单波长太阳能灯诱杀核桃楸大蚕蛾1)
2014-03-08王建军栾庆书金若忠云丽丽王琴姜旭
王建军 栾庆书 金若忠 云丽丽 王琴 姜旭
(辽宁省林业科学研究院,沈阳,110032)
责任编辑:程 红。
核桃楸大蚕蛾(Dictyoploca japonica),又名银杏大蚕蛾、白果蚕、白毛虫、漆毛虫,属鳞翅目(Lepidoptera)大蚕蛾科(Saturniidae)胡桃大蚕蛾属(Dictyoploca),在我国分布广泛,是一种重要的林业和药用植物害虫[1],[2]996。它主要以幼虫食害核桃、银杏、漆树、杨、桦、栎、李、梨等20 科30 属38 种经济林木[3]。核桃楸大蚕蛾在我国1年发生1 代,以卵越冬。雌蛾体长26~60 mm,翅展95~150 mm,雄蛾体长25~40 mm,翅展90~125 mm,体色不一,灰褐、黄褐或紫褐色[2]996。在辽宁抚顺、本溪、丹东等地,越冬卵于5月中旬羽化,6月下旬至7月上旬老熟幼虫营茧,成虫8月下旬至9月羽化,交配产卵后完成1 个世代[4-7]。近年来,该虫在我国一些地区频繁发生,局部暴发成灾且有蔓延趋势,给部分地区造成巨大的损失[8-9]。
为保证生态安全,国外防治此类食叶害虫多采用综合防治技术,其中生物防治比例较高[10]。而国内防治核桃楸大蚕蛾多使用化学药剂,导致害虫的抗药性增加和生态环境的破坏[10]。在森林害虫特别是食叶害虫的综合治理中,灯光诱杀常作为一种经济、简便和不污染环境的治虫措施加以利用[11]。利用频振式杀虫灯防治棉田害虫[12]和舞毒蛾[13],以及利用单波长的太阳能灯诱杀舞毒蛾[14]都取得较好的灭虫效果。在防治核桃楸大蚕蛾方面,也有使用频振式杀虫灯诱杀成虫的报道[15],但缺乏系统性的研究,更未见利用单波长太阳能灯诱杀成虫的报道。核桃楸大蚕蛾羽化期较长,在辽宁省本溪地区从8月下旬到9月下旬都有成虫羽化[7]。为了更有效地控制核桃楸大蚕蛾的危害,2013年开展了不同波长的灯光诱杀试验。通过灯诱,筛选诱杀核桃楸大蚕蛾的最佳波长和最适时段,比较不同时段灯诱核桃楸大蚕蛾的雌雄性比,选择装水容器的颜色和高度,解剖灯诱雌虫并统计其卵量等。旨在探索诱杀核桃楸大蚕蛾的最佳灯诱组合和检测所诱成虫的携卵状态,达到采用无公害手段压低害虫种群数量,安全有效、绿色环保的控制核桃楸大蚕蛾的目的。
1 试验地概况
试验地位于辽宁省桓仁满族自治县向阳乡向阳村于家街,占地面积约为10 hm2,树龄15~20 a,天然次生核桃楸林,株高超过15 m,林木郁闭度0.65。据前期调查,核桃楸大蚕蛾虫口密度为23.9 头·株-1,属中度危害。
2 材料与方法
2.1 材料
诱虫灯由深圳市富巍盛科技有限公司生产,型号为FWS-DBL-1,功率8 W,是精确的单波长灯(共18 种波长,为富巍盛公司的专利产品),对照灯(CK)是同等功率的全光谱灯。
2.2 研究方法
灯诱的最佳波长与时间筛选:于8月下旬,选择晴天无风的夜晚,将不同波段的太阳能灯置于林间距离地面1.5 m 的高处,灯距为30 m,灯下30 cm 处放置盛水容器(直径80 cm,高15 cm),加入3 L 清水、10 g 洗衣粉。将波长为320、340、351、360、365、368、380、400、420、445、460、480、520、525、545、560、575、585 nm 的诱虫灯和对照灯(CK)共19 盏,按上述方式在核桃楸林地一字排开。每晚于黄昏时18:30开灯,于19:30、20:30、21:30、22:30、23:30 各收集一次落到水中的成虫,记录上述5 个时段不同诱虫灯诱杀核桃楸大蚕蛾的数量和性别;记录不同波长太阳能灯诱杀非靶标昆虫(其他鳞翅目、鞘翅目和膜翅目昆虫)的种类和数量。每处理重复3 次。
自然羽化的成虫与灯诱成虫性别比较:8月上旬,在同一地点林中的树干、灌木丛、杂草丛等位置随机采集核桃楸大蚕蛾蛹,运回室内放在圆形透明玻璃瓶中(直径7 cm,高10 cm),用透明纱布盖住瓶口并用橡皮筋束紧,在室温条件下待成虫羽化。记录羽化成虫的数量和性别,将室内羽化成虫的雌雄比与野外灯诱成虫的雌雄比作比较。
盛水容器颜色及放置高度对诱虫量的影响:8月下旬,于成虫羽化盛期,在林间放置1 盏365 nm的诱虫灯,以灯为中心辐射状于地面放置银色、蓝色、绿色、黄色、白色共5 种颜色的装水容器,围成一个圆圈。18:30 开灯,分别在19:30、20:30、21:30、22:30、23:30 各收集一次落到水中的成虫,统计不同颜色的容器中诱杀核桃楸大蚕蛾的数量,重复3次。将2 盏波长为365 nm 的诱虫灯在核桃楸林地紧挨(距离0.5 m)放置(避免由于地点不同核桃楸大蚕蛾种群密度的差异),并在灯下各放置1 个盛水容器,1 个距地面1.2 m,另1 个放在地面上。18:30 开灯,在19:30、20:30、21:30、22:30、23:30 各收集1 次落到水中的成虫,统计两种不同高度容器中诱杀核桃楸大蚕蛾的数量。重复3 次。
灯诱雌虫携卵状况:解剖、统计所有诱集到的雌虫的剖腹卵量,并按剖腹卵量的大小将携卵量分为不同层次,分析不同层次所占比例。
数据统计与分析:所有数据用SPSS 13.0 分析。不同波长和不同时段对核桃楸大蚕蛾的诱杀量,对非靶标昆虫诱杀量和不同颜色盛水容器的诱杀量,使用One -Way ANOVA 中Dunnett T3 检验进行比较;不同时段诱杀核桃楸大蚕蛾的雌雄性比使用r×c 列联表的卡方检验;采集的茧蛹在室内羽化的大蚕蛾成虫与灯诱成虫的雌雄性比差异使用2 ×2列联表的卡方检验;放置于不同高度诱杀容器的诱杀量差异使用独立样本T 检验。
3 结果与分析
3.1 诱杀核桃楸大蚕蛾的最佳波长
供试诱虫灯对核桃楸大蚕蛾的诱杀总量、雌虫量、雄虫量的趋势一致(表1)。360 nm 诱虫灯的诱杀总量最大(11.7 头),且显著大于对照灯(CK)和其他诱虫灯的诱杀总量(F =3.003,df =18,p <0.05)。360 nm 的诱虫灯诱集雌虫最大量为7.3 头,诱集的雄虫的最大量为4.3 头。

表1 不同波长太阳能灯诱杀核桃楸大蚕蛾成虫的数量
3.2 不同时段诱杀核桃楸大蚕蛾的数量和雌雄性比
诱虫量随时间延迟呈先增加后减少的趋势,19:30—20:30(F =19.813;df =3;p <0.05)和20:30—21:30(F =8.196,df =3,p <0.05)的诱虫量都显著大于其他时段。5 个时段诱杀核桃楸大蚕蛾雌雄性比呈先增加后减少的趋势,χ2检验结果显示,5个时段雌雄性比无显著差异(χ2=1.015,p >0.05)。19 盏诱虫灯诱杀核桃楸大蚕蛾的总量为(71.6 ±8.6)头,其中雌蛾(50.0 ±8.5)头,雄蛾(21.6 ±17.0)头,雌雄比2.3∶ 1。不同时段诱杀量比例和雌雄性比见表2。
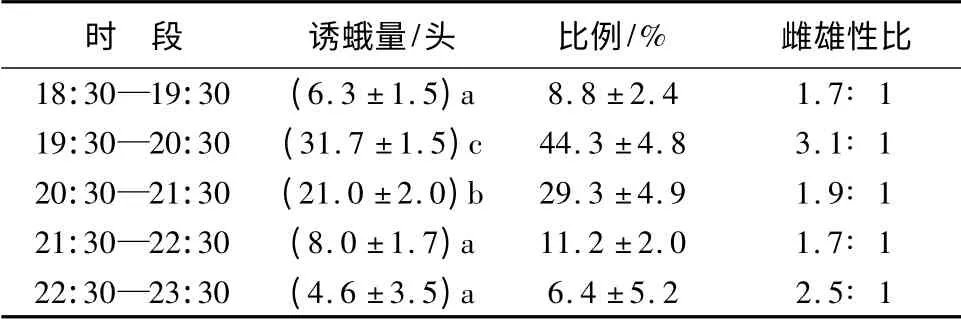
表2 不同时段灯诱核桃楸大蚕蛾的数量、比例和雌雄性比
野外采集的核桃楸大蚕蛾蛹共有1 239 头,最终羽化的成虫数量为869 头,羽化率70.1%。其他蛹由于天敌破坏,感染细菌、病毒或者在采集及运输过程中的损坏等原因没有羽化。羽化部分(70.1%)的雌雄性比为0.8∶ 1.0;灯诱核桃楸大蚕蛾成虫雌雄性比为2.3∶ 1,二者差异显著(χ2=23.5,p<0.05)。
3.3 盛水容器颜色和不同高度对诱虫量的影响
使用银色、蓝色、绿色、黄色、白色5 种颜色的盛水容器,360 nm 太阳能灯诱杀核桃楸大蚕蛾的平均数量分别为(2.3 ±0.8)、(2.3 ±0.4)、(0.7 ±0.2)、(0.7±0.2)、(2.7 ±0.2)头。其中,白色容器的诱杀量最大,且显著大于黄色和绿色容器的诱杀量(F=12.0;df =2;p <0.05,但与银色和蓝色容器的诱杀量相比无显著差异(F=0.05,df=5,p >0.05)。
盛水容器放置于距地面1.2 m 高度时,360 nm太阳能灯诱杀核桃楸大蚕蛾的数量为(9.3 ±2.6)头。盛水容器放置于地面上时,360 nm 太阳能灯诱杀核桃楸大蚕蛾的数量为(14.0 ±5.6)头。两种高度处理结果差异不显著(t = -0.759,df =4,p >0.05)。
3.4 雌虫携卵比例和携卵量
试验共诱捕雌虫296 头,携卵量最高为366 粒,平均为(121 ±13)粒。其中携卵量0~20 粒的有80头,占27.0%;携卵量大于20 粒的有216 头,占73.0%。说明单波长太阳能灯对已产卵和未产卵的核桃楸大蚕蛾成虫都有诱杀作用,诱杀的雌虫大部分(73.0%)处于未产卵或部分产卵状态。
3.5 对非靶标昆虫的影响
诱虫灯对非靶标昆虫的诱杀量见表3。368 nm灯对其他鳞翅目昆虫的诱杀量最大(72.0 头),360 nm 灯的诱杀量次之(69.3 头),但二者与对照灯的诱杀量(41.3 头)均无显著差异(F =1.051,df=2,p>0.05);供试诱虫灯之间对鞘翅目和膜翅目昆虫的诱杀量无显著差异(F=0.952,df=18,p >0.05)。

表3 不同波长太阳能灯对非靶标昆虫的诱杀量
4 结论与讨论
360 nm 太阳能灯对核桃楸大蚕蛾诱杀总量最大,可以作为林间诱杀成虫的最佳波长。19:30—21:30 的诱虫量占诱虫总量的72.0%。装水容器置于地面上比置于1.2 m 高度时有更好的诱杀效果,可能是因为雌蛾腹部大、身体重,致使其飞行能力受到一定影响[2]998。诱虫灯对核桃楸大蚕蛾的诱杀量平均值较小,应该与其林间的种群密度相关,主要原因是核桃楸大蚕蛾的羽化期持续时间较长(约30 d)[7],羽化时间不集中。同年8月初在同一地点采集的核桃楸大蚕蛾蛹在室内羽化出的869 头成虫中,雌雄性比为0.8∶ 1.0,说明自然羽化的雌虫量小于雄虫。而单波长太阳能灯诱杀核桃楸大蚕蛾雌雄性比为2.3∶ 1,雌性多于雄性。由于与核桃楸大蚕蛾同时期羽化的昆虫较多,试验中对非靶标昆虫的诱杀量也较大。360 nm 灯对核桃楸大蚕蛾和同期羽化的其他鳞翅目都具有较好的诱杀效果,但对于大多数林木来说,鳞翅目是害虫,所以对多种鳞翅目昆虫的诱杀会减少其他害虫的危害,却不会增加核桃楸大蚕蛾对林木危害的风险;供试诱虫灯对鞘翅目和膜翅目昆虫的诱杀量均无显著差异,说明360 nm 诱虫灯对天敌或可能性天敌的杀灭作用不会显著大于其他波长灯,可部分维持天敌对核桃楸大蚕蛾的持续控制能力。
根据试验结果,考虑到太阳能灯的电量(一般能维持4~6 h)。可在核桃楸大蚕蛾的羽化期以19:30—21:30 为主要诱虫时段,选用360 nm 的太阳能诱虫灯和白色装水容器,容器放置于地面上,林间诱杀核桃楸大蚕蛾成虫,以降低核桃楸大蚕蛾的虫口密度,减少对林木的危害。大部分诱杀到的核桃楸大蚕蛾雌虫是携卵的,但诱捕到雌虫的携卵量和自然羽化雌虫的怀卵量相比是否有差异,还有待进一步研究。
[1] 孙琼华,罗昌文,邓锡枝,等.银杏大蚕蛾的生物学和防治技术研究[J].林业科学研究,1991,4(3):273 -279.
[2] 萧刚柔.中国森林昆虫[M].北京:中国林业出版社,1992:996-98.
[3] 冯有贵.康县银杏大蚕蛾的经济性状调查及利用[J].北方蚕业,2009,30(1):11 -12.
[4] 许水威,叶淑琴,王立明.用期距法预测银杏大蚕蛾发生期[J].辽宁林业科技,2003,(2):10 -12.
[5] 张彩华,刘凤云,齐荣和,等.栗蚕的生物学特性[J].中国蚕业,2005,26(2):50 -51.
[6] 杨宝山,张希科,曹兰娟,等.银杏大蚕蛾生物学特性及防治技术[J].农药,2008,47(2):153 -154.
[7] 高丹,宋莉莉.银杏大蚕蛾在本溪地区生活史及习性与测报方法[J].内蒙古林业调查设计,2009,32(5):123 -124.
[8] 朱红,张培芳,田耀辉,等.银杏大蚕蛾对甘肃陇南核桃的危害[J].落叶果树,2007(1):25 -26.
[9] 李有忠,张芳保,王培新,等.银杏大蚕蛾的灾变规律与控灾技术研究[J].中国森林病虫,2009,28(2):20 -22,34.
[10] 葛君,胡春祥.8 种植物源药剂对银杏大蚕蛾幼虫的毒力测定及防治[J].东北林业大学学报,2010,38(2):69 -72.
[11] 尤德康,宋玉双.杀虫灯已成为森林害虫综合治理的重要工具[J].中国森林病虫,2002,21(增刊):4 -6.
[12] 王秀梅,地力拜尔·艾买提,王海燕.频振式杀虫灯防治棉田害虫效果研究[J].现代农业科技,2011(19):186,188.
[13] 迟安荣.频振式杀虫灯诱杀舞毒蛾技术研究[J].内蒙古林业调查设计,2012,35(1):83,85.
[14] 栾庆书,杜勇,云丽丽,等.舞毒蛾灯光诱杀研究[J].中国森林病虫,2014,33(2):36 -40.
[15] 袁照蓉.银杏大蚕蛾发生的为害及物理防治[J].农技服务,2012,29(9):1049.
