川西高山森林生态系统林下生物量及其随林窗的变化特征
2014-02-09吴福忠杨万勤常晨辉
肖 洒,吴福忠,杨万勤,常晨辉,李 俊,王 滨,曹 艺
四川农业大学生态林业研究所,林业生态工程重点实验室,高山森林生态系统定位研究站,四川 成都 611130
川西高山森林生态系统林下生物量及其随林窗的变化特征
肖 洒,吴福忠,杨万勤*,常晨辉,李 俊,王 滨,曹 艺
四川农业大学生态林业研究所,林业生态工程重点实验室,高山森林生态系统定位研究站,四川 成都 611130
作为森林生态系统的重要组成部分,林下植被及其残体的分布受到林冠层的影响,但迄今有关林窗对林下植被和残体生物量的影响尚无研究报道。于2013年8月2日至20日,以海拔3 600 m的川西岷江冷杉原始林林下植被为研究对象,根据区域内的坡向和林分组成等因素设置3个100 m×100 m的典型样地,调查其生物量及其随林窗的变化特征。在每个样地内选择3个大林窗,在林窗、林缘和林下分别设置3个20 m×20 m的样方,调查粗木质残体长度或高度、大小头直径、枯立木记录胸径、腐烂等级等;在林窗、林缘和林下分别设置3个5 m×5 m的样方,采用“收获法”收集样方内直径在2.5~10 cm之间的细木质残体和灌木生物量;在林窗、林缘和林下分别设置3个1 m×1 m的样方来调查凋落物储量和草本生物量;在1 m×1 m的样方内随机选择1个20 cm×20 cm的小样方来调查地被植物生物量。结果表明,(1)川西高山森林生态系统总生物量为72.75 t·hm-2,其中林下生物量为67.92 t·hm-2,占生态系统生物量的95.17%。活体植被以灌木为主,其生物量为9.81 t·hm-2;残体部分以粗木质残体为主,其储量为53.00 t·hm-2;(2)林窗对灌木、草本、地被植物的影响各不相同,且不同物种的灌木生物量表现出不同的分布规律;草本生物量表现出明显的“边缘效应”,在林缘显著高于林下;林窗和林缘的地被植物生物量相对较低;(3)粗木质残体储量从林下到林窗呈现减小的趋势,但总体储量仍然较大,林窗和林缘的细木质残体储量高于林下。这些结果为认识高山森林生态系统林下生物量及其格局,以及林窗在森林生态系统的重要作用提供了基础理论依据。
高山森林;林下植被;生物量;林窗
林下植被是森林林下灌木、草本和藤本植物等的统称,是森林生态系统的重要组成部分,在维持生态系统多样性、生态功能稳定性、等方面具有独特的功能和作用(杨昆和管东生, 2006; 刘凤娇和孙玉军, 2011; 黎燕琼等, 2010)。林下植被生物量的变化在一定程度上反映了森林植被演替、人类活动、自然干扰、气候变化等的影响(刘凤娇和孙玉军, 2011)。已有研究主要集中在林下灌木层和草本层(太立坤等, 2009; 王祖华等, 2010; 于立忠等, 2006),忽略了地被植物对维持近地面微环境的作用和水土涵养的能力,此外,凋落物层和木质残体在森林生态系统养分循环中起着重要的桥梁和纽带的作用(郭剑芬等, 2006; 彭少麟等, 2002; 杨万勤等, 2007; Harmon等, 1986)。林下植被生物量的变化受林龄和林分郁闭度的影响较大,高山森林生态系统林分结构以成熟林和过熟林为主,树木的正常死亡以及频繁的干扰促进了林窗的形成(吴庆贵等, 2013),从而影响了林分光照条件、水分条件、温度条件以及土壤性质(王微等, 2004; 陶建平等, 2006),最终使得林下生态系统的生物量产生差异,但缺乏必要的研究。
川西高山森林生态系统地处青藏高原东缘和长江上游地区,在在区域气候调节、涵养水源和生物多样性保护保育等方面具有不可替代的作用和地位,是我国典型的生态脆弱带和全球气候变化的敏感区(Yang等, 2005; Liu等, 2014)。研究表明,高纬度和高海拔地区对全球气候变化十分敏感,受低温限制、频繁的山地灾害导致的浅薄土层以及频繁的强风、暴雪、冰雹、雷击等自然灾害影响,高山森林林窗干扰频繁(Zhang等, 2005; 吴庆贵等, 2013)。林下植被生物量大小分配差异与森林类型特征有关,并与森林的发育阶段紧密相连,森林类型在生长发育过程中的差异导致林内环境的变换,影响林内光照、水分及土壤理化性质、林内温湿度等自然环境,林下植被差异较大。因此,在本团队前期的研究结果的基础上,以川西高山森林林下生态系统为研究对象,进一步研究其生物量对林窗格局的响应,为进一步研究川西高山森林生态系统物质循环和能量流动提供基础资料,为深入理解森林生态系统过程以及可持续管理森林生态系统提供科学依据。

表1 研究区土壤理化性质Table 1 Soil physical and chemical properties in study area (mean±SD, n=5)
1 研究区域与研究方法
1.1 研究区域概况
本研究区域位于四川省阿坝州米亚罗自然保护区毕棚沟(E102°53′~102°57′,N31°14′~31°19′),地处青藏高原-四川盆地的过渡地带,四姑娘山北麓,区域总面积180 km2。区域气候属丹巴-松潘半湿润气候,随着海拔上升,气候呈现出暖温带、温带、寒温带、亚寒带和冰冻带的垂直分布规律。依据在该区3个地点连续2年的气象记录,区域内的年平均温度为2~4 ℃,最高气温23.7 ℃,最低温度为-18.1 ℃。年降雨量随着海拔高度的变化而变化,大约为801~850 mm,绝大多数降雨集中在5~8月。受季风的影响,区域内干湿季节差异显著:干季日照强、降水少、气候寒冷、空气干燥;湿季日照少、降雨多、气候温暖、多云雾。冬季较低的气温导致土壤的季节性冻融,土壤季节性冻结期长达4~6月,冻融明显。土壤发育缓慢,主要为雏形土(Cambisols)和新成土(Primosols),石砾、石块含量多,地表凋落物较厚。植被垂直成带明显,其类型和生境随海拔及坡向而分异。主要森林植被随海拔分异为岷江冷杉(Abies faxoniana)原始林、岷江冷杉和红桦(Betula albosinensis)混交林、岷江冷杉次生林。研究区域内土壤基本理化性质见表1。
1.2 研究方法
2013年8月2日至20日,以研究区域内海拔3600 m左右的岷江冷杉原始林林下植被为研究对象,根据区域内的坡向和林分组成等因素设置3个 100 m×100 m的典型样地,对灌木、草本、地被植物、凋落物和木质残体展开调查和采样。
1.2.1 乔木生物量测定
在100 m×100 m的样地内,按照“五点法”设置5个20 m×20 m的样方,分别记录样地内乔木的树种、胸径、树高,然后通过平均实验形数法计算乔木的蓄积(林昌庚, 1964; 吕勇等, 2009),再通过生物量-蓄积模型计算乔木生物量(黄从德等, 2009)。
1.2.2 林下灌木生物量的测定
在100 m×100 m样地内按照林窗、林缘、林下分别随机设置3个5 m×5 m的样方。在5 m×5 m的小样方内,逐一测量灌丛的冠幅和高度,记录每一株灌木的基径和高度,根据杨昆与管东生的林下植被生物量模型估算其生物量(杨昆和管东生, 2007)。
1.2.3 草本和地表植物生物量的测定
草本和地表植物的研究方法采用直接收获的方法,在100 m×100 m样地内按照林窗、林缘、林下分别随机设置3个1 m×1 m的样方,将样方里面的草本全部收获称量鲜重以后带回实验室;在1 m×1 m的样方内随机选择1个20 cm×20 cm的小样方,收集全部地表植物称量鲜重以后带回实验室,置于105 ℃的烘箱中杀青30 min,然后将烘箱温度调至65 ℃烘干至恒重。
1.2.4 凋落物储量的测定
在上述1 m×1 m的样方内,把直径<2.5 cm的非木质残体全部收集带回实验室,包括凋落叶、细枝及花、果实等细小凋落物,称量鲜重以后带回实验室,在105 ℃烘干至恒重。
1.2.5 细木质残体储量的测定
在上述5 m×5 m的样方内,收集样方内直径在2.5~10 cm之间的细木质残体,称量鲜重后,取3份混合样,在实验室将所取得的混合样在105 ℃条件下烘干至恒重。
1.2.6 粗木质残体储量的测定
在每块样地,选择3个大林窗,在每个林窗内设置1个20 m×20 m的样方,同时在林缘和林下分别设置3个20 m×20 m的样方,每个样方之间的间距均超过5 m,每块样地包括9个样方。在每个20 m×20 m的样方内逐一记录直径≥10 cm的粗木质残体(倒木、枯立木、大枯枝、根桩),记录内容包括长度或高度、大小头直径、枯立木记录胸径、腐烂等级等,对于长度超出样方大小的粗木质残体,只记录其在样方内的部分。对Ⅰ、Ⅱ、Ⅲ腐烂等级的粗木质残体,取圆盘带回实验室,Ⅳ、Ⅴ腐烂等级的粗木质残体用封口袋直接采样。采用排水法(张利敏等, 2010; 张修玉等, 2009)测定其体积(V),烘干后得到其重量G,粗木质残体的密度即为G与V的比值,再用密度分别乘以样地内CWD的体积,即为CWD的质量。
1.3 数据处理与统计分析
采用Microsoft Excel 2007软件进行数据的整理,SPSS 20.0软件进行统计分析,采用单因素方差分析(one-way ANOVA)比较地被层生物量在林窗、林缘、林下之间的差异。

表2 高山森林生态系统生物量及其比例Table 2 Alpine forest ecosystem biomass and its proportion t·hm-2

表3 灌木生物量及其分布Table 3 Shrub biomass and its distribution kg·hm-2
2 结果与分析
2.1 高山森林生态系统生物量分配
川西高山森林生态系统总生物量为72.75 t·hm-2,林下生物量为67.92 t·hm-2(表2),其中木质残体储量最大,占72.85%,灌木生物量次之,占13.48%,其他层片的生物量或储量表现为地被植物>凋落物>草本。
川西高山森林生态系统总生物量为72.75t·hm-2,远低于西藏色季拉山原始冷杉林生态系统生物量(方江平和项文化, 2008),也低于贡嘎山峨眉冷杉林(Abies fabri)(罗辑, 2000)。和中国平均89.09 t·hm-2相比略低,和四川省105.04 t·hm-2的平均森林生物量相比,川西高山森林生物量低于四川省平均森林生物量。研究区域呈现出乔木生物量比例降低,灌木和木质残体生物量及储量比例升高的趋势(程小琴等, 2012; 田大伦等, 2014),这是因为研究区域已处于群落演替的关键期(吴庆贵等, 2013),森林类型为成熟林向过熟林过渡时期,大量林窗的形成不但减少了乔木的生物量、增加了木质残体的储量,同时也使得灌木层的比例相应身高。
2.2 灌木生物量随林窗的变化
相对于林缘,林窗和林下物种丰富度较高,优势物种明显,林窗和林下均以冰川茶藨子和蔷薇为优势种,灌木生物量及其分布见表3。总体来说,灌木生物量呈现出林窗>林缘>林下的趋势。林下灌木是森林生态系统中的一个重要组成部分,在整个森林生态系统营养元素的积累、循环、维持生物多样性,尤其是在促使林分养分步入良性生物循环和对森林林分未来的结构和组成以及受干扰后森林演替等方面具有不可忽视的作用(杨昆和管东生, 2007; Taylor等, 2004; Oakley等, 2006)。林窗的形成不仅对森林的环境特征有着重要影响(崔宁洁等, 2014),更会影响林下灌木生物量的积累和分配。林窗形成以后,引起光照强度、大气温度和土壤含水量等环境因子的改变,不同物种对环境因子改变的适应性不一样,使得不同灌木生物量表现出不同的分布规律。
2.3 地被层生物量和储量随林窗的变化
从表4可知,草本生物量和细木质残体储量表现为林窗和林缘高于林下,地被植物、凋落物和粗木质残体则表现为林下>林窗>林缘。草本层作为林分的最底层,对其生物量的研究常被忽视,但草本层对生态系统稳定、碳的循环速率、水资源保持及再分配等方面都发挥着重要作用。林窗和林缘的草本生物量显著高于林下,且林缘大于林窗,说明高山森林草本生物量受林窗的影响显著,并表现出明显的“边缘效应”,林窗改变了照射到地上的有效光辐射(Denslow等, 1998),增加了林窗和林缘的草本生物量。此外,地被植物多表现为喜阴、耐湿的生活习性,地被植物生物量与草本生物量分配格局截然相反。
凋落物和粗木质残体的储量受林分林龄及林分密度的影响较大,一般情况下,不同发育阶段凋落物和粗木质残体的储量受上层乔木的树冠郁闭度及树型等因素的影响。林窗的干扰又再次影响了乔木的郁闭度,凋落物和粗木质残体的储量表现为在林下最大,林缘最小。
2.4 林下生态系统不同类型生物量的重要性
灌木、草本和地表植物是森林生态系统林下活体植被的重要组成,它们参与了森林生态系统的物质循环和能量转化,凋落物和木质残体构成了森林生态系统林下的残体部分,它们连接着植物和土壤,为实现养分的归还和再次利用提供了可能。如表5所示,林下生态系统活体植被与残体部分的比例在1.77%~2.29%之间,活体植被与木质残体的比例为2.05%~2.55%,粗木质残体占木质残体的比例达到了83.46%~93.10%。灌木与活体植被的比例大于99.90%,说明灌木是构成林下活体植被的主要成分。虽然活体部分在林下占据着重要的位置,但其生物量与残体部分相比还是较低,说明高山森林生态系统凋落物和木质残体的储量巨大,尤其是粗木质残体的储量。

表4 地被层生物量和储量及其分布Table 4 Biomass and reserves on forest floor and its distribution kg·hm-2
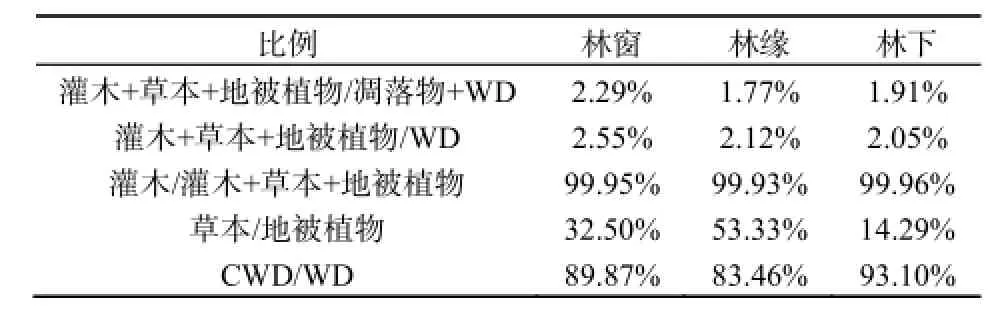
表5 林下不同类型生物量的比例格局Table 5 The proportion of different types of forest understory biomass
3 结论
(1)川西高山森林生态系统总生物量为72.75 t·hm-2,林下生物量为67.92 t·hm-2。其中,灌木生物量为9.81 t·hm-2,草本生物量为1.08 t·hm-2,地被植物生物量为3.86 t·hm-2,凋落物储量为1.50 t·hm-2,木质残体储量为53.00 t·hm-2。活体植被以灌木为主,残体部分以粗木质残体为主。
(2)林窗对灌木、草本、地被植物的影响各不相同,且不同物种的灌木生物量表现出不同的分布规律;草本生物量表现出明显的“边缘效应”,在林缘显著高于林下;林窗和林缘的地被植物生物量相对较低。
(3)粗木质残体储量从林下到林窗呈现减小的趋势,但总体储量仍然较大,林窗和林缘的细木质残体储量高于林下。
综上所述,频繁发生的极端灾害(雪灾、强降雨、干旱)、火灾、飓风等极端气候事件对生态系统造成了直接和间接影响,相对于其他森林,川西高山森林生态系统的总生物量更低,乔木生物量比例降低,灌木和木质残体比例升高,森林更新过程更为复杂。此外,林窗的形成导致微环境中的改变,从而引起微地形、光环境、空气温湿度和土壤养分及资源有效性等因子发生相应的变化,使森林环境形成较大的异质性,从而减少了整个生态系统的生物量。这些结果为认识高山森林生态系统林下生物量及其格局,以及林窗在森林生态系统的重要作用提供了基础理论依据。
DENSLOW J S, ELLISON A M, SANFORD R E. 1998. Treefall gap size effects on above-and below-ground processes in a tropical wet forest[J]. Journal of Ecology, 86(4): 597-609.
HARMON M E, FRANKLIN J F, SWANSON F J, et al. 1986. Ecology of coarse woody debris in temperate ecosystems[J]. Advances in Ecological Research, 15: 133-302.
LIU X, BAO W K. 2014. Understory plant assemblages present distinct short-term responses to the clear-cutting of an old-growth spruce forest near an alpine timberline[J]. Canadian Journal of Forest Research, 44(6): 562-571.
OAKLEY B B, NORTH M P, FRANKLIN J F. 2006. Facilitative and competitive effects of a N2fixing shrub on white fir sap lings[J]. Forest Ecology and Management, 233: 100-107.
TAYLOR A H, HUANG J Y, ZHOU S Q. 2004. Canopy tree development and undergrowth bamboo dynamics in old-growth Abies-Betula forests in southwestern China: A 12-year study[J]. Forest Ecology and Management, 200: 347-360.
YANG W Q, WANG K Y, KELLOMÄKI S, et al. 2005. Litter dynamics of three subalpine forests in Western Sichuan[J]. Pedosphere, 15(5): 653-659.
ZHANG Y D, LIU S R, ZHAO C M. Spatial pattern of subalpine forestrestoration in west Sichuan[J]. 2005. Chinese Journal of Plant Ecology, 16(9): 1706-1710.
程小琴, 韩海荣, 康峰峰. 2012. 山西油松人工林生态系统生物量、碳积累及其分布[J]. 31(10): 2455-2460.
崔宁洁, 刘洋, 张健, 等. 2014. 林窗对马尾松人工林植物多样性的影响[J]. 应用与环境生物学报, 20(1): 8-14.
方江平, 项文化. 2008. 西藏色季拉山原始冷杉林生物量及其分布规律[J]. 林业科学, 44(5): 17-23.
方精云, 刘国华, 徐嵩龄. 1996. 我国森林植被的生物量和净生产量[J].生态学报, 16(5):497-508.
郭剑芬, 杨玉盛, 陈光水, 等. 2006. 森林凋落物分解研究进展[J]. 林业科学, 42(4):93-100.
黄从德. 张健. 杨万勤, 等. 2009. 四川省森林植被碳储量的空间分异特征[J]. 生态学报, 29(9): 5115-5121.
黎燕琼, 郑绍伟, 龚固堂, 等. 2010. 不同年龄柏木混交林下主要灌木黄荆生物量及分配格局[J]. 生态学报, 30(11): 2809-2818.
林昌庚. 1964. 林木蓄积量测算技术中的干形控制问题[J]. 林业科学, 9(4): 87-97.
刘凤娇, 孙玉军. 2011. 林下植被生物量研究进展[J]. 世界林业研究, 24(2): 53-58.
罗辑, 杨忠, 杨清伟. 2000. 贡嘎山森林生物量和生产力的研究[J]. 植物生态学报, 24(2): 191-196.
吕勇, 刘辉, 王才喜. 2009. 杉木林分蓄积量不同测定方法的比较[J]. 中南林业科技大学学报: 社会科学版, 21(4): 50-53.
彭少麟, 刘强. 2002. 森林凋落物动态及其对全球变暖的响应[J]. 生态学报, 22(9): 1534-1544.
太立坤, 余雪标, 杨曾奖, 等. 2009. 三种类型森林林下植物多样性及生物量比较[J]. 生态环境学报, 18(1): 229-234.
陶建平, 宋利霞. 2006. 亚高山暗针叶林不同林冠环境下华西箭竹的克隆可塑性[J]. 生态学报, 26 (12): 4019-4026.
田大伦, 李雄华, 罗赵慧, 闫文德. 2014. 湘潭锰矿废弃地不同林龄栾树人工林碳储量变化趋势[J]. 34(8): 2137-2145.
王微, 陶建平, 李宗峰, 等. 2004. 卧龙自然保护区亚高山暗针叶林林隙特征研究[J]. 应用生态学报, 15(11): 1989-1993.
王祖华, 李瑞霞, 王晓杰, 等. 2010. 间伐对杉木人工林林下植被多样性及生物量的影响[J]. 生态环境学报, 19(12): 2778-2782.
吴庆贵, 吴福忠, 杨万勤, 等. 2013. 川西高山森林林隙特征及干扰状况[J]. 应用与环境生物学报, 19(6): 922-928.
杨昆, 管东生. 2006. 林下植被的生物量分布特征及其作用[J]. 生态学杂志, 25(10): 1252-1256.
杨昆, 管东生. 2007. 森林林下植被生物量收获的样方选择和模型[J].生态学报, 27(2): 0705-0714.
杨万勤, 邓仁菊, 张健. 2007. 森林凋落物分解及其对全球气候变化的响应[J]. 应用生态学报, 18(12): 2889-2895.
于立忠, 朱教君, 孔祥文, 等. 2006. 人为干扰(间伐)对红松人工林林下植物多样性的影响[J]. 生态学报, 26(11): 3757-3764.
张利敏, 王传宽. 2010, 东北东部山区11种温带树种粗木质残体分解与碳氮释放[J]. 植物生态学报, 34(4): 368-374.
张修玉, 管东生, 张海东. 2009, 三种森林粗死木质残体(CWD)的储量与分解特征[J]. 生态学报, 29(10): 5227-5236.
Understory biomass and its characteristics as affected by forest gap in the alpine forest ecosystem in west Sichuan
XIAO Sa, WU Fuzhong, YANG Wanqin, CHANG Chenhui, LI Jun, WANG Bin, CAO Yi
Long-term Research Station of Alpine Forest Ecosystem, Key Laboratory of Ecological Forestry Engineering, Institute of Ecology & Forestry, Sichuan Agricultural University, Chengdu 611130, China
Understory vegetation and its debris, as an important component in the forest ecosystem, can be greatly affected by forest canopy. As yet, little information has been available on the effects of forest gaps on understory plant and its debris biomass. In order to understand the changes of understory plant and its debris biomass as affected by forest gaps, therefore, three 100 m×100 m plots were sampled in a Minjiang fir (Abies faxoniana) primary forest ecosystem around 3 600 m altitude in western Sichuan in August, 2013. Coarse woody debris with length/height, basic diameter, diameter at breast height and decay class were investigated in each of three 20 m×20 m sampling quadrats in forest gap center, gap edge and closed canopy, respectively. Fine woody debris and shrub biomass were investigated in each of three 5 m×5 m sampling quadrats in forest gap, gap edge and closed canopy, respectively. Moreover, non-woody litter and herb biomass were harvested in each of three 1 m×1 m sampling quadrats in forest gap, forest edge and closed canopy, respectively. Each of 20 cm×20 cm sampling quadrat was randomly selected to harvest groundcover vegetation biomass in the above 1 m×1 m quadrat. The results indicated that: (1) the total biomass was 72.75 t·hm-2in the sampled alpine forest ecosystem, and understory biomass was 67.92 t·hm-2which accounted for 95.17% of the total biomass in whole ecosystem. Shrub biomass was 9.81 t·hm-2and occupied the majority proportion of living vegetation, but coarse woody debris biomass was 53.00 t·hm-2and occupied the majority of dead debris. (2) Shrub, herb and groundcover vegetation biomass showed different responses to forest gap. Shrub biomass was significantly higher in forest edge than that in closed canopy, although the biomass of groundcover vegetation was relatively lower in forest gap and forest edge compared with that in closed canopy. (3) Coarse woody debris biomass showed a decrease tendency from closed canopy to gap center, but the storage of fine woody debris in gap center and forest edge was larger than that in closed canopy. These results provide efficient foundation in understanding the understory biomass and its pattern in these alpine forest ecosystems as well as understanding the important role of gaps.
alpine forest; understory vegetation; biomass; forest gap
S715-3
A
1674-5906(2014)09-1515-05
肖洒,吴福忠,杨万勤,常晨辉,李俊,王滨,曹艺. 川西高山森林生态系统林下生物量及其随林窗的变化特征[J]. 生态环境学报, 2014, 23(9): 1515-1519.
XIAO Sa, WU Fuzhong, YANG Wanqin, CHANG Chenhui, LI Jun, WANG Bin, CAO Yi. Understory Biomass and Its Characteristics As Affected by Forest Gap in the Alpine Forest Ecosystem in West Sichuan [J]. Ecology and Environmental Sciences, 2014, 23(9): 1515-1519.
国家自然科学基金项目(31170423;31270498);国家“十二五”科技支撑计划(2011BAC09B05);四川省杰出青年学术与技术带头人培育项目(2012JQ0008;2012JQ0059);中国博士后科学基金特别资助(2012T50782)
肖洒(1988年),男,硕士研究生,主要从事森林生态学研究。E-mail:xxss513@126.com *通信作者:杨万勤(1969年),男,教授,博士,主要从事森林生态,面源污染控制与环境修复,生态工程方面的研究。E-mail:scyangwq@163.com
2014-06-30
