紫茎泽兰多糖的免疫调节活性研究
2014-02-15万春燕周绪斌
万春燕,周绪斌
(1.中国农业大学,北京 100093;2.新疆天康畜牧生物技术股份有限公司,新疆 乌鲁木齐 830032)
1 紫茎泽兰多糖的提取
取100 g阴凉处干燥的紫茎泽兰叶片,将叶片剪碎,辗成粉,40目过滤筛过滤。再将其溶于1 000 mL蒸馏水中,于超声波提取装置中提取40 min,收集上清液,沉淀物再悬浮于1000 mL蒸馏水,再次通过超声处理提取30 min,将得到的上清液与前一次所得合并,最后在旋转蒸发器中将水相蒸发至干,得到的残留物溶解在蒸馏水中,并在4℃下冷冻保存。
2 紫茎泽兰多糖的纯化制备
将萃取液在3 000 g/min离心25 min,80 ℃下进行浓缩8 h,以制备多糖,将上清液用Sevag法脱蛋白。必要时可用紫外分光光度计测 250~280 nm没有吸收峰方可确定除净了蛋白。将上层清液用2 mol/L 氢氧化钠调至pH=7,加热回流用1%活性炭脱色。
取除杂蛋白后的溶液10 mL,装入截留分子量为8 000~15 000的透析袋中透析脱盐及小分子杂质,透析液离心(3 000 r/min,10 min),上清于80 ℃水浴浓缩至原体积的1/3。然后加入4倍体积95%乙醇(v/v)沉淀24 h后,离心(3 000 r/min,15min),沉淀用无水乙醇洗涤2次,乙醚洗涤1次,真空干燥得紫茎泽兰多糖样品。
以葡萄糖作标准曲线,用苯酚-浓硫酸分光光度法测醇沉淀物中的EAP糖含量。
3 紫茎泽兰多糖对人源细胞和鼠源细胞的免疫调节作用
3.1 多糖的处理
1)将提取的紫茎泽兰多糖溶于蒸馏水中,经0.45 μm和0.22μm的过滤器过滤,用细胞培养液稀释;
2)将T25细胞培养瓶培养的人源(A549)细胞和鼠源(RAW264.7)细胞经胰酶消化后加入细胞培养液使其最终体积为20 mL,向6孔培养液中加入2 mL/孔,同时加入紫茎泽兰多糖溶液200 μL/孔,使其最终浓度为50,100,200,400μg/mL,另外设脂多糖(LPS)组,每个梯度3个重复。空白对照加入200 μL相应的磷酸缓冲(PBS)液。
3)培养12 h,24 h,36 h每个浓度取3个样品,提取细胞总RNA,采用荧光定量PCR方法检测紫茎泽兰多糖的免疫相关细胞因子IL-6,TNF-α,IFN-β,IFN-γ等。
3.2 采用Trizol法提取细胞总RNA,采用随机引物合成cDNA
3.3 荧光定量PCR
用β-actin作为内参。获得的实时定量PCR数据用2-ΔΔCT方法分析相关基因的表达。
3.4 数据分析
使用GraphPad Prism V5.01作图并进行统计学分析。P<0.05为差异显著,在图中用“*”表示。
4 结果
4.1 紫茎泽兰多糖对人源细胞(A549)的免疫调节作用
4.1.1 不同浓度的EAP对A549细胞的影响
实验选取的紫茎泽兰多糖浓度分别为50μg/mL、100μg/mL、200 μg/mL和 400μg/mL, 分 别 作 用 于A549细胞,提取RNA,反转录为cDNA,用相对荧光定量PCR检测细胞因子IL-6、TNF-α、IFN-β和IFN-γ的mRNA表达情况。结果表明在200μg/mL时4种细胞因子的表达量都较高(P<0.05)。
EAP终浓度分别为50 μg/mL、100μg/mL、200μg/mL和400μg/mL,LPS对照的浓度为50 ng/mL,PBS组为阴性对照。从图1中可以看出,IL-6、TNF-α、IFN-β和IFN-γ在EAP浓度为200 μg/mL时其mRNA的表达都显著提高(P<0.05)。EAP浓度200μg/mL与其他浓度相比提高效果最明显。
4.1.2 不同时间点EAP对A549细胞的影响
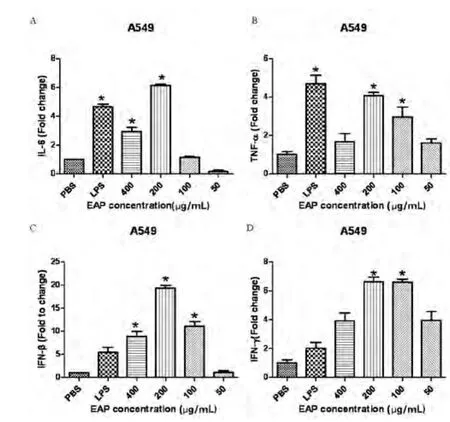
图1 采用相对荧光定量PCR方法测定不同浓度的EAP对A549细胞因子表达的影响
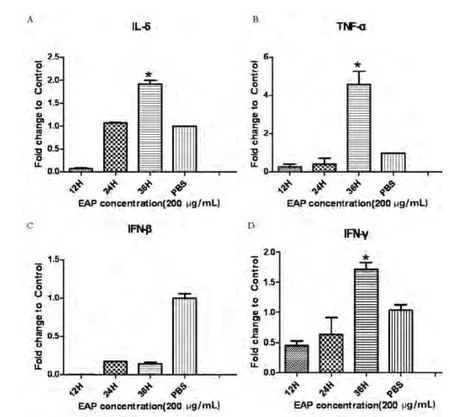
图2 采用相对荧光定量PCR方法测定不同时间点EAP对A549细胞的影响
实验选取了紫茎泽兰多糖浓度为200 μg/mL,分别作用时间12 h、24 h和36 h于A549细胞,提取RNA,反转录为cDNA,用相对荧光定量PCR检测细胞因子IL-6、TNF-α、IFN-β和IFN-γ的mRNA表达情况。结果表明EAP浓度200 μg/mL作用36 h,IL-6、TNF-α和IFN-γ的表达量都较高(P<0.05),IFN-β无统计学意义。
分别取EAP作用的3个时间点即12 h,24 h和36 h,测定不同时段细胞因子的表达情况,从实验所得结果来看,36 h时EAP的作用相对较好,IL-6,TNF-α和IFN-γ都得到了上调表达(P<0.05)。IFN-β在3个时间段的结果不显著(图2-2C)。
4.2 紫茎泽兰多糖对鼠源细胞的免疫调节作用
4.2.1 不同浓度的EAP对RAW264.7细胞的影响
实验选取了紫茎泽兰多糖(EAP)浓度分别为50μg/mL、100μg/mL、200μg/mL和 400μg/mL,分别作用于RAW264.7细胞,提取RNA,反转录为cDNA,用相对荧光定量PCR检测细胞因子IL-6、TNF-α、IFN-β和IFN-γ的mRNA表达情况。结果表明在200μg/mL时4种细胞因子的表达量都较高(P<0.05)。
实验分别设定了紫茎泽兰多糖的4个浓度梯度,50μg/mL、100μg/mL,200μg/mL和400μg/mL,另外设PBS空 白 对 照 组。IL-6、TNF-α、IFN-β 和IFN-γ 都 在EAP浓度为200μg/mL的时候表达量最高。200μg/mL的紫茎泽兰多糖与PBS组相比差异显著(P<0.05)。
4.2.2 不同时间点EAP对RAW264.7细胞的影响

图3 采用相对荧光定量PCR方法测定不同浓度的EAP对RAW264.7细胞的影响
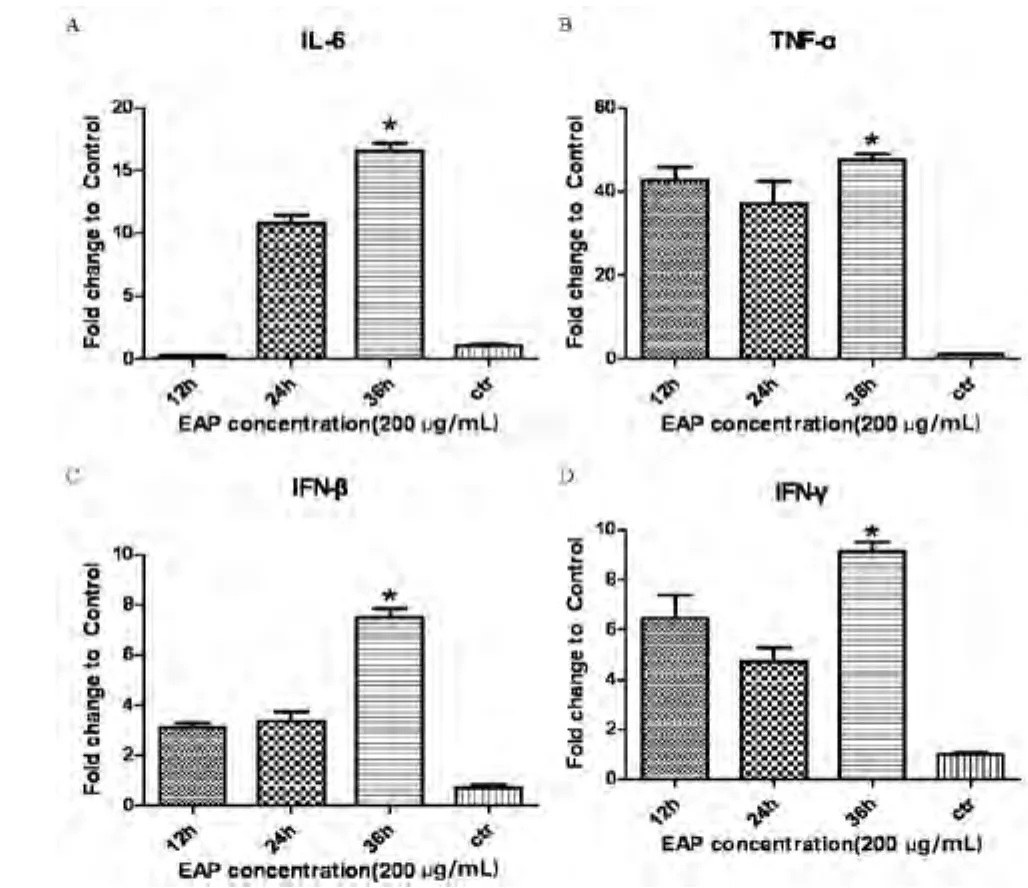
图4 采用相对荧光定量PCR方法测定不同时间点EAP对RAW264.7细胞的影响

图5 测定不同浓度EAP对小鼠巨噬细胞RAW264.7的吞噬李氏杆菌的刺激作用
实验选取的紫茎泽兰多糖浓度为200μg/mL,分别作用时间12 h、24 h和36 h于A549细胞,提取RNA, 反转录为cDNA,用相对荧光定量PCR检测细胞因子IL-6、T N F-α、I F N-β和I F N-γ的mRNA表达情况。结果表明EAP浓度200μg/mL作用36 h,IL-6、TNF-α和IFN-γ的表达量都较高(P<0.05),IFN-β结果不显著。
从这4个因子(IL-6、TNF-α、IFN-β和IFN-γ)的表达情况来看,36 h与12 h和24 h相比,更具有优势,与对照组相比,差异显著(P<0.05)。
4.3 巨噬细胞的吞噬能力测定
实验测定了EAP浓度分别为100μg/mL、200μg/mL和400μg/mL时RAW264.7细胞对李氏杆菌的吞噬作用,用LPS作阳性对照,结果表明200μg/mL的EAP具有促进RAW264.7细胞吞噬细菌的作用。
分别选取了100μg/mL,200μg/mL,400μg/mL的EAP浓 度 梯 度,Lipopolysaccharides(LPS)为 50 ng/mL。以LPS为阳性对照,PBS为阴性对照,EAP作用36 h后,200μg/mL浓度的紫茎泽兰多糖与阴性对照组相比吞噬能力较好(P<0.05)。
5 分析与讨论
目前关于植物多糖的抗氧化能力和增强宿主免疫活性的作用方面的研究还较少。本试验通过对体外人源细胞A549细胞和鼠源细胞RAW264.7的细胞因子的测定发现,在EAP浓度为200μg/mL,作用时间为36 h时表达水平较高。EAP对A549细胞和RAW264.7细胞的免疫调节活性与EAP的剂量成一定的依赖关系。EAP能显著提高这2种不同来源细胞 IL-6,TNF-α,IFN-β,IFN-γ 的 mRNA的表达水平。IL-6主要由巨噬细胞、成纤维细胞、B细胞、T细胞等多种细胞产生的,IL-6可以调节细胞的生长与分化,具有调节免疫应答、急性期反应、造血功能等,并在机体的抗感染免疫中发挥重要作用。许多抗原或非抗原物质可诱导IL-6的产生,如植物多糖和细胞因子等,TNF-α和IL-6可以互相诱导产生,互相进行调控。IL-6可通过核转录因子(NF-κB)介导炎症,NF-κB在激活上皮细胞、巨噬细胞和T细胞等细胞中的各种促炎症因子起着重要作用。TNF-α是由活化的单核细胞、巨噬细胞、T细胞等产生的能使肿瘤坏死的因子,脂多糖(LPS)、病毒、真菌、免疫复合物和其他非抗原物质均具有刺激TNF-α产生的作用。TNF-α具有抗病毒、细菌、激活T细胞,促进IL-6的产生和分泌的作用,并诱导炎症反应,促进主要组织相容性抗原Ⅱ类抗原(MHCⅡAg)的表达等,与宿主防御反应相关。TNF-α在浓度较低时,它主要作为内皮细胞和白细胞自分泌和旁分泌的调节物质,调节免疫反应,参与抗病毒、抗肿瘤作用等。干扰素(IFN)是动物机体细胞在诱导剂作用下产生的一组低分子量的具有抗病毒、抗肿瘤、免疫调节活性的糖蛋白。IFN-β具有免疫调节作用,它能促进大多数细胞MHCⅠ类抗原的表达,激活细胞毒性T淋巴细胞(CTL)和自然杀伤(NK)细胞,另外还具有一定的广谱抗病毒作用。T细胞受到刺激后可产生IFN-γ,目前认为,巨噬细胞活化因子(MAF)的主要活性存在于IFN-γ中,IFN-γ能诱导MHCⅡ类抗原的表达,协同TNF促进细胞杀伤病原微生物。紫茎泽兰多糖作为细胞体外刺激物,它能以一定的作用机制调节上皮细胞和巨噬细胞内IL-6、TNF-α、IFN-β、IFN-γ的表达,这对细胞的免疫反应具有一定的作用。
以往的研究表明巨噬细胞受到刺激后,能够产生细胞因子如IL-6和TNF-α。T细胞分泌的IL-6能刺激巨噬细胞的免疫反应。巨噬细胞分泌的TNF-α已确定是重要的宿主调节分子,与肿瘤坏死相关。紫茎泽兰多糖对RAW264.7细胞和A549细胞具有一定的免疫调节作用,这种结构的多糖可能与细胞表面受体有一定关联,从而导致免疫反应的发生,但这还需要进一步的研究加以证明。巨噬细胞在抵御宿主感染和破坏外来病原体和肿瘤细胞中发挥了重要作用。在哺乳动物中,吞噬是一个重要的防御机制,它能够使机体免受病原体的入侵,巨噬细胞、树突状细胞和粒细胞对凋亡细胞具有清除作用。巨噬细胞的非特异性防御功能可能是在吞噬过程中消除阶段通过激活溶酶体磷酸酶而发挥的,由于吞噬细胞的监管和效应细胞在免疫系统的作用,通过增强巨噬细胞功能,将适用于微生物感染和癌症的治疗领域。
研究结果表明,EAP可以刺激A549细胞和RAW264.7细胞的免疫反应,可能是由于提取方法,单糖单位,结构特征和分子质量等的综合作用。也可能是由于巨噬细胞膜上的受体(包括dectin-1,甘露糖受体,Toll样受体等),这些受体识别多糖将产生细胞增殖,引起一氧化氮(NO)、活性氧(ROS)的产生,细胞因子或者趋化因子的生产,增强吞噬作用。一旦免疫反应开始,巨噬细胞将使用NO、ROS或TNF-α攻击外来病原体和诱导细胞凋亡,然后吞噬或传递给抗原呈递细胞和自然杀伤细胞或细胞毒性T淋巴细胞来识别并消除它们。因此EAP被认为是新型植物多糖,可以用来激活免疫系统,迅速形成巨噬细胞作为主体的第1道防线。因此, EAP对于免疫佐剂的开发或免疫调节剂的研制作用甚大,有可能会成为一种广谱的抗病毒药物。
略
