分别贴头孢他啶和头孢他啶/克拉维酸(30μg/10μg),头孢噻肟和头孢噻肟/克拉维酸(30μg/10μg),二者中任一抑菌环差值大于或等于5mm时判断为产ESBLs。药敏纸片为英国Oxoid产品。质控菌株为大肠埃希菌ATCC25922和ATCC35218,铜绿假单胞菌ATCC27853。
1.3 统计学处理 采用WHONET 5.6软件对阳性样本进行耐药分析,SPSS 13.0软件进行χ2检验,以P<0.05为差异有统计学意义。
2 结 果
2.1 G-杆菌构成及标本来源情况 636株G-杆菌中,检出率最高的前4位是:鲍曼不动杆菌45.3%、肺炎克雷伯菌17.1%、铜绿假单胞菌14.3%和大肠埃希菌13.4%。ICU G-杆菌主要以痰标本来源为主,见表1。

表1 G-杆菌构成比及标本来源
2.2 ICU非发酵菌药敏实验结果 ICU非发酵菌的检出率(65.0%)高于肠杆菌科细菌(33.0%),其中鲍曼不动杆菌检出率最高(45.3%)。本研究鲍曼不动杆菌仅对多黏菌素(96.9%)、阿米卡星(78.8%)和妥布霉素(66.7%)敏感率较高,对其余抗菌药物耐药情况严重。铜绿假单胞菌虽然耐药率低于鲍曼不动杆菌,但可选用的抗菌药物同样不多,对多黏菌素(94.5%)、阿米卡星(66.0%)和哌拉西林(55.0%)敏感率高。嗜麦芽窄食单胞菌对抗菌药物中介率较高,也呈现多重耐药状况,仅对多黏菌素(94.1%)、米诺环素(94.1%)和喹诺酮类(>70%)敏感率较高。洋葱伯克霍尔德菌对3、4代头孢菌素、β内酰胺/β内酰胺抑制剂、米诺环素、复方新诺明的敏感率均在70%以上,见表2。
2.3 ICU肠杆菌科产ESBLs大肠埃希菌和肺炎克雷伯菌与非产ESBLs株药敏结果比较 ICU大肠埃希菌ESBLs分离率为70.6%,肺炎克雷伯菌ESBLs分离率为69.7%。亚胺培南和美罗培南外,产ESBLs大肠埃希菌和肺炎克雷伯菌株对抗菌药物耐药率明显比非产ESBLs株高。另外,产ESBLs株的肺炎克雷伯菌对抗菌药物敏感率低于产ESBLs株的大肠埃希菌,产ESBLs肺炎克雷伯菌耐药状况更严重。发现3株耐碳青霉烯类的大肠埃希菌和5株耐碳青霉烯类的肺炎克雷伯菌,见表3。

表2 ICU非发酵菌药敏实验结果(%)

表3 ICU产ESBLs株与非产ESBLs株药敏结果比较(%)
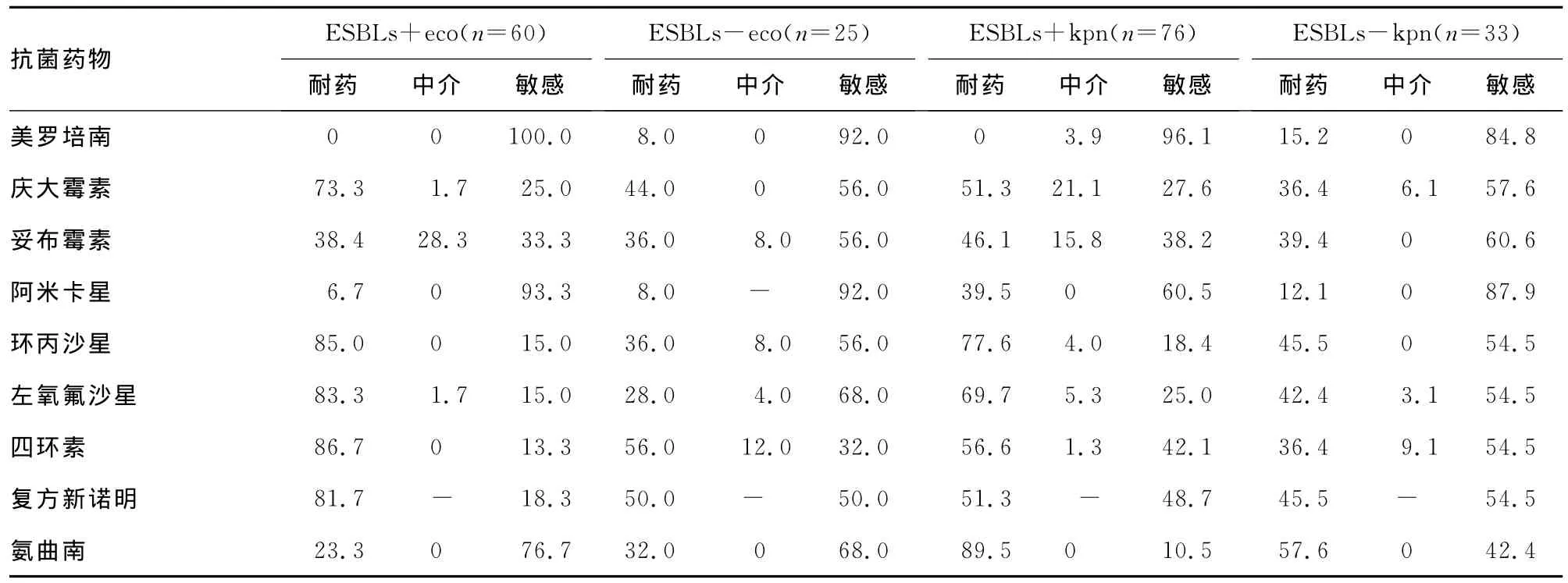
续表3 ICU产ESBLs株与非产ESBLs株药敏结果比较(%)
3 讨 论
该院ICU分离到的84.9%G-杆菌来自于痰样本,说明G-杆菌是ICU中引起呼吸道疾病的重要致病菌。而鲍曼不动杆菌、肺炎克雷伯菌、铜绿假单胞菌和大肠埃希菌是G-杆菌的重要构成者,与2011年全国细菌耐药监测网数据一致[2]。其中鲍曼不动杆菌是ICU检出率最高的细菌,与鲍曼不动杆菌是我国2008~2011年耐药监测网中ICU分离率最高的细菌一致[3-5]。说明ICU的感染以条件致病菌为主,这与其患者长期卧床、需建立人工气道、侵入性操作及病情严重有关。
本研究鲍曼不动杆菌多重耐药情况严重,仅对多黏菌素、阿米卡星和妥布霉素有较高的敏感性,对米诺环素中等敏感,对其余抗菌药物高度耐药。鲍曼不动杆菌对亚胺培南的耐药率为81.6%,多重耐药株数量增加,给临床治疗提出了严峻挑战[6-8]。鲍曼不动杆菌耐药机制复杂,包括基因突变、产生修饰酶和灭活酶、膜孔蛋白通道缺失,及发挥外排泵作用等多种机制共同作用[9-10]。多重耐药鲍曼不动杆菌提示人们光靠一类药物长期治疗鲍曼不动杆菌的感染是不妥的,采取联合用药来治疗或许是更好的方法,如碳青霉烯类或喹诺酮类加氨基糖苷类或β-内酰胺/β-内酰胺抑制剂复合物联合用药。有报道称头孢哌酮-舒巴坦和米诺环素的联合疗效令人满意(有效率达62.5%);本研究中多黏菌素对多重耐药鲍曼不动杆菌体外药敏实验也表现了良好的疗效,因而可作为治疗多重耐药鲍曼不动杆菌的备选药物[11]。
铜绿假单胞菌由于产生金属-β-内酰胺酶(MBLs)因而对碳青霉烯类药物耐药,同时GyrA 83、87位氨基酸和ParC87位氨基酸突变导致其对喹诺酮类药物耐药,更重要的是铜绿假单胞菌生物膜的形成使其对抗菌药物形成了天然屏障,使其成为检出率逐步上升且多重耐药的非发酵菌[12-14]。在本研究中仅对多黏菌素、阿米卡星和哌拉西林敏感性高。另外,嗜麦芽窄食单胞菌由于产生头孢菌素酶(AmpC酶)和碳青霉烯酶,可破坏头孢类抗菌药物和碳青霉烯类抗菌药物,也呈现多重耐药特征,对抗菌药物中介率较高,也是ICU常见的条件致病菌[15]。
产生ESBLs是肠杆菌科细菌对β-内酰胺类药物耐药的最主要机制,CTX-M型是世界大部分地区的主导基因型,我国以CTX-M-14、CTX-M-15和 CTX-M-3ESBLs为 主[16-17]。本 院ICU大肠埃希菌、肺炎克雷伯菌ESBLs分离率分别为70.6%和69.7%,与产ESBLs大肠埃希菌和肺炎克雷伯菌株对抗菌药物耐药率明显高于非产ESBLs株。大肠埃希菌ESBLs株除喹诺酮和四环素外对其余抗菌药物中介率比非产ESBLs株高;肺炎克雷伯菌ESBLs株除头孢曲松和四环素外对其余抗菌药物中介率比非产ESBLs株高,尤其β内酰胺/β内酰胺抑制剂体外药敏结果产ESBLs株的中介率高于非产ESBLs株。同时,肺炎克雷伯菌ESBLs株对抗菌药物敏感率低于大肠埃希菌ESBLs株,产ESBLs肺炎克雷伯菌耐药状况更严重,对其敏感性较高的药物仅有亚胺培南、美罗培南和阿米卡星,对产ESBLs大肠埃希菌则敏感性较高的药物除亚胺培南、美罗培南和阿米卡星外,还可选用头孢哌酮/舒巴坦和氨曲南。对非产ESBLs大肠埃希菌可使用3、4代头孢菌素、β内酰胺/β内酰胺抑制剂、碳青霉烯类、氨基糖苷类抗菌药物。
2010~2012年本院ICU共发现8株耐碳青霉烯类的肠杆菌科细菌。根据耐药表型推测为质粒介导的KPC型碳青霉烯酶的耐药传播,KPC型酶活性可被β-内酰胺抑制剂如克拉维酸和他唑巴坦抑制。我国自从检出KPC-2型酶后,KPC型酶在肠杆菌科细菌中有一定程度流行,应引起临床重视[18-19]。
综上所述,本院ICU分离到的G-杆菌以非发酵菌为主,并具有多重耐药特征,临床可选择多黏菌素或与氨基糖苷类联合用药进行治疗。大肠埃希菌和肺炎克雷伯菌ESBLs分离率高,检出耐碳青霉烯类的大肠埃希菌和肺炎克雷伯菌,ICU应加强耐药菌株监测,并做好带菌患者的消毒隔离工作,控制病原菌传播。
[1] Clinical and Laboratory Standards Institute(CLSI).Performance standards for antimicrobial susceptibility testing[S].CLSI,M100-S21,2011.
[2] 朱任媛,张小江,杨启文,等.卫生部全国细菌耐药监测网2011年ICU来源细菌耐药监测[J].中国临床药理学杂志,2012,28(12):905-909.
[3] 张丽,杨文航,肖盟,等.2010年度卫生部全国细菌耐药监测网报告:ICU来源细菌耐药性监测[J].中华医院感染学杂志,2012,20(1):34-38.
[4] 陈宏斌,张小江,赵颖,等.卫生部全国细菌耐药监测网(Mohnarin)2009年度报告:ICU来源细菌耐药监测[J].中国临床药理学杂志,2011,27(7):483-489.
[5] 肖永红.Mohnarin 2008年度ICU细菌耐药性监测[J].中华医院感染学杂志,2010,20(16):2384-2388.
[6] 李苏利,李杨,华川,等.多重耐药鲍氏不动杆菌耐药现状及临床对策[J].中国医院感染学杂志,2005,15(12):1438-1440.
[7] 王刚,黄洁,秦帅,等.不同抗菌药物对ICU泛耐药鲍氏不动杆菌感染的疗效比较[J].上海交通大学学报:医学版,2011,31(1):111-114.
[8] Anton Y,David C.Hospital-acquired infections due to gram-negative bacteria[J].NEJM,2010,362(19):1804-1813.
[9] 王玮玮,王厚照.多重耐药鲍氏不动杆菌耐药机制研究进展[J].国际检验医学杂志,2011,32(21):2487-2489.
[10] 李苏利,李杨,华川,等.多重耐药鲍氏不动杆菌耐药现状及临床对策[J].中国医院感染学杂志,2005,15(12):1438-1440.
[11] Huang J,Tang YQ,Sun JY.Intravenous colistin sulfate:a rarely used form of polymyxin E for the treatment of severe multi-drug resistant gram-negative bacterial infections[J].Scand J Infect Dis,2010,42(4):260-265.
[12] Wang C,Wang J,Mi Z.Pseudomonas aeruginosa producing VIM-2Metallo-beta-lactamase and carrying two aminoglycoside-modifying enzymes in China[J].J Hosp Infect,2006,62(4):522-524.
[13] Heisig P.Inhibitors of bacterial topoisomerases:mechanism of action and resistance and clinical aspects[J].Planta Med,2001,67(1):3-12.
[14] Akasaka T,Tanaka M,Yamaguchi A,et al.TypeⅡ topoisomerase mutations in fluroquinolone-resistant clinical strains of Pseudomonas aeruginosa isolated in 1998and 1999:role of target enzyme in mechanism in fluroquinolone resistance[J].Antimicrob Agents Chemother,2001,45(8):2263-2268.
[15] 佘军,单才华.嗜麦芽寡养单胞菌β-内酰胺酶检测及耐药性分析[J].中华医院感染学杂志,2008,18(8):1158-1160.
[16] Livermore DM,Canton R,Gniadkowski M,et al.CTX-M:changing the face of ESBLs in Europe[J].J Antimicrob Chemother,2007,59(2):165-174.
[17] Liu W,Chen L,Li H,et al.Novel CTX-Mβ-lactamase genotype distribution and spread into multiple species of Enterobacteriaceae in ChangSha,Southern China[J].J Antimicrob Chemother,2009,63(5):895-900.
[18] Wei ZQ,Du XX,Yu YS,et al.Plasmid-mediated KPC-2in a Klebsiella pneumoniae isolate from China[J].Antimicrob Agents Chemother,2007,51(2):763-765.
[19] 姚慧琳,范德胜,陆士海,等.产 KPC-2型碳青霉烯酶肠杆菌整合子分布研究[J].临床检验杂志,2012,30(4):281-283.
