半干旱区人工封育草场植被群落生态位研究
——以宁夏盐池县长期定位监测点为例
2014-02-28刘建康张克斌王黎黎王志述王冠琪
刘建康,张克斌,王黎黎,王志述,王冠琪
北京林业大学水土保持与荒漠化防治教育部重点实验室,北京100083
半干旱区人工封育草场植被群落生态位研究
——以宁夏盐池县长期定位监测点为例
刘建康1,张克斌2,王黎黎3,王志述3,王冠琪3
北京林业大学水土保持与荒漠化防治教育部重点实验室,北京100083
人工封育是草场恢复和重建的一种主要措施。为研究半干旱区人工封育草地植物种群生态位,以宁夏盐池县人工封育区3种不同处理方式(长期完全封育、短期完全封育、短期不完全封育)为研究对象,将封育区分为封育核心区(E区)、封育边缘区(E1区)和封育外围区(E2区),通过分析在该地区出现的27种植物种群的Levins生态位宽度指数和Pianka生态位重叠指数,揭示植物种群生态位在干旱荒漠环境下的变化规律,为进一步分析该地区群落稳定性提供依据。研究结果表明:1)核心区和外围区生态位宽度最大的都是刺沙蓬Salsola ruthenica lljin,分别为0.832和0.810,而边缘区生态位宽度最大的是丝叶山苦荬Lxeris chinensis var. graminifolia,为0.742,这2种植物的生物学特性决定了它们能够很好的适应封育区的环境,并很好的利用封育区的资源,结果是它们的生态位宽度明显高于其他物种。2)对三区内不同植物种群的生态位宽度分析结果显示,三区的群落稳定性表现为E1区>E2区>E区;长期的完全封育并不利于封育区内植物群落的稳定;而短期的完全封育措施比不完全封育措施更有利于植被群落的稳定。3)通过分析三区内不同植物种之间的生态位重叠得出,E区生态位重叠最大值出现在草地风毛菊Saussurea amara、苦豆子Sophora alopecuroides L、沙芦草Agropyron mongolicum Keng与臭蒿Artemisia hedinii之间,都是0.98。E1区生态位重叠最大值是达乌里胡枝子Lespedeza davurica和草木樨状黄芪Astragalus melilotoides之间(0.82)。E2区生态位重叠最大的一对是草木樨状黄芪和冰草Agropyron cristatum(0.99)。相关分析表明:生态位宽度与生态位重叠之间并没有明确的线性关系。生态位宽度窄的物种之间可能有较高的生态位重叠,反之,则较低,但并不是完全如此。
封育;草原植被;生态位宽度;生态位重叠;半干旱区;群落稳定性
生态位(Niche)是一个即抽象,而又富有内涵的生态学名词,它作为生态学上的一个重要概念,最早是由Joseph Grinnel(1917)于1917年提出的。Grinnell认为生态位是“恰好被一个种或一个亚种所占据的最后单位”,他强调生态位的空间概念,所以我们可以把生态位理解为空间生态位(Space Niche)。自Grinnell提出生态位概念以来国外的许多学者又提出了多种不同的定义(Elton,1927;Cause,1934;Krebs,1978),如Elton(1927)提出的“营养生态位”(生态位的定义是它在生物环境中的位置及它与食物和天敌的关系)、Gause(1934)提出的“竞争排斥原理”(竞争使两个相似的物种极少占据相同的生态位)等。他们还对生态位宽度测度、生态位在不同资源条件下的变化与适应、植物种群对资源的分割利用、物种生态位关系与种间竞争和共存的联系等进行了细致深入的研究。目前,国内学者对生态位及测度方法(王刚等,1984;尚玉昌,1988;朱春全,1993)方面的研究也取得了一定的成绩,如李军玲等(2003)对关帝山亚高山灌丛草甸群落优势种群的生态位进行了研究,其结果表明,种群生态位宽度越大,对环境的适应能力越强,对资源的利用能力也越强;种群生态位重叠越大,种群间的生态相似性越大,利用资源的相似性程度越高;生态位宽的种群对生态位窄的种群可能有较高的重叠值,反之则较低。
近年来随着生态位理论的发展,生态位研究已
经成为理论生态学的一个重要研究内容。目前,对多种植物类型的生态位都有研究,如亚热带乔木(汪建华等,2001)、亚高山草甸(王刚和张大勇,1991)、高寒草甸(魏志琴等,2004)等,但是关于半干旱区人工封育草地植物种群生态位方面的研究相对较少,而这正是本文研究的重点。本文结合国家林业局定点监测项目(宁夏盐池荒漠化监测项目),通过对2013年荒漠化草原人工封育区27种植物的研究,分析它们的生态位情况,以揭示植物生态位在干旱荒漠环境下的变化规律,同时进一步分析该地区植物群落稳定性,这对荒漠化地区的生态恢复工作有着重要的理论和实践意义。
1 研究区自然概况
盐池县位于宁夏回族自治区东部长城沿线,全县南北长110 km,东西宽66 km,辖区总面积8661.3 km2,全县位于北纬37°04′~38°10′、东经106°30′~107°41′之间,是宁夏面积最大的县,占全区总面积的16.7%。盐池县地处毛乌素沙漠南缘,黄土高原北部,属于典型的过度地带,即:在地形方面,是从南向北由黄土高原向鄂尔多斯台地(沙地)过渡的地带;在气候方面,是自半干旱区向干旱区过渡的地带;在植被方面,是从干草原植被向荒漠植被过渡的地带;在资源利用方面,是从农区向牧区过渡的地带。正是这种在气候、地理等自然条件方面的过渡性使该县自然资源存在多样性和脆弱性等特点。
盐池县地形主要为剥蚀的准平原,境内地势南高北低,海拔在1295~1951 m之间,南部为黄土丘陵区,中北部为鄂尔多斯缓坡丘陵区。该县常年干旱少雨,风大沙多,年均气温为8.1 ℃,极端最高均温为34.9 ℃,极端最低温为-24.2 ℃,年均无霜期为165 d,年降水量仅在250~350 mm之间,且由南向北、由东南向西北递减,但年蒸发量多达2100 mm,属于典型的中温带大陆性气候。土壤类型主要是灰钙土,其次是黑沪土和风沙土,还有些许黄土及少量盐土、白浆土等。植被在区系上属于欧亚草原区亚洲中部亚区,是中国中部草原区的过度地带,它包含多种植被类型,主要是灌丛、草原、草甸、沙地植被和荒漠植被,其中灌丛、草原、沙地植被不仅数量较大,而且分布也较广。植被群落中常见植物种类主要是旱生和中旱生类型,其中旱生类型主要分布于干草原区,中旱生类型主要分布于草甸区。
2 研究方法
2.1 样地设置
本文根据当地主要土地利用(荒漠化)类型及主要荒漠化治理工程种类,按照典型性、科学性和代表性的设置原则,将研究区域选在毛乌素沙地西南缘的距盐池县城20 km处的柳杨堡乡人工封育区。本实验将封育区按长期完全封育、短期完全封育、短期不完全封育3种不同的处理方式,分别分为封育核心区(E区)、封育边缘区(E1区)和封育外围区(E2区),且三区在一条直线上(图1)。封育核心区始于1991年第1批全国防沙治沙试验示范,用铁丝网作围栏,目的在于完全排除野生动物和家畜的采食;封育边缘区自2002年以来采取完全封育措施;封育外围区虽然也从2002年开始进行封育,但仍受一些人类活动(如放牧、樵采)的影响。3种处理方式相距较近,自然条件差异不大。
2.2 野外调查
本次调查选取时间为2013年生长季节(8月),方法是从E开始沿着图1中箭头所指的方向在E、E1、E2内各随机布设10个1 m×1 m的样方,共30个样方。主要调查内容有:植物名称、植物种数、株数、盖度、高度、生物量(鲜质量)等。
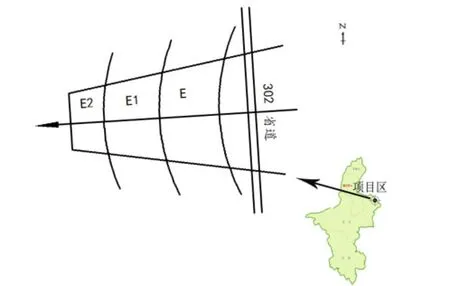
图1 封育区分布图Fig.1 The distribution map in fencing area
2.3 数据处理方法
2.3.1 重要值
植被数量生态学中的重要值(Important Value)最早是由Curtis和Mclntosh(1951)在研究森林群落时提出的,他们认为“重要值是综合衡量某个物种在森林群落中作用和地位的数量指标”,我们可以通过分析物种的重要值来了解群落种群的变动情况。尽管不同学者对于重要值的表达有着不同的见解,但是真正生态学意义上的重要值公式(王育松和上官铁梁,2010)如下:
重要值=(相对多度+相对频度+相对盖度+相对高度+相对生物量)/5
2.3.2 生态位宽度
生态位宽度反映的是物种或种群对环境适应的状况或对资源利用的程度(李德志等,2006)。本文采用Levins生态位宽度计测公式(Levins,
1968):
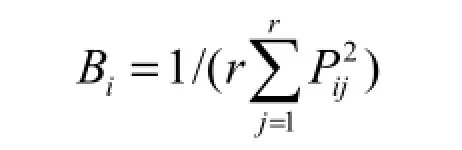
式中:Bi表示物种i的生态位宽度;Pij表示物种i在第j资源位上的重要值占它所在全部资源位上重要值的比例;r表示样方数。
2.3.3 生态位重叠
生态位重叠是生态位计测过程中的一个重要指标,是指不同物种之间生态位重叠或共有生态位空间的现象,即两个或两个以上的物种分享或竞争资源位或资源状态的现象(张润涛和郭健,2008)。本文采用Pianka生态位重叠指数(Pianka,1973)来计测生态位重叠,公式如下:
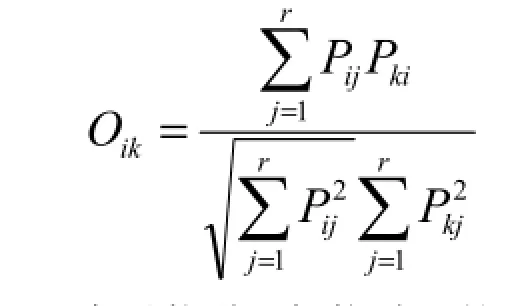
式中:Oik表示物种i与物种k的生态位重叠指数,其他符号含义同上式。
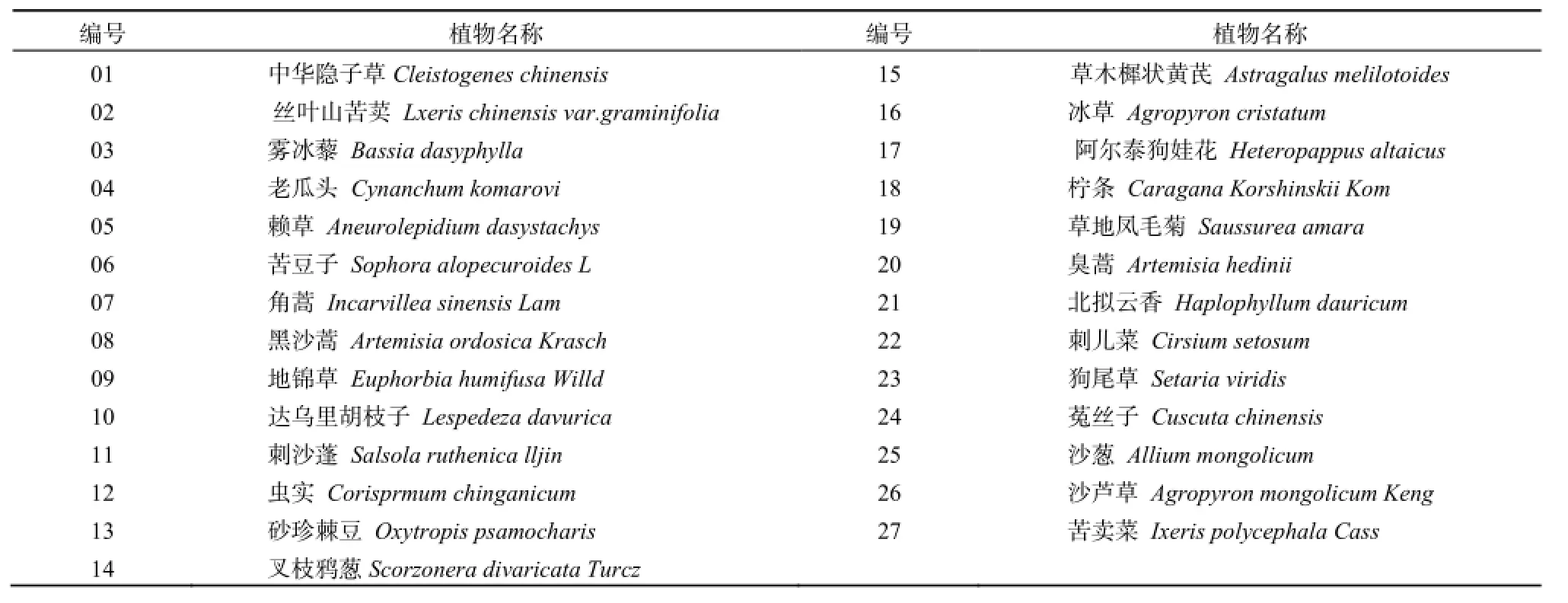
表1 宁夏盐池县人工封育区植被名称编号Table 1 Plant names and code in the enclosed grassland plots at Yanchi County of Ningxia Province
3 结果与分析
在野外调查中总共出现27种植物,对这些植物进行统一编号(表1),本文中生态位宽度表和生态位重叠表中出现的植物名称均由此编号代替。
3.1 生态位宽度分析
由于生态位宽度常用来表示被一个物种所利用的不同资源位的总和,因此,生态位宽度值的大小体现了种群在群落中的竞争地位,种群生态位越宽,表明在群落中竞争能力越强;种群生态位越窄,表明在资源环境的竞争中处于劣势地位(李军玲等,2003)。而群落稳定性是指在外界因子或干扰活动的作用下,种群各组分抵抗变化或是保持平衡状态的倾向(Odum,1971)。环境变化、外来物种入侵和人类活动都能影响群落的稳定性,而判断群落稳定性的两个最重要因素是群落的抵抗力和恢复力,种群的组成、结构和功能可作为衡量稳定性的重要尺度。
由表2可知,2013年封育核心区共出现22种植物,生态位宽度最大的3种是刺沙蓬Salsola ruthenica lljin(0.832)、丝叶山苦荬Lxeris chinensis var. graminifolia(0.815)和角蒿Incarvillea sinensis Lam(0.766)。其中刺沙蓬以其特殊的环境适应性出现在调查的每一个样方中,占据最多的资源。如刺沙蓬属温带旱生植物,耐干旱能力较强,株丛的大小和多少主要受降雨量的制约;对土质的要求不高,除潮湿和已经盐渍化的土壤外,在其它多种土壤中均可生长,并主要生于沙质或少陈质土壤之上。以上3种植物的生态位宽度远远高于其他物种,这表明它们是核心区的优势物种,在群落中地位最高,对资源环境的利用能力最强,占据了最多的自然资源和生存空间,这必然抑制其他物种在群落中的生长,导致该区其他物种生态位宽度较小,生态位宽度平均值在3种不同处理方式中最小,而方差却最大(图2),这说明该区植被群落最不稳定。
在封育边缘区共出现17种植物,其中刺沙蓬生态位宽度最大,达到0.810,远大于排在第2位的虫实Corisprmum chinganicum(0.605)和第3位的狗尾草Setaria viridis(0.504)。这类似于核心区,但在该区刺沙蓬并不是出现在每一个样方中,表明它比较适合在该区生长,相对其他物种其竞争力更强,更能适应该区的生长环境,是该区的优势物种。如图2所示,外围区生态位宽度平均值大于核心区,但方差小于核心区,大于边缘区,说明外围区生态位宽度的分布较核心区更为均匀,植被群落稳定性较核心区强,较边缘区弱。
封育外围区内共出现16种植物,少于核心区
(22种)和外围区(17种),其中生态位宽度最大的是丝叶山苦荬(0.742)、阿尔泰狗哇花Heteropappus altaicus(0.588)、黑沙蒿Artemisia ordosica Krasch(0.573)和刺沙蓬(0.558)。可以看出,丝叶山苦荬的生态位宽度明显大于其他物种,而其他3个物种的生态位宽度非常接近,这说明丝叶山苦荬在群落中生态适应范围很大,能很好的适应该区的资源环境,这与其适应干旱能力强的生物学特性有关。边缘区生态位宽度平均值最大,但方差最小(图2),这表明相对于边缘区和核心区,其他各主要建群物种的生态位宽度分布更为平均。这些说明在边缘区出现的植物对资源和空间的利用能力相对比较接近,物种之间的关系更为协调、平衡,其群落稳定性也就最强。
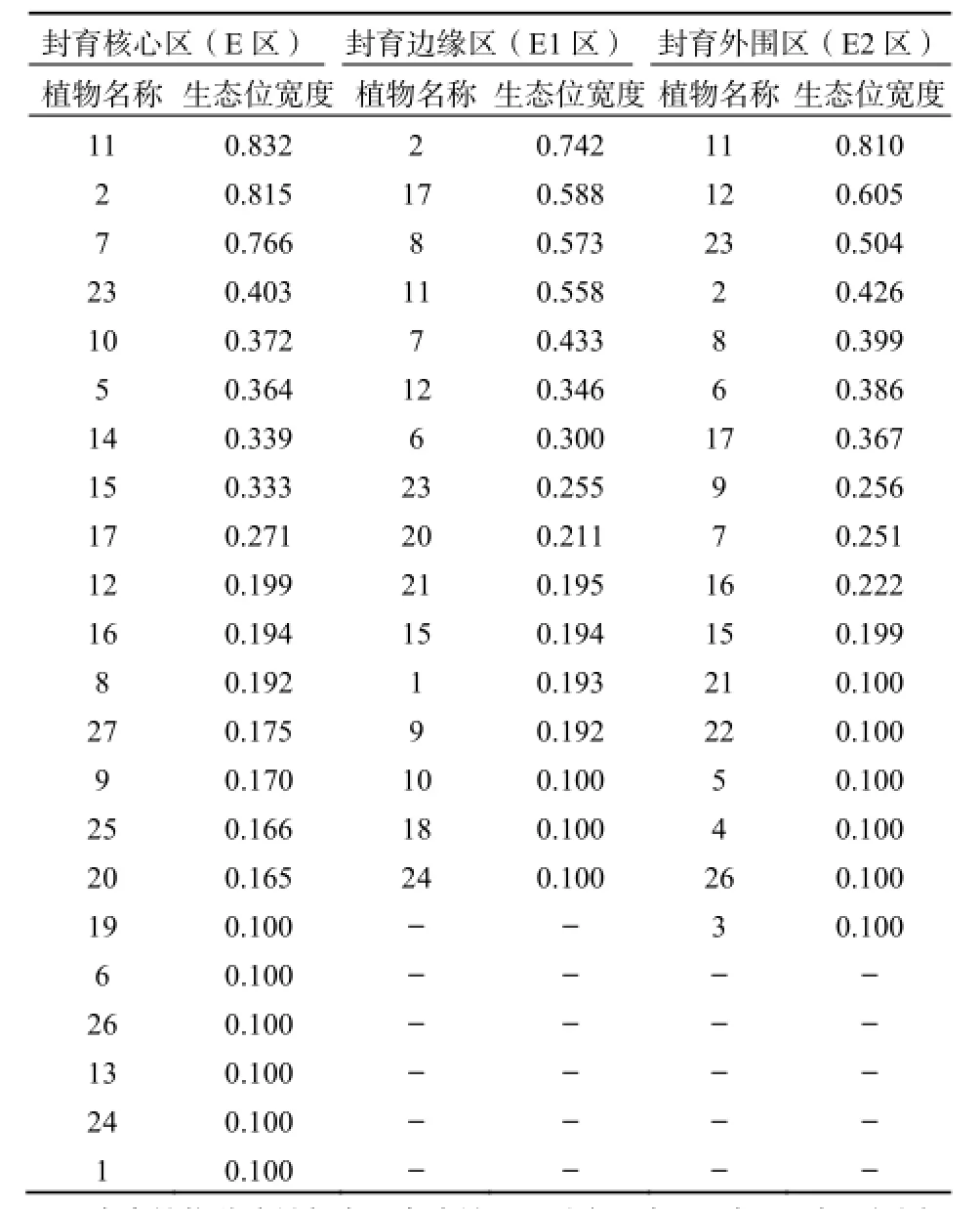
表2 2013年人工封育区植物生态位宽度Table 2 The niche breadth of plant in artificial fencing area in 2013
通过以上对封育区内3种不同处理方式中出现的植物种群生态位宽度的分析可以得出,其群落稳定性表现为E1区>E2区>E区。上述现象出现的主要原因是封育核心区自1991年以来长达32年的围栏封育使该区受人类活动的干扰很小,在土壤表层形成生物结皮,尽管生物结皮对荒漠地区的植被演替具有重要意义,有利于改善土壤理化性质(高广磊等,2014),有利于提高群落内的生物多样性(杨晓晖等,2005),但由于生物结皮的形成抑制了降雨向土壤深层的入渗,降低了土壤内含水量,因此这并不利于那些对环境适应能力弱的植物种群的生长。而边缘区和外围区自2002年开始封育,且它们的群落稳定性都比核心区强,这表明长期的完全封育并不利于封育区内植物群落的稳定。尽管外围区和边缘区从同1年开始封育,但因外围区不是完全封育,其内部仍受一些人类活动的影响,这导致植物生长状况与边缘区不同,其群落稳定性不如边缘区,这说明短期的完全封育比不完全封育更有利于植被群落的稳定。
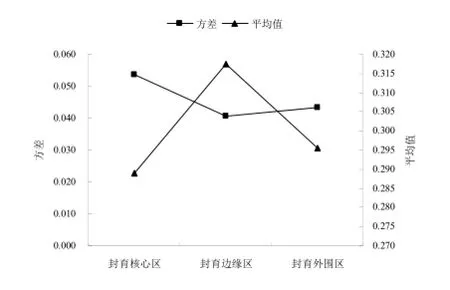
图2 不同处理方式生态位宽度方差和平均值对照Fig.2 The variance and average value of niche width in different ways
3.2 生态位重叠分析
生态位重叠反映的是物种之间对资源利用的相似程度和竞争关系(Spies,1990)。由于环境中的资源是有限的,所以生态位重叠高的2个物种之间存在着激烈的竞争,这种竞争最终会导致生态位重叠的降低。封育核心区、封育边缘区、封育外围区的生态位重叠计测结果,分别见表3、表4、表5。
封育核心区生态位重叠最大值出现在草地风毛菊Saussurea amara和臭蒿Artemisia hedinii、苦豆子Sophora alopecuroides L和臭蒿、沙芦草Agropyron mongolicum Keng和臭蒿之间,都是0.98,这4种植物生态位宽度值都非常小,最大的是臭蒿(0.165),其余3种均为的0.100,而核心区生态位宽度值最大的刺沙蓬和丝叶山苦荬,它们与其他物种之间的重叠度都不高,最高值出现在刺沙蓬与角蒿之间,重叠度为0.89(表3)。说明生态位宽度窄的物种之间可能有较高的生态位重叠,反之则较低。
封育边缘区生态位重叠最大的三对是达乌里胡枝子Lespedeza davurica和草木樨状黄芪Astragalus melilotoides、达乌里胡枝子和北拟云香Haplophyllum dauricum、中华隐子草Cleistogenes chinensis和刺沙蓬,分别为0.82、0.81、0.81,这些植物中刺沙蓬的生态位宽度最大是0.558,生态位宽度值在该区出现的所有植物中排名第4,而其他几个物种都非常低,该区中生态位宽度最大的丝叶山苦荬与其他物种之间的生态位重叠度大多数
都不高,最高值出现在与刺沙蓬之间(0.74),这仅比最高值小一点(表4)。由此进一步分析得出:较高的生态位重叠也可能出现在生态位宽的物种与生态位窄的物种之间。
封育外围区与核心区、边缘区相似,生态位重叠最大值并没有出现在生态位宽度最大的几种物种之间,而是出现在生态位宽度较窄的草木樨状黄芪和冰草Agropyron cristatum之间(0.99),而生态位宽度最大3个物种与其他物种的生态位重叠基本都不高,最大的是丝叶山苦荬与狗尾草、刺沙蓬和狗尾草之间,为0.71,远大于平均水平。这验证了通过表3和表4得出的结论。由以上分析可以判定:生态位宽度与生态位重叠之间并没有明确的联系。生态位宽度窄的物种之间可能有较高的生态位重叠,反之,则较低,但并不是完全如此。

表3 封育核心区(E区)植物生态位重叠Table 3 The niche overlaps of plant in E plot
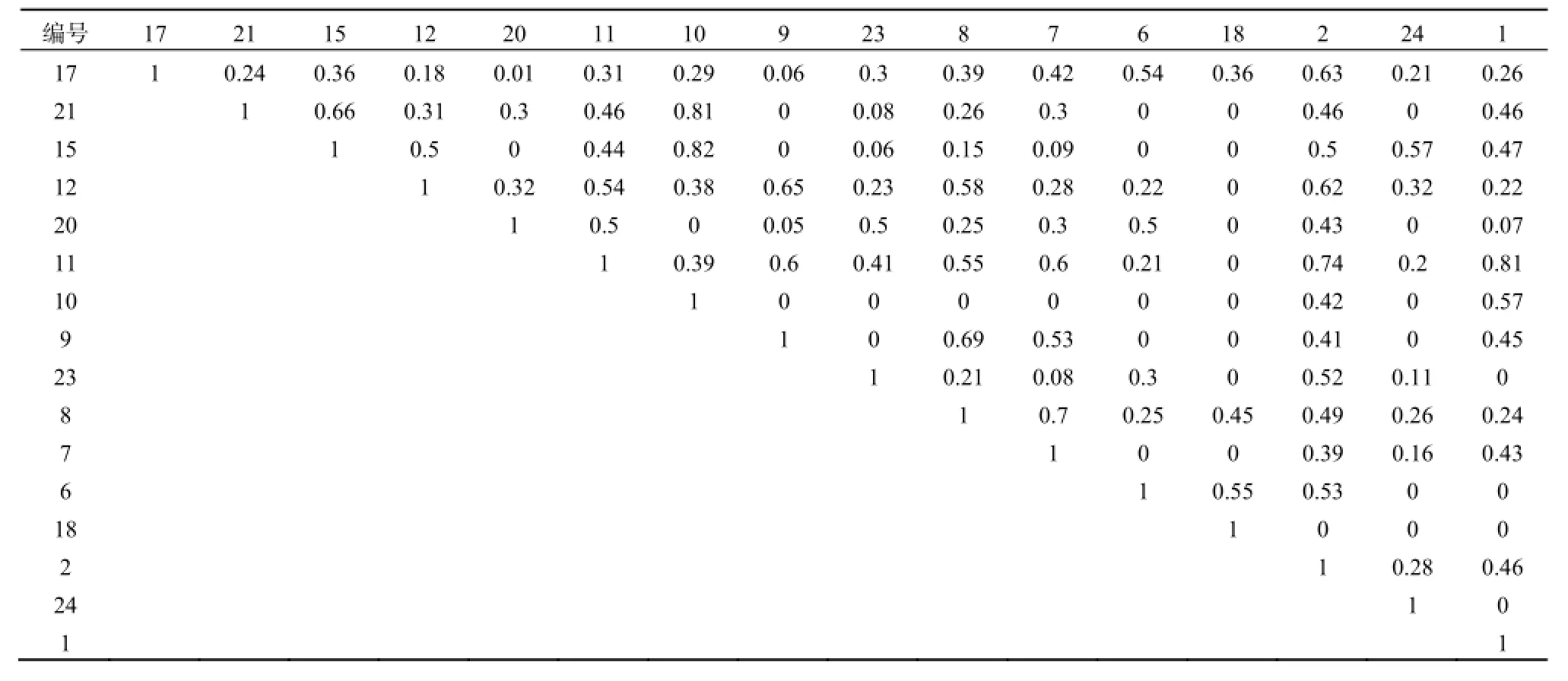
表4 封育边缘区(E1区)植物生态位重叠Table 4 The niche overlaps of plant in E1 plot
4 结论与讨论
4.1 结论
1)生态位宽度反映的是物种或种群对环境适
应的状况或对资源利用的程度(李德志等,2006),生态位宽度越大表明物种对资源的利用能力越大,对环境的适应能力越强。封育核心区和外围区生态位宽度最大的都是刺沙蓬,分别为0.832和0.810,而封育边缘区生态位宽度最大的是丝叶山苦荬,为0.742。刺沙蓬和丝叶山苦荬的生物学特性决定了它们能够很好的适应封育区的环境,并很好的利用封育区的资源,结果是它们的生态位宽度明显高于其他物种。
2)人工封育是退化草场地区一种有效的促进植被恢复的措施(李永宏,1995),在我国的草原恢复中起到了重要的作用,并得到了广泛的应用。对三区内不同植物种群的生态位宽度及其分布的分析结果显示,三区的群落稳定性表现为E1区>E2区>E区;长期的完全封育并不利于封育区内植物群落的稳定;而短期的完全封育措施比不完全封育措施更有利于植被群落的稳定。
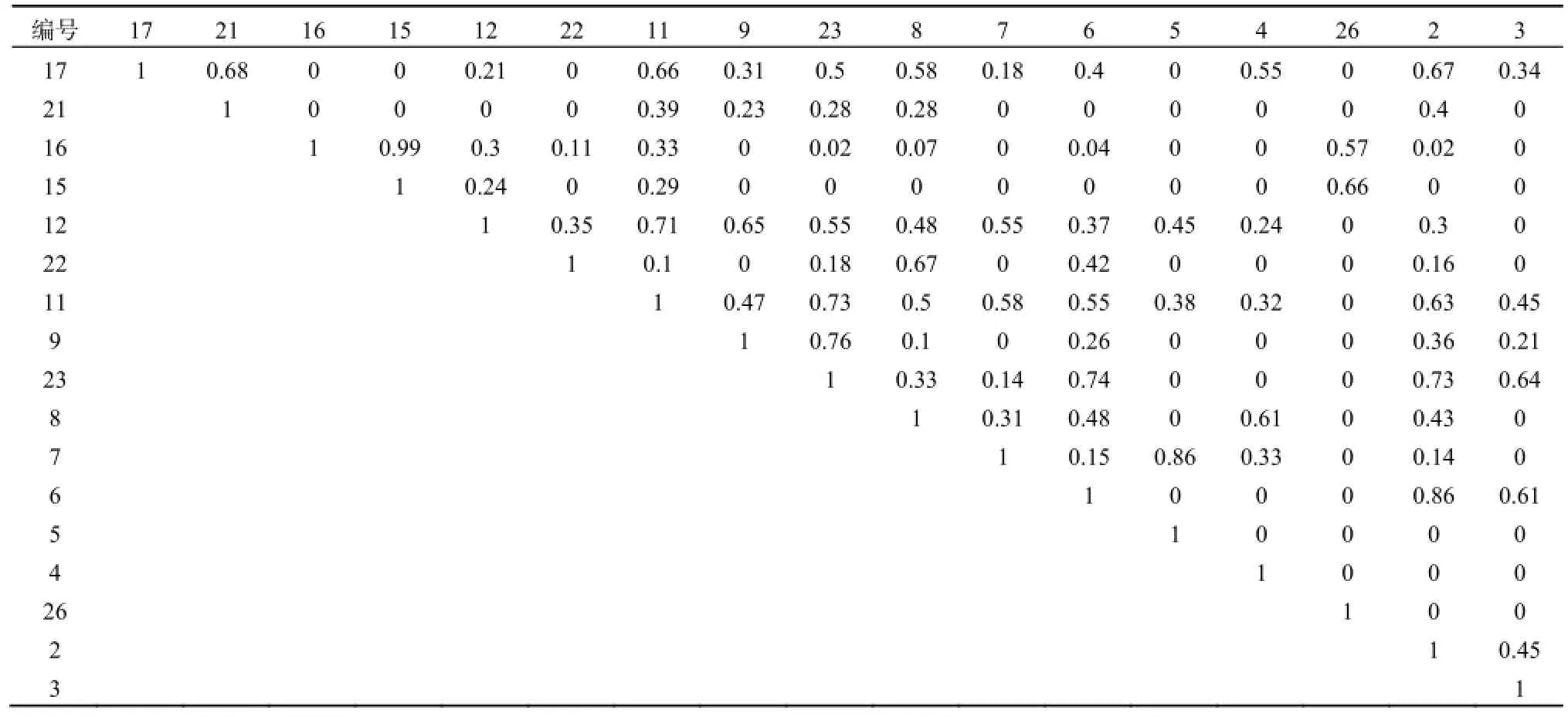
表5 封育外围区(E2区)植物生态位重叠Table 5 The niche overlaps of plant in E2 plot
3)生态位重叠是指不同物种之间生态位重叠或共有生态位空间的现象,即两个或两个以上的物种分享或竞争资源位或资源状态的现象(Spies,1990)。通过计算三区内不同物种之间的生态位重叠得出,E区生态位重叠最大值出现在草地风毛菊、苦豆子、沙芦草与臭蒿之间,都是0.98。E1区生态位重叠最大值是达乌里胡枝子和草木樨状黄芪之间(0.82)。E2区生态位重叠最大的一对是草木樨状黄芪和冰草(0.99)。本文相关分析表明:生态位宽度与生态位重叠之间并没有明确的联系。生态位宽度窄的物种之间可能有较高的生态位重叠,反之,则较低,但并不是完全如此。
4.2 讨论
人工封育是草场恢复和重建的一种主要的措施(Turner,1990),因其简单易行、投资少、见效快和便于推广的特性在各国得到广泛采用,且效果显著。尤其是在年降水量少、生态环境脆弱的干旱、半干旱草原地区,在封育初期,因围栏避免了牲畜对草场的破坏,确实有利于植被得到快速恢复(Yagil等,2002)。但经过一定时间的封育后,因地表枯枝落叶的增加,土壤表层结皮厚度增加,这抑制了降雨的渗透,使得植物可利用水分减少,导致植物生长受到抑制,使得部分植物退化,草原植物多样性降低(张克斌等,2008)。这表明人工封育并不是时间越长越好,当封育超过一定年限后,植物的生长会受到抑制,群落稳定性降低。因此,建议当地政府合理的调整草场管理模式,采用不完全封育(即封育和利用相结合)的措施经过一定时间的封育后,对草场进行适当的放牧或者刈割。在放牧过程中牲畜的活动破坏了结皮,加速降雨的入渗,这能促进植物的生长,提高草场的生产力,而适当的刈割能促进植物的再生长,加速植物群落的循环(程积民和邹厚远,1998)。尽管封育过程中对草场的利用能促进草场植物的更新,但注意利用要适时、适度,在什么时间、用什么强度进行利用才能保持草场的生产力需要进一步的调查研究,这样才能避免因人为因素再次造成草场退化。因各地自然条件不同,如何根据当地实际情况制定合理的封育措施,也将是今后研究的重点课题。
CAUSE G F. 1934. The Struggle for Existence[M]. Baltimore: William&
Wilkins, 19-20.
CURTIS J T, MCLNTOSH R P. 1951. An Upland Forest Continuum in the Prairie-forest Border Region of Wisconsin[J]. Ecology, 32: 476-496.
ELTON C. 1927. Animal Ecology[M]. London: Sidgewick and Jackson: 63-68.
GRINELL J. 1917. The Niche Relationship of the California Thrasher[J]. The Auk, 34: 427-433.
KREBS C J. 1978. Ecology: The Experimental Analysis of Distribution and Abundance [M]. New York: Fairfield Graphics: 225-228.
LEVINS R. 1968. Evolution in Changing Environments: Some Theoretical Explorations [M]. USA: Princeton University Press.
ODUM E D. 1971. Fundamentals of Ecology[M]. Philadelphia, USA: WB. Saunders Co.
PIANKA E R. 1973. The Structure of Lizard Communities[J]. Annual Review of Ecology and Systematics, 4: 53-74.
SPIES T A. 1990. Canopy Gaps in Douglas-fir Forests of the Cascade Mountains [J]. Can J For Res, 20(3): 649-658.
TURNER R M. 1990. Long-term Vegetation Change at a Fully Protected Sonoran Desert Site[J]. Ecology, 7: 464-477.
YAGIL O, AVI P, Jaime K. 2002. Grazing Effect on Diversity of Annual Plant Communities in a Semi-arid Rangeland: Interactions with Small-scale Spatial and Temporal Variation in Primary Productivity[J]. Journal of Ecology, 90: 936-946.
程积民, 邹厚远. 1998. 封育刈割放牧对草地植被的影响[J]. 水土保持研究, (1): 36-54.
高广磊, 丁国栋, 赵媛媛, 等. 2014. 生物结皮发育对毛乌素沙地土壤粒度特征的影响[J]. 农业机械学报, 45(1): 115-120.
李德志, 石强, 臧润国, 等. 2006. 物种或种群生态位宽度与生态位重叠的计测模型[J]. 林业科学, 42(7): 95-103.
李军玲, 张金屯, 郭逍宇. 2003. 关帝山亚高山灌丛草甸群落优势种群的生态位研究[J]. 西北植物学报, 23(12): 2081-2088.
李永宏. 1995. 内蒙古典型草原地带退化草原的恢复动态[J]. 生物多样性, 3(3): 125-130.
尚玉昌. 1988. 现代生态学中的生态位理论[J]. 生态学进展, 5(2): 77-84.
汪建华, 黄林, 李旭光. 2001. 重庆四面山常绿阔叶林主要乔木种群生态位特征[J]. 重庆三峡学院学报, 17(4): 76-80.
王刚, 张大勇. 1991. 亚高山草甸弃耕地植物群落演替的数量研究IV.组分种生态位分析[J]. 草地学报, 1(1): 93-99.
王刚, 赵松岭, 张鹏云, 等. 1984. 关于生态位定义的探讨及生态位重叠计测公式改进的研究[J]. 生态学报, 4(2): 119-127.
王育松, 上官铁梁. 2010. 关于重要值计算方法的若干问题[J]. 山西大学学报: 自然科学版, 33(2): 312-316.
魏志琴, 李旭光, 郝云庆. 2004. 珍稀濒危植物群落主要种群生态位特征研究[J]. 西南农业大学学报: 自然科学版, 26(1): 1-4.
杨晓晖, 张克斌, 侯瑞萍, 等. 2005. 半干旱沙地封育草场的植被变化及其与土壤因子间的关系[J]. 生态学报, 25(12): 3212-3219.
张克斌, 卢晓杰, 李瑞. 2008. 北方农牧交错带沙地生物结皮研究[J]. 干旱区资源与环境, (4): 147-151.
张润涛, 郭健. 2008. 浅谈生态位理论的意义及应用[J]. 林业科技情报, 40(4): 64-66.
朱春全. 1993. 生态位理论及其在森林生态学研究中的应用[J]. 生态学杂志, 12(4): 41-46.
Vegetation niche of enclosed grassland in semi-arid area——Taking Yanchi of Ningxia as an example
LIU Jiankang1, ZHANG Kebin2, WANG Lili3, WANG Zhishu3, WANG Guanqi3
College of Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China
Artificial enclosure is one of the major restoration measures in degraded grassland. In order to find the vegetation niche of enclosed desertification rangeland in semi-arid area, this paper studied the three different treatments (long-term complete enclosure; short-term complete enclosure; short-term incomplete enclosure) in enclosure area which located in Yanchi County of Ningxia. The enclosure area has been divided into the long-term complete enclosure region (E zone), short-term complete enclosure region (E1 zone) and short-term incomplete enclosure region (E2 zone). By analyzing the Levins niche breadth index and Pianka niche overlapping index of 27 kinds of plants in this area, reveal the variation law of plants niche in the arid desert environment, and provide the basis for further analysis of community stability in this region. The results showed that: 1) The niche breadth of Salsola ruthenica lljin was the biggest in both E zone and E2 zone, which were respectively 0.832 and 0.810. While the dominate species in E1 zone was Lxeris chinensis var.graminifolia, the niche breadth was 0.742. It was their own biological characteristics that decided they can adapt the environment very well in enclosed area, and use the resources of enclosure region very well. The result was that their niche breadth was significantly higher than other species. 2) The analysis of the different plants' niche breadth in three areas revealed that: in terms of community stability, E2 zone was worse than E1 zone, and better than E zone. The long-term fencing enclosure was unfavorable for community stability. While short-term complete enclosure was more beneficial for community stability than incomplete enclosure. 3) By analyzing niche overlapping between different plants species in three areas drawn, the maximum niche overlapping value in E area appeared between Saussurea amara, Sophora alopecuroides L, Agropyron mongolicum Keng and Artemisia hedinii, both were 0.98. The maximum value in E1 area was between Lespedeza davurica and Astragalus melilotoides (0.82), in E2 area was between Astragalus melilotoides and Agropyron cristatum(0.99). Correlation analysis showed that there was no clear linear relationship between niche breadth and niche overlapping. There may have narrow niche overlapping between narrow niche breadths species, the opposite is low, but not exactly so.
enclosure; grassland vegetation; niche breadth; niche overlapping; semi arid region; community stability
Q948
A
1674-5906(2014)05-0762-07
国家自然科学基金项目(30771764);国家林业局“宁夏盐池荒漠化定位监测项目”
刘建康(1988年生),男,硕士研究生,研究方向为荒漠化防治与检测。E-mail:liujiankang1989@126.com
*通信作者:张克斌(1957年生),男,教授,博士生导师,研究方向为荒漠化防治与检测。E-mail:ctccd@126.com
2014-03-29
刘建康,张克斌,王黎黎,王志述,王冠琪. 半干旱区人工封育草场植被群落生态位研究——以宁夏盐池县长期定位监测点为例[J]. 生态环境学报, 2014, 23(5): 762-768.
LIU Jiankang, ZHANG Kebin, WANG Lili, WANG Zhishu, WANG Guanqi. Vegetation niche of enclosed grassland in semi-arid area——Taking Yanchi of Ningxia as an example [J]. Ecology and Environmental Sciences, 2014, 23(5): 762-768.
