长白山阔叶红松林土壤呼吸对氮沉降增加的响应1)
2014-03-06李伟斌金昌杰井艳丽吴家兵袁凤辉关德新王安志
李伟斌 金昌杰 井艳丽 吴家兵 袁凤辉 关德新 王安志
(中国科学院沈阳应用生态研究所,沈阳,110164)
责任编辑:戴芳天。
森林生态系统作为陆地生态系统的主体,维持着巨大的土壤碳库(占全球土壤碳库的73%),因此森林生态系统土壤呼吸的大小在维持全球生态系统碳平衡以及调节全球气候等方面有重要作用[1]。作为一个复杂的生物化学过程,土壤呼吸不仅受植被及微生物的影响,而且受到土壤温湿度、有机质质量分数、土壤pH值、土壤氮磷钾质量分数的共同影响[2]。近年来由于人工施氮和化石燃料的燃烧使大气沉降的氮量增加到原来的3~5倍,且还有增加的趋势[3]。虽然轻度的氮沉降会促进植被的生长,增加植被对大气CO2的固定量,但过量的沉降氮对陆地生态系统也会产生一些负面影响,如生物多样性的降低、土壤酸化等[4-5],进而影响生态系统的结构与功能,最终影响森林生态系统的土壤呼吸。所以在全球变暖与大气氮沉降等环境问题日益严重的情况下,研究森林土壤呼吸作用对大气氮沉降的响应是评估碳循环动态、预测全球碳收支的基础,已成为全球科学家们关注的热点。国内外学者通过模拟氮沉降实验展开了相关研究,如Cleveland和Townsend[6]在热带雨林的研究发现施氮增加了植物的细根生物量,从而促进了土壤呼吸速率;涂利华等[7]在华西雨屏区苦竹林的研究表明,氮沉降增加了微生物的生物量碳、氮含量,提高了土壤微生物呼吸速率,进而促进了土壤总呼吸;另外,贾淑敏等[8]研究发现氮沉降减少了红松与水曲柳的细根生物量,从而降低了土壤呼吸速率。总之,由于受众多因子的影响,土壤呼吸速率对氮沉降的响应在不同研究区域、不同生态系统都是不同的,主要表现为氮沉降对土壤呼吸的促进[6]、抑制[9-10]和无影响[11-12]3种情况。由此可见,森林生态系统土壤呼吸对氮沉降增加的响应仍然有很大的不确定性。
我国作为全球三大氮沉降地区之一,年均氮沉降量12.9 kg·hm-2·a-1[13]。现有关于氮沉降影响土壤呼吸的研究主要集中在华南[14]、西南[7,15]、中南[16]及蒙古草原[17]等地,而对北方温带森林的研究较少。长白山森林生态系统是我国典型的温带森林,氮沉降量为23.6 kg·hm-2·a-1(大气网络监测数据),已超过了我国平均氮沉降量以及欧洲一些国家的氮沉降量,因此展开长白山森林生态土壤呼吸对氮沉降的响应研究具有极为重要的意义。笔者通过野外模拟试验,探讨氮沉降增加对长白山阔叶红树林土壤呼吸通量的影响,为进一步揭示氮沉降对土壤呼吸的影响机理提供理论依据。
1 研究区概况
本试验于吉林省安图县二道白河镇(41°42'45″~42°45'18″N,127°33'30″~128°16'48″E)中国科学院长白山森林生态系统定位站1号标准地原始阔叶红松林内完成,试验区海拔高度748 m,坡度1°~5°。该区属受季风影响的温带大陆性山地气候,春季干燥,夏季短且温暖湿润,秋季凉爽,冬季寒冷且漫长。年平均气温3.6℃,无霜期约120 d,年降水700~800 mm,主要集中在4—9月份。林下土壤为在火山灰母质上发育的暗棕色森林土。
该原始阔叶红松林平均林龄180 a左右,主要建群种有红松(Pinus koraiensis)、水曲柳(Fraxinus mandshurica)、蒙古栎(Quercus mongolica)、椴树(Tiliaamurensis)和色木槭(Acer mono)。林分为复层结构,平均株高26 m,郁闭度0.8,立木约560株·hm-2。灌木主要有忍冬(Lonicera japonica)、东北山梅花(Philadelphus schrenkii)和毛榛(Corylus mandshurica)等。草本有苔草(Carex spp)、山茄子(Brachybotrys paridiformis)和一些蕨类等。
2 研究方法
2.1 实验设计
2011年初在1号标准地原始阔叶红松林内建立氮沉降试验样地,在样地中建立16个15 m×15 m的样方,每个样方之间设置大于5 m宽的缓冲区域。添加氮素为尿素,共设置对照,低氮(23 kg·hm-2·a-1),中氮(kg·hm-2·a-1)和高氮(69 kg·hm-2·a-1)4个氮添加水平,每个水平4个重复。从2011年6月开始,进行施氮处理。具体方法是根据氮处理水平将每个样方每次需要施加的尿素溶解在5 L水中,用喷雾器均匀地喷洒至样方内,对照样方则喷洒同样量的水。每年施氮分3次进行,分别于6、7、8月初等量施加。
2.2 土壤呼吸、温度的测定
在每个样方中随机选取2个点安放PVC管(内径10 cm,高4.5 cm),插入土壤约2 cm。从2012年5月下旬到10月31日,利用便携式LI-6400分析仪(Li-Cor Inc,Lincoln,NE,USA)对土壤呼吸速率进行定期测定(平均每月3、4次,测定区间在早上9:00—11:00内),每月至少测一次土壤呼吸的日变化,测定时间间隔为2 h。在测定土壤呼吸速率的同时,用LI-6400自带的温度探头同步测定5 cm处的土壤温度。
2.3 凋落物的收集
采用对角线法在每块样方的对角线上设置3个接凋落物的尼龙网(1 m×1 m),平均2个月收集凋落物一次,并用烘箱65℃烘干至恒质量,然后用电子天平称质量。
2.4 数据处理与分析
每月对土壤呼吸测定后,所得到的数据为每个测定点的土壤呼吸速率值,将不同处理中各个测定点的数据进行平均,即得到不同处理水平的平均土壤呼吸速率值。在不同处理中凋落物网收集的年凋落物量的平均值分别代表各个处理的年凋落物量。
采用指数方程V=a ebT来拟合土壤呼吸速率与土壤温度间的关系,其中V为土壤呼吸速率,T为土壤温度,a为土壤温度是0℃时的土壤呼吸速率,b为温度响应常数[18]。采用Q10进行土壤呼吸对温度的敏感性进行定量分析,具体方法是:先将土壤呼吸速率与对应的土壤温度用上式的指数函数进行回归拟合,然后用公式Q10=e10b计算[19]。
最后采用单因素方差分析和多重比较检验土壤呼吸的昼夜、季节变化、不同处理间土壤呼吸速率、温度、凋落物量的差异性。统计分析在SPSS、Excel软件中完成,绘图在Origin软件中完成。
3 结果与分析
3.1 氮沉降对土壤呼吸昼夜变化的影响
土壤呼吸的昼夜变化测定的结果如表1所示。总体而言,氮沉降没有改变土壤呼吸的昼夜变化趋势,各处理水平的土壤呼吸均存在明显的昼夜波动,且变化趋势相似。早晨土壤呼吸速率最小,傍晚土壤呼吸速率最大。各处理水平日均土壤呼吸速率从大到小依次为:中氮(3.21μmol·m-2·s-1)、低氮(3.07μmol·m-2·s-1)、对照(1.95μmol·m-2·s-1)、高氮(1.63μmol·m-2·s-1),由此可知,低、中氮处理促进了阔叶红松林土壤呼吸作用,而高氮抑制了呼吸作用。方差分析结果表明除了对照与高氮处理间、中氮与低氮处理间土壤呼吸速率差异不显著外,其他各处理之间都存在显著差异(P<0.05)。
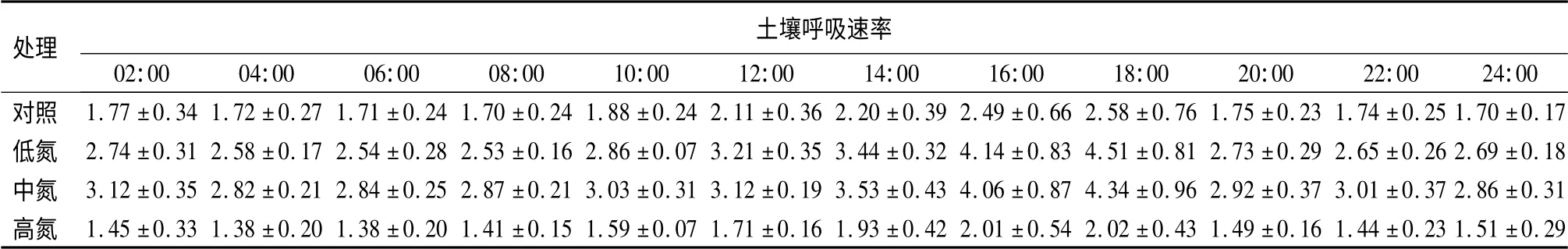
表1 土壤呼吸速率的昼夜变化 μmol·m-2·s-1
3.2 氮沉降对土壤呼吸季节变化的影响
对植被生长期内(5—10月份)不同月份的土壤呼吸测定后发现呼吸速率有明显的季节波动,其中对照组可代表长白山阔叶红松林在自然状态下的土壤呼吸情况(表2)。在生长期内,土壤呼吸速率变化明显,最大值为2.27μmol·m-2·s-1(7月份),最小值为0.72μmol·m-2·s-1(10月份),与温度的变化趋势基本一致。
在整个生长季内,低、中氮处理提高了阔叶红松林的土壤呼吸速率,高氮处理降低了阔叶红松林的土壤呼吸速率。低、中和高氮处理的年平均土壤呼吸速率分别为2.86,2.95、1.44μmol·m-2·s-1,与对照组(1.84μmol·m-2·s-1)相比较发现,低、中氮分别比对照高出55.4%和60.3%,高氮却比对照低21.7%。方差分析结果表明5、6月份的呼吸速率除了对照与高氮处理间、中氮与低氮处理间土壤呼吸速率的差异不显著外,其他处理间都存在显著差异;7、8、9月份只有高氮与中氮、高氮与低氮和中氮与对照处理的土壤呼吸速率间存在显著差异;而10月份只有高氮与低氮和高氮与中氮处理的土壤呼吸速率间存在显著差异。由此可知,在整个生长期的不同阶段(从展叶到叶落)氮处理对土壤呼吸速率影响的程度也存在差异。
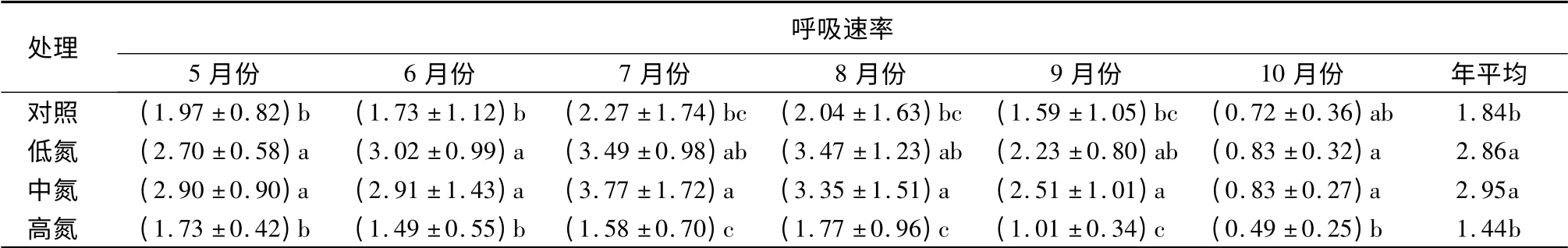
表2 土壤呼吸速率生长季内的变化 μmol·m-2·s-1
3.3 氮沉降对凋落物量和土壤温度的影响
对各处理水平间5 cm土壤温度、年凋落物量进行比较(表3)。可以看出,各处理水平之间的土壤温度没有明显差异,而氮沉降显著增加了凋落物量,对照、低氮、中氮和高氮处理的年总凋落物量分别为264.77、382.14、306.32、300.49 g/m2,与对照组相比高氮、中氮、低氮处理年凋落物量分别增加了13.49%、15.69%、44.32%。

表3 氮沉降对凋落物量和土壤温度的影响
3.4 土壤呼吸与土壤温度的关系
土壤温度往往是影响土壤呼吸速率的主要因素,通过对各处理水平土壤呼吸速率与5 cm土壤温度间的相关性进行回归分析(图1)可知:各处理的土壤呼吸速率与5 cm土壤温度都存在显著的指数关系(表4),其中中氮处理的相关性最好(R2=0.789),低氮处理的次之(R2=0.727),而高氮处理的相关性最差(R2=0.502)。利用回归方程中的温度响应常数计算得到对照、低氮、中氮和高氮处理水平土壤呼吸速率温度敏感性常数Q10分别为1.93、2.34、2.94和1.54。
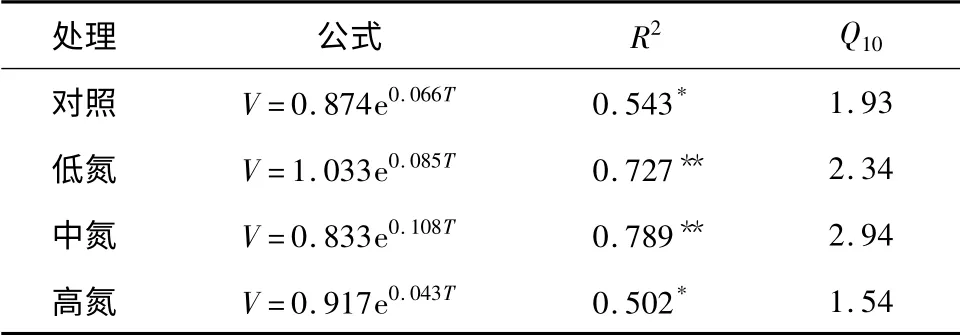
表4 土壤呼吸与土壤温度间的相关性结果
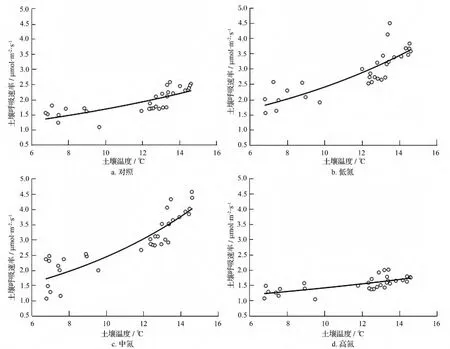
图1 土壤呼吸速率与5 cm处土壤温度的关系
4 讨论
4.1 阔叶红松林土壤呼吸对氮沉降的响应
大气氮沉降能够促进土壤的氮矿化率,改变土壤与凋落物的C/N比以及土壤微生物的活性,从而影响土壤呼吸速率[20]。然而关于氮沉降对森林土壤呼吸影响的研究结果不一,主要有增加[6-7]、降低[9-10,16]和无影响[11-12]3种。本次氮沉降模拟实验显示:低、中水平氮沉降显著促进了土壤呼吸,土壤呼吸速率分别提高了55.4%和60.3%,而高氮沉降却抑制了土壤呼吸作用,呼吸速率降低了21.7%,表明氮沉降对土壤呼吸存在着阈值效应。分析其原因,主要是该区域中土壤氮元素质量分数的本底值低于植被与微生物对氮素的需求量[21],而低、中水平氮素的增加正好满足了这种需求,增加了微生物的活性和植物的生长速率,即表现为促进了土壤呼吸速率。然而高氮添加使土壤中氮素质量分数达到饱和,此时氮素不再是限制微生物与植物生长的营养元素,因此氮素的添加可能会影响微生物群落,使植物体内的营养元素失衡,甚至会加剧磷、钙等营养元素的限制,引起土壤的酸化[22-23],在这种情况下土壤呼吸速率会明显降低。贾淑敏等[8]指出土壤呼吸降低的关键是氮沉降减少了植物的细根生物量。在本研究中,低、中氮处理还没有使土壤氮达到饱和,因此添加之后提高了土壤呼吸速率,这与涂利华等[7]在华西竹林的研究结果基本一致;而高氮处理使土壤氮质量分数达到了饱和,抑制了土壤呼吸作用,此结果与在中亚热带樟树林[24]、亚热带湿地人工林[25]和北亚热带落叶阔叶林[26]的研究结果相似。
本研究表明氮沉降明显增加了植被的凋落物量,这与氮沉降能促进植物生长、增加凋落物量从而增加碳汇的研究结论[3,9,27]一致。土壤呼吸主要由根呼吸、微生物呼吸和凋落物等有机质分解三部分组成,土壤呼吸对氮沉降的响应是各个组分对氮沉降响应的总和。在土壤氮质量分数没有达到饱和之前,氮沉降增加了根系组织与微生物细胞中的氮质量分数,从而促进了植物根系与微生物的代谢[11],增加了凋落物总量及其氮质量分数,加快了凋落物的分解速率[28-29],即增加了土壤有效碳的输入量,进而促进了土壤呼吸速率的增加。然而当土壤氮质量分数达到饱和时,进一步添加氮很难再增强根系与微生物的代谢以增加凋落物量,相反会增加根系组织与微生物对磷等其他营养元素的需求而起到反作用;再者C/N比是影响凋落物分解速率的主要因子[30-31],氮沉降降低了凋落物中的C/N比,加剧了其降解过程中的C限制,由于能量的供应不足减小了分解速率[28,32-33]。
4.2 氮沉降对阔叶红松林土壤呼吸温度敏感性的影响
本研究发现土壤呼吸速率随5 cm土壤温度的增加呈指数形式增长,这与国内外其他相关研究结果基本一致[15-16,34]。土壤呼吸是地下微生物、植物根系、动物代谢以及有机质分解作用的总和,土壤温度通过影响上述生理过程中的酶活性而影响土壤呼吸。长白山地区冬季漫长,年平均气温较低,温度升高增强了酶活性,加速了植物与微生物对氮素的利用效率和土壤有机质的分解[35],提高了土壤呼吸速率。因此土壤温度是该研究区内控制土壤呼吸的主要驱动因子。
土壤呼吸对温度的敏感性可以用Q10值来定量衡量。已有研究表明在不同的生态系统、不同的研究区域Q10值不同[36],即便是在同一研究区域内,土壤呼吸的各组分的Q10值也存在差异[15]。本研究中自然状态下的Q10值为1.93,介于温带阔叶(Q10=2.23)和针叶林(Q10=1.69)之间[34]。低氮与中氮处理的Q10值分别为2.34和2.94,均高于对照,而高氮处理的Q10值为1.54,低于对照。这说明低、中氮处理增强了土壤呼吸对温度的敏感性,而高氮处理则降低了土壤呼吸对温度的敏感性。这不同于氮沉降只增强[7]或只减弱[14]土壤呼吸温度敏感性的研究结论,可能是由于土壤微生物及根系细胞的代谢受制于土壤温度,中、低水平的氮添加增加了土壤微生物数量及活性还有根系细胞的酶活性,进而使土壤呼吸温度敏感性增强,而高氮处理则起到了反作用。
由于森林生态系统响应氮素增加存在着长期的累积效应和适应过程,相对于国外长达10 a多的慢速氮添加实验研究[37]来说,本研究由于模拟氮沉降处理时间较短,一些结论还需要进一步的深入研究来检验,特别是需要在植被和土壤碳库蓄积与周转的长期监测基础上,明晰长白山地区温带森林生态系统关键生态过程的氮沉降饱和阈值。
[1]Post W M,Emanuel W R,Zinke P J,et al.Soil pool and world life zones[J].Nature,1982,298:156-159.
[2]张东秋,石培礼,张宪洲.土壤呼吸主要影响因素的研究进展[J].地球科学进展,2005,20(7):778-785.
[3]Janssens IA,Dieleman W,Luyssaert S,et al.Reduction of forest soil respiration in response to nitrogen deposition[J].Nature Geoscience,2010,3(5):315-322.
[4]Hoegberg P,Fan H,Quist M,et al.Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest[J].Global Change Biology,2006,12(3):489-499.
[5]Dise N,Rothwell J,Gauci V,et al.Predicting dissolved inorganic nitrogen leaching in European forests using two independent databases[J].Science of the Total Environment,2009,407(5):1798-1808.
[6]Cleveland C C,Townsend A R.Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere[J].Proceedings of the National Academy of Sciences,2006,103(27):10316-10321.
[7]涂利华,胡庭兴,黄立华,等.华西雨屏区苦竹林土壤呼吸对模拟氮沉降的响应[J].植物生态学报,2009,33(4):728-738.
[8]贾淑霞,王政权,梅莉,等.施肥对落叶松和水曲柳人工林土壤呼吸的影响[J].植物生态学报,2007,31(3):372-379.
[9]Olsson P,Linder S,Giesler R,et al.Fertilization of boreal forest reduces both autotrophic and heterotrophic soil respiration[J].Global Change Biology,2005,11(10):1745-1753.
[10]Jia S,Wang Z,Li X,et al.N fertilization affects on soil respiration,microbial biomass and root respiration in Larix gmelinii and Fraxinus mandshurica plantations in China[J].Plant and Soil,2010,333(1/2):325-336.
[11]Samuelson L,Mathew R,Stokes T,et al.Soil and microbial respiration in a loblolly pine plantation in response to seven years of irrigation and fertilization[J].Forest Ecology and Management,2009,258(11):2431-2438.
[12]Jiang J,Zong N,Song M,et al.Responses of ecosystem respiration and its components to fertilization in an alpine meadow on the Tibetan Plateau[J].European Journal of Soil Biology,2013,56:101-106.
[13]LüC Q,Tian H Q.Spatial and temporal patterns of nitrogen deposition in China:Synthesis of observational data[J].Journal of Geophysical Research:Atmospheres,2007,112(D22):D22S05-1-10.
[14]Mo J,Zhang W,Zhu W,et al.Nitrogen addition reduces soil respiration in a mature tropical forest in southern China[J].Global Change Biology,2008,14(2):403-412.
[15]Tu L H,Hu T X,Zhang J,et al.Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem[J].Soil Biology&Biochemistry,2013,58:255-264.
[16]郑威,闫文德,王光军,等.施氮对亚热带樟树林土壤呼吸的影响[J].生态学报,2013,33(11):3425-3433.
[17]Yan L,Chen S,Huang J,et al.2010.Differential responses of auto and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe[J].Global Change Biology,16(8):2345-2357.
[18]Li Y L,Otieno D,Owen K,et al.Temporal variability in soil CO2emission in an orchard forest ecosystem[J].Pedosphere,2008,18(3):273-283.
[19]Xu M,Qi Y.Spatial and seasonal variations of Q10determined by soil respiration measurements at a Sierra Nevadan forest[J].Global Biogeochemical Cycles,2001,15(3):687-696.
[20]吴家兵,井艳丽,关德新,等.氮沉降对森林碳汇功能影响的研究进展[J].世界林业研究,2012,25(2):12-16.
[21]胡艳玲,韩士杰,李雪峰,等.长白山原始林和次生林土壤有效氮含量对模拟氮沉降的响应[J].东北林业大学学报,2009,37(5):36-38.
[22]Matson P M,McDowell WH,Townsend A R,et al.The globalization of N deposition:ecosystem consequences in tropical environments[J].Biogeochemistry,1999,46(1/3):67-83.
[23]Zhang G,Taylor GJ.Effects of biololgical inhibitors on kinetics of aluminum uptake by excised roots and purified cell wall material of aluminum-tolerant and aluminum-sensitive cultivars of Triticum aestivum[J].Journal of Plant Physiology,1992,138(5):533-539.
[24]张徐源,闫文德,马秀红,等.模拟氮沉降对樟树人工林土壤呼吸的短期效应[J].中南林业科技大学学报,2012,32(3):109-113.
[25]张徐源,闫文德,郑威,等.氮沉降对湿地松林土壤呼吸的影响[J].中国农学通报,2012,28(22):5-10.
[26]胡正华,李涵茂,杨燕萍,等.模拟氮沉降对北亚热带落叶阔叶林土壤呼吸的影响[J].环境科学,2010,31(8):1726-1732.
[27]Pregitzer K S,Burton A J,Zak D R,et al.Simulated chronic nitrogen deposition increases carbon storage in Northern Temperate forests[J].Global Change Biology,2008,14(1):142-153.
[28]Kuperman R G.Litter decomposition and nutrient dynamics in oak-hickory forests along a historic gradient of nitrogen and sulfur deposition[J].Soil Biology&Biochemistry,1999,31(2):237-244.
[29]Vestgarden L S.Carbon and nitrogen turnover in the early stage of Scots pine(Pinus sylvestris L.)needle litter decomposition:effects of internal and external nitrogen[J].Soil Biology&Biochemistry,2001,33(4/5):465-474.
[30]Zhang D,Hui D,Luo Y,et al.Rates of litter decomposition in terrestrial ecosystems:global patterns and controlling factors[J].Journal of Plant Ecology,2008,1(2):85-93.
[31]Prescott C E.Litter decomposition:what controls it and how can we alter it to sequester more carbon in forest soils[J].Biogeochemistry,2010,101(1/3):133-149.
[32]Berg B.Litter decomposition and organic matter turnover in northern forest soils[J].Forest Ecology and Management,2000,133(1/2):13-22.
[33]Song M H,Jiang J,Cao G M,et al.Effects of temperature,glucose and inorganic nitrogen inputs on carbon mineralization in a Tibetan alpine meadow soil[J].European Journal of Soil Biology,2010,46(6):375-380.
[34]Bowden R D,Davidson E,Savage K,et al.Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest[J].Forest Ecology and Management,2004,196(1):43-56.
[35]杨万勤,王开运.森林土壤酶的研究进展[J].林业科学,2004,40(2):152-159.
[36]Boone R D,Nadelhoffer K J,Canary J D,et al.Roots exert a strong influence on the temperature sensitivityof soil respiration[J].Nature,1998,396:570-572.
[37]Entwistle E M,Zak D R,Edwards I P.Long-Term Experimental Nitrogen Deposition Alters the Composition of the Active Fungal Community in the Forest Floor[J].Soil Science Society of America Journal,2013,77(5):1648-1658.
